
Abstract
Lymphedema is a medically irresolvable condition. The lack of therapies addressing lymphatic vessel dysfunction suggests that improved understanding of lymphatic cell differentiation and vessel maturation processes is key to the development of novel, regenerative medicine, and tissue engineering approaches. In this review we provide an overview of lymphatic characterization markers and morphology in development. Further, we describe multiple differentiation processes of the lymphatic system during embryonic, postnatal, and pathogenic development. Using the example of pathogenic Kaposi's sarcoma-associated herpes infection, we illustrate the involvement of the Notch and PI3K pathways for lymphatic transdifferentiation. We also discuss the plasticity of certain cell types and biofactors that enable transdifferentiation toward the lymphatic lineage. Here we argue the importance of pathway-associated induction factors for lymphatic transdifferentiation, including growth factors such as vascular endothelial growth factor receptor-C and interleukins, and the involvement of extracellular matrix characteristics and dynamics for morphological functionality.
Introduction
T
Here we review the various principles governing cellular differentiation and morphogenesis toward lymphatic vessel formation in vivo and in vitro, focusing on factors determining cell fate and distinctive biomarkers, which allow a detailed observation of these processes.
Characterization of Lymphatic Endothelial Cell Commitment
Markers for lymphatic development and differentiation
The ability to identify lymphatic endothelial cells (LECs) with specific markers has facilitated understanding of cell differentiation into the lymphatic lineage. The most common markers used to characterize LECs from other cells are prospero-related homeobox transcription factor (Prox-1), endothelial hyaluronan receptor (LYVE), Podoplanin, and vascular endothelial growth factor receptor-3 (VEGFR-3). A detailed review about markers in lymphangiogensis has recently been published by Baluk and McDonald [1]. Here we focus on markers with the specific purpose of attempting to document the differentiation process from an undifferentiated cell type into the lymphatic lineage and their subsequent maturation into tubular structures.
Prox-1 is expressed at an early stage in embryonic development, and represents the first cell commitment toward the lymphatic lineage [2]; in adulthood it controls the on/off set of lymphatic genes related to morphogenesis [3], in conjunction with homeobox protein-D8 transcription [4]. Prox-1 has been successfully used to identify differentiation of LECs in vitro; moreover, blood endothelial cells (BEC) have been reprogrammed to express a lymphatic phenotype using Prox-1 [5]. Here, cellular reprogramming was associated with suppression of several BEC characteristic genes such as E-selectin, interleukin (IL)-8, laminin, collagens, neuropilin-1, and vascular endothelial growth factor C (vegf-c), and with induction of several lymphatic characteristic genes, including VEGFR-3, podoplanin, and desmoplakin I and II. Prox-1 overexpression in BECs induced only 20% of lymphatic genes when compared with freshly isolated lymphatic cells and suppressed only 40% of blood vessel genes compared with nonreprogrammed BECs. These studies serve both to highlight the importance of Prox-1 transcription in the differentiation process, as well as the need for additional factors to change cell fate terminally [5,6]. Prox-1 and nuclear factor of activated T-cells cytoplasmic calcineurin-dependent 1 (NFATc1) are colocalized exclusively in LECs during embryonic development, promoting the organization of sprouting LECs [7].
Other biomarkers commonly used to demonstrate lymphatic competence (Table 1) are VEGFR-3/Flt4, LYVE-1, and chemokine expression of secondary lymphoid-organ chemokine (CCL21/SLC). VEGFR-3 and its associated growth factor initiate migration from the anterior cardinal vein [8]; it is required for migration and proliferation, but not for vessel tubulogenesis in adults [9]. LYVE-1 represents a major component of the extracellular matrix (ECM) binding to hyaluronan, although it is expressed during embryonic development in early lymphatic and blood vessels [10 –12], it is absent from mature BECs [13]. The complete functions of LYVE-1 in lymphatics remain largely unknown.
LECs, lymphatic endothelial cells; NFATc1, nuclear factor of activated T-cells cytoplasmic calcineurin-dependent 1; Prox-1, prospero-related homeobox transcription factor-1; VEGFR, vascular endothelial growth factor receptor; BECs, blood endothelial cells.
A specific molecule for immunological cell–cell communication within the lymphatic system is CCL21, which is induced by VEGF-C [14], and thus is a late marker in embryonic development. Its paracrine function directs CCR7-positive cells, such as mesenchymal stem cell (MSCs), toward lymphatic vessels. In inflammatory conditions, in particular, CCL21 recruits MSCs to support tissue regeneration, as these cells transdifferentiate into numerous other cells types [15,16].
Although it appears that there is sufficient evidence that expression of Prox-1, NFATc1, VEGFR-3, and LYVE-1 contribute to the early commitment of LECs, the evidence is not incontrovertible as illustrated in the development of hybrid blood and lymphatic vessels, which can express most of the lymphatic markers [17,18]. Separation of lymphatic from blood vessels in embryonic development requires lymphocyte cytosolic protein 2 (SLP-76) and spleen tyrosine kinase (Syk) signaling [17]. Fu et al. [18] hypothesized that the maintenance of the lymphatic phenotype in adult tissue requires persistent expression of O-glycoproteins, which are associated with the surface expression of podoplanin/T1α. Podoplanin and SLP-76 are supposed to cointeract and are required during lymphatic blood separation [19]. During embryonic development, podoplanin expression is maintained on Prox-1-positive cells by budding of cells committed to the lymphatic lineage [20]. Moreover, podoplanin is an essential part of the motile/adhesive apparatus in the plasma membrane of the lymphatic vessel wall [20 –22], which promotes adhesion and migration [20], and is essential for vessel formation [23].
LECs express several additional biomarkers during their development, which are mostly shared with blood endothelial cells. However, in combination with the biomarkers presented in Table 1, the view of the temporal differentiation process could be expanded. In general, a sufficient number of biomarkers must be examined to minimize the risk of misinterpretation and to conclude, without doubt, a true lymphatic commitment. In addition, most lymphatic markers such as Prox-1, LYVE-1, NFATc1, and VEGFR are expressed less likely on mature LECs [24], which makes a broad analytical approach including the morphological structure essential.
Morphology of LECs and lymphatic vessels
In 2D in vitro culture, LECs adopt a cobblestone morphology similar to vascular endothelial cells [25 –27]. The junctions between adjacent LECs form a characteristic wavy outline giving them the appearance of an oak-leaf in profile. Although this renders the monolayer relatively impermeable, it is, nevertheless, selective to fluid and cell trafficking by formation of open junctions and flap valves [25,28,29]. LECs form typical junctional complexes with adherens, tight and occasionally gap junctions [30]. Unlike mature lymphatics, immature lymphatic cells are connected by zipper-like junctions, which develop to form discontinuous, button-like tight junctions and zona occludens consisting of linear segments of platelet endothelial cell adhesion molecule-1, VE-cadherin, and occludin, in addition to claudin-5, T-cadherin, and plakophilin 2 [25,28]. Therefore, phenotype of initial lymphatics can be determined by expression of cell–cell adhesion markers, and morphology oak-leaf characteristics of mature cells. Beside cell–cell adhesions, cell–matrix adhesions interaction via integrins (eg, α9β1) [5] and expression of ECM proteins like elastin microfibril protein 1 (Emilin1) [13,31], fibronectin [32], collagen type IV [32] and type XVIII [33], reelin, and the establishment of anchoring filaments through an irregular web of fibrillin deposition also show the temporal development of immature, initial lymphatic differentiation toward lymph collective vessels [34]. Initially, LECs lack a basal lamina and connect directly with anchoring filaments that radiate from the endothelial cells into a network of oxytalan, elaunin, and elastic fibers of the connective tissue [35], but this becomes more and more complex during lymphatic maturation. Mature vessels develop a interwoven elastin network around the vessel that connect to distinctive formation and orientations of collagen fibers to support its mechanical stability [36]. The lymphatic system matures from early stage tubular structures into larger collecting vessels with internal valves and ducts, the walls of which contain 1 or 2 layers of smooth muscle cells (SMCs). The lumen of the latter collecting vessel is irregular, between 20 and 70 μm in diameter [35]. The characterization of the lymphatic morphology and adhesion molecule expression illustrates the unique features of lymphatic capillaries during development and functionality.
To date, specific markers are limited to characterize lymphatics beyond the developmental stage and hence a broad analysis of the vessel morphology and ECM and is required to confirm lymphatic phenotype (Fig. 1). Nevertheless, documentation of the lymphatics is well defined, and state-of-the-art analytical tools are available to undertake research in the transdifferentiation process and biological development of the lymphatic system.
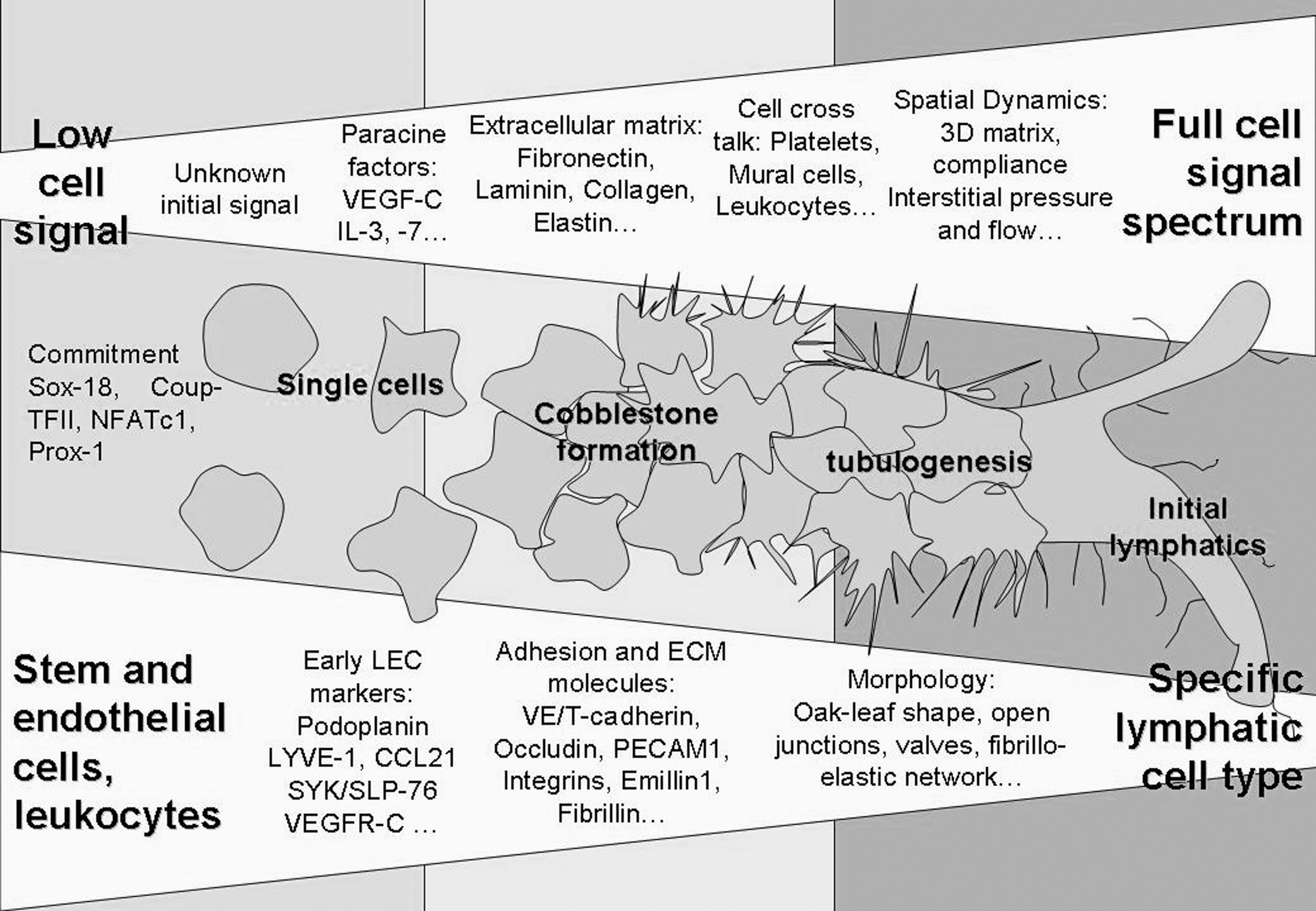
In vitro cell trans-differentiation with respect to cellular signals, such as intercellular communication and environmental factors, shows that the closer the cell environment is to the native state, the more specifically the lymphatic cell type responds. The inducing trans-differentiation signal is yet unknown, but paracrine signals like vascular endothelial growth factor (VEGF)-C lead to initial lymphatic endothelial cell phenotype. Additional factors provided by the extracelluar matrix (ECM; eg, fibronectin and collagen) or through cell crosstalk (eg, mural cells) and hemodynamic factors (eg, interstitial flow) are required to terminate the lymphatic cell fate and specify morphology and function toward initial lymphatics and collecting vessels. The developing stages can be observed through expression of lymphatic-specific markers in early development. ECM and adhesion makers as well specific morphology can be assigned to monitor maturation of the lymphatics.
Source and Formation of LECs In Vivo
Embryonic development of the lymphatic system
The favored theory in embryonic development of lymphatic tissue is that LECs arise by sprouting from embryonic veins in the jugular and perimesonephric areas [63]. This differentiation process is initiated by the promoter fragment SOX18 (sex determining region Y box 18) at the anterior cardinal vein [64]. It promotes the commitment of embryonic venous endothelial cells into the lymphatic lineage by activation of Prox-1 [65]. Together with the NFATc1 [7] and the orphan nuclear receptor (Coup-TFII) [66], it leads to protein expression and morphological characteristics of LECs [6,67]. Although it has been argued that this process is solely venous in origin, hematopoietic- or mesenchyme-derived Prox-1-positive lymphatic progenitors are present during embryonic development, but it seems that these cells do not contribute to the lymphatic development in embryonic mice [68]. However, it has been proposed that LECs originate from mesodermal lymphangioblasts as has been observed in chicks [69]. During embryonic development the mesenchyme of the dermatomes and the mediastinum gives rise to Prox-1-positive circulating mesenchymal cells and leukocytes, which integrate into growing murine lymphatics [70]. It may be argued that lymphatics may have a dual origin, and are able to rise from several cell types (Fig. 2). However, it is questionable whether nonvenous-derived endothelial cells independently form lymphatic vessel and organs, or whether they support the lymphatic development as observed in postnatal vessel formation. More fundamental research is required to confirm that nonvenous-derived LECs are same with venous-derived LECs to establish beyond doubt a dual origin for these cells.
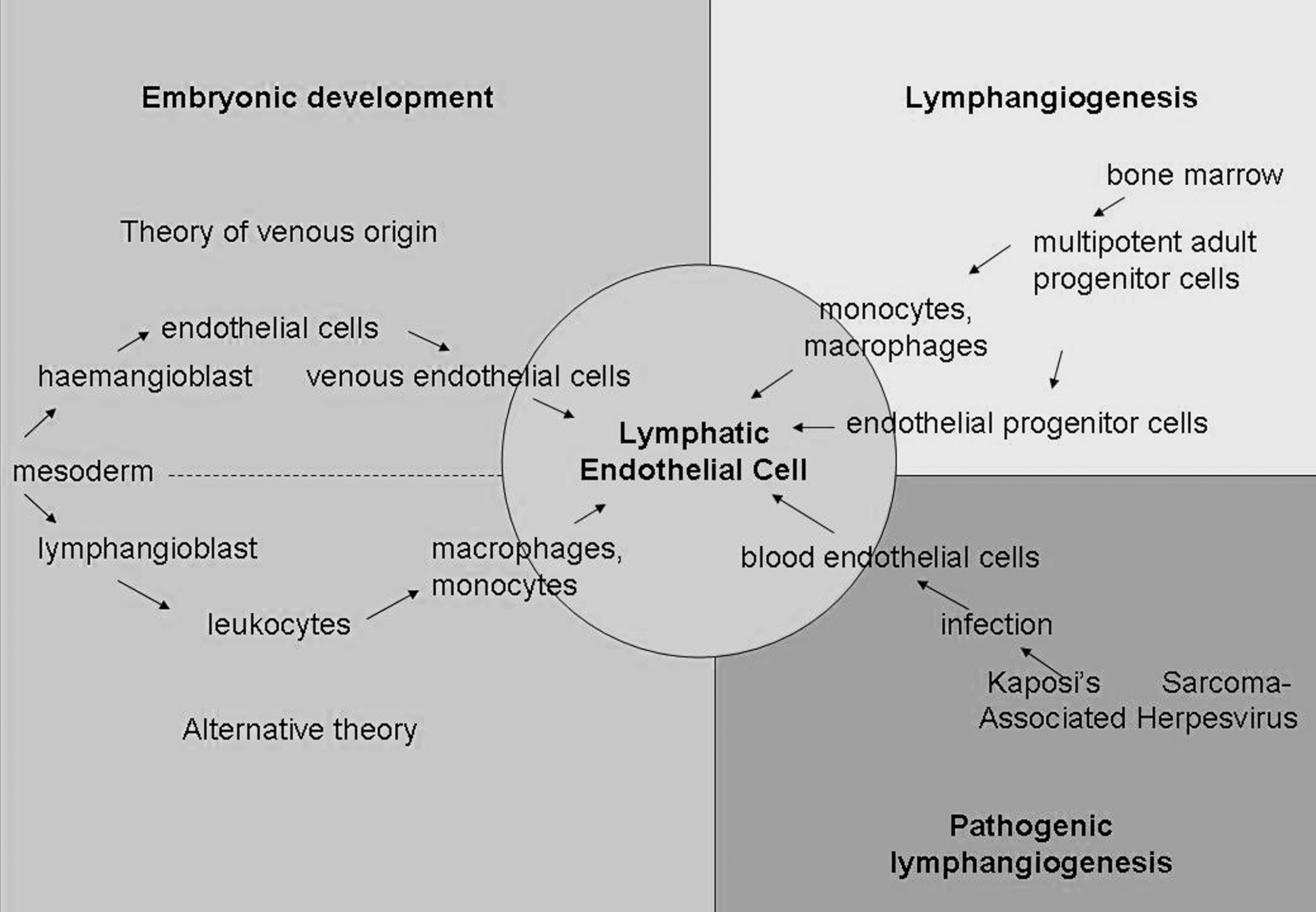
Known differentiation principles of cells in vivo toward the lymphatic phenotype. The figure shows embryonic development of the lymphatic endothelial cells, lymphangiogenesis, which occurs during wound healing in adult tissue and pathogenic lymphangiogenesis. Cells participating in the differentiation process are restricted to the embryonic or bone-marrow-derived monocytes, macrophages, and endothelial progenitor cells. In addition, viral reprogramming through Kaposi's sarcoma-associated herpesvirus of the closely related blood endothelial cells shows a unique principle in lymphatic trans differentiation.
Postnatal development of LECs
Direct observation of the differentiation of multipotent adult stem and progenitor cells have been made for the most part during wound healing. Endothelial progenitor cells (EPCs) and monocytes derived from multipotent adult stem cells of the bone marrow are stimulated to differentiate into vascular endothelial cells as part of the process of postnatal vasculogenesis [71]. An analogous principle for lymphatics has been assumed, and the involvement of circulating endothelial progenitor cells (CEPs) in adult lymphangiogenesis has been demonstrated in kidney transplants [72], and within angiomyolipomas [73]. Stronger evidence for this assertion is provided by observation of bone-marrow-derived CD11b+ macrophages and VEGFR-3-positive EPCs, which gave rise to neolymphatics in the cornea [74,75]. These studies suggest that bone-marrow-derived monocytes and CEPs may play a role in secondary lymphangiogenesis in analogous fashion to vasculogenesis.
Hematopoietic cell-derived CEPs are similar to bone-marrow-derived CEPs and are actively incorporated during adult hemangiogenesis [76], but do not appear to be incorporated during lymphatic development [74]. Despite their close relationship to LECs in terms of lineage, CEPs lack the ability to express Prox-1[77], which initiates the differentiation cascade and the separation between blood vessels and lymphatics [63]. Further, the regulation of the vascular and lymphatic phenotype is controlled via Syk and SLP-76 expression and their absence in mature vascular tissue, led Sebzda et al. [47] to conclude that “endothelial precursors destined for the lymphatic system must first completely exit the blood vascular system and failure to do so could result in blood-lymphatic connections.”
Therefore, it is unlikely that hematopoietic stem cells and hematopoietic cell-derived CEPs are able to differentiate toward the lymphatic lineage in vivo, due to their close but distinctive systemic separation. It is known from in vitro studies that hematopoietic cell-derived CEPs are able to express lymphatic endothelial-specific surface markers [78 –80], which suggests their transdifferential potential; however, it remains to be shown whether or not these cells remain systemically functional or develop a hybrid phenotype.
Pathogenic development of LECs
A third pathogenic form of lymphangiogenesis is observed in AIDS patients. Kaposi's sarcoma (KS) is a highly vascularized tumor of mainly spindle cells of endothelial origin [81]. The latent pathogen in KS is KS-associated herpesvirus. KS-associated herpesvirus infection of blood endothelial cells induces LEC phenotype, including expression of transcription factors Prox-1, VEGFR-3, and podoplanin, generally with an accompanying upregulation of >70% of the previously identified lymphatic lineage-specific genes [82,83]. The detailed mechanisms are unclear, but it is believed that KS-associated herpesvirus induces first de-differentiation toward EPCs indicated by CD133 expression [84], followed by differentiation toward the lymphatic lineage.
Certain other pathogenic conditions are known to induce lymphatic growth such as filarial infections and cancer. Extremely high numbers of Prox-1-expressing cells have been detected in malignant tissues [85], and in the case of filariasis neotubulogenesis of the lymphatic vessels is induced [86]. Both medical conditions indicate an underlying mechanism for cellular transdifferentiation toward the lymphatic lineage. Pathogenic models are useful tools for the understanding of transdifferentiation mechanisms. Currently, the KS-associated herpesvirus offers the best opportunity to explore conversion of blood endothelial cells into lymphatics in detail, which may provide insights into tissue regeneration as well as treatments for pathogenic lymphangiogenesis as a result of tumors or filarial infections.
Cell types having the potential for neolymphatic development
Cells involved in embryonic, adult, and pathogenic lymphangiogenesis (Fig. 2) all show a high potential for differentiation in vitro. Moreover putative lymphatic EPCs have been identified in human fetal liver and cord blood [77,80]. In this regard VEGFR-3, CD34, and CD133-positive stem and progenitor cells from different tissue origins were shown to be capable of differentiation toward the lymphatic lineage [80]. CD34-expressing cells from adipose tissue can also differentiate into EPCs [87], and LYVE-1 has been reported to coexpress with adipocytes [49]. Besides endothelial (progenitor) cells, embryonic and bone-marrow-derived stem cells have been shown to possess the capability to transdifferentiate into the lymphatic lineage (Table 2). Other cell types, such as adipose tissue stem cells, stromal cells, and leucocytes may also be capable of transdifferentiation into the lymphatic lineage; the latter cells that reside in and are transported via the lymphatic system interact directly with LECs, express certain lymphatic markers (eg, Syk), and directly influence lymphangiogenesis [88]. Leucocytes, especially macrophages, have demonstrated their plasticity during lymphatic regeneration [74], showing that lymphangiogenesic potential is not restricted to stem and progenitor cells only. Given that transdifferentiation toward the lymphatic lineage is facilitated by terminally differentiated cells of the myeloid lineage, for example, macrophages, it is reasonable to anticipate that certain proximal cell populations will possess the cellular plasticity needed to transdifferentiate into LECs and form into vessels.
Stimuli are shown in bold.
TCP, tissue culture plastic.
Signaling Pathways in Lymphatic Differentiation
The signaling pathways that are active during differentiation toward the lymphatic lineage are unknown, but some inferences can be drawn from the differentiation of vascular endothelial cells. As mentioned earlier, venous endothelial cells are believed to be progenitors of LECs during embryonic development; hence, venous signaling downstream is likely to be common to lymphatic differentiation too. In this regard signaling through PI3K/Akt activates the differentiation toward the venous type [89] by maintaining its identity through transcription factor Coup-TFII [90]. Although lymphatic cells are believed to be venous in origin, they display intercellular pathways, including ALK1, which have a crucial function in arterial [91] and lymphatic differentiation [92]. Further, LECs express Notch 1, 4 [93], which is essential for arterial differentiation [94]. Thus, ALK1 and Notch pathways suggest some crossover between lymphatics and arterial lineages.
Activation of Notch signaling pathway—including Notch1, 4 and its ligand, Delta-like 4 (Dll4)—is initiated through transcription factor FOXc2 and VEGF [95,96] (Fig. 3). Siekmann et al. argues that during early angiogenesis VEGF-A activates the Notch and MAPK differentiating while blocking PI3K and downregulating VEGFR-2, -3 [97]. Downregulation of VEGFR-3 has been achieved with a Dll4 zebrafish knock out model [98]. However, VEGFR-3 is upregulated during lymphatic differentiation by Notch signaling [93]. Moreover, infection with KS-associated herpesvirus in endothelial cells activates a signal cascade via membrane glycoprotein gp130 toward PI3K/Akt [83] and Notch signaling pathways are directly as well indirectly activated via Dll4 [99,100]. It is believed that KS-associated herpesvirus induces transdifferentiation to a lymphatic phenotype via those pathways. Hence, it can be argued that activation of PI3K/Akt and Notch signaling are significant for lymphatic transdifferentiation (Fig. 3). In direct contrast to arterial and venous differentiation, lymphatic cells have to maintain both PI3K/Akt and Notch signals. However, in the case of Notch-activation LECs might fail to activate their essential PI3K/AKT pathway, because the activation of ERK/Notch inhibits PI3K/Akt pathways and conversely PI3K/Akt inhibits ERK/Notch in endothelial cells to maintain either the venous or the arterial phenotype [89] (Fig. 3). To regulate both pathways Notch has to be alternatively activated. Hayashi and Kume (2008) demonstrated in microvascular endothelial cells that activation of PI3K by VEGF-A increases FOXc2 inducer promoter activity of Notch-ligand Dll4 [101]. The activation of Notch via PI3K describes a new link between the 2 pathways, which could also play a role during lymphatic differentiation.
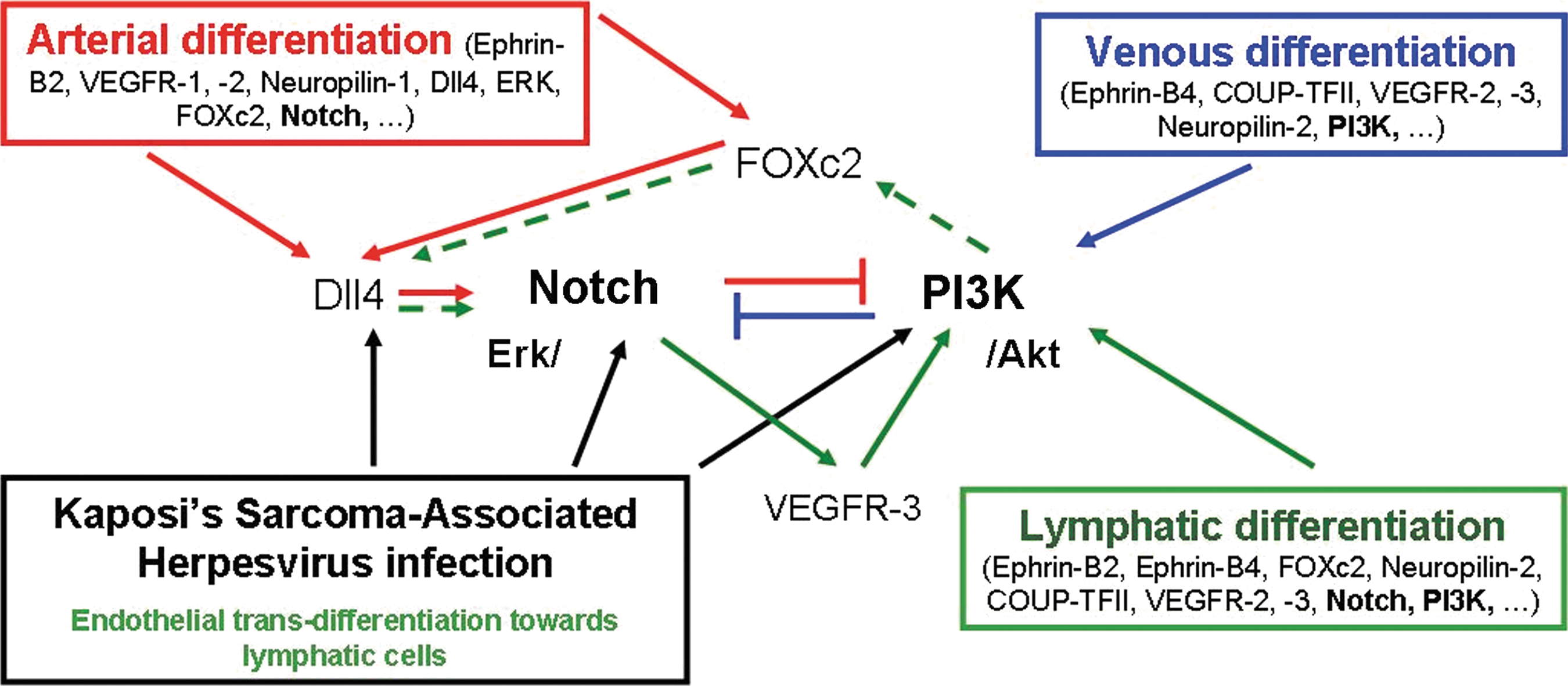
Arterial, venous, and lymphatic differentiation depend on Notch and PI3K pathway signaling. Arterial differentiation (red arrows) is multiple regulated through ERK or transcription factor C2 (FoxC2) toward Notch ligand Delta-like 4 (Dll4). Notch upregulates VEGFR-3 expression and degrades PI3K signaling to maintain its function. Venous differentiation (blue arrows) is maintained through PI3K/Akt signaling, which is the antipode to ERK/Notch. Lymphatic differentiation (green arrows) demands activation of both PI3K/Akt and Notch signaling. Kaposi's Sarcoma-associated herpesvirus activates both signaling pathways to differentiate endothelial cells into lymphatics (black arrows). Lymphatic signal pathways appear to activate the PI3K with soluble factors such as interleukin-3, interleukin-7, and VEGF-A/-C/-D via VEGFR-3, which again induces Notch ligand Dll4 activation via transcription factor FoxC2. Evidence of signaling up/downstream of Notch or PI3K/Akt toward essential lymphatic transcription factors such as Prox-1 has not been observed so far, but expression of both arterial and venous-specific, pathway-associated proteins by lymphatic cells suggest a strong link between arterial and venous development. Color images available online at
Nevertheless, much of the pathway of lymphatic differentiation remains obscure. It is not clear yet whether and how Notch and PI3K/Akt influence lymphatic differentiation and Prox-1 transcription. Further, the controversial role of COUP-TFII as an inhibitor of Notch in developing LECs (discussed by Oliver and Srinivasan [102]), and recent reports that LECs early commitment is independent of Notch signaling [66] via cross control mechanism introduced by Kang et al [103] contradict the assumption that Notch is a relevant pathway in lymphatic differentiation. However, during transdifferentiation and in pathogenic condition, hemangioma-derived stem cells differentiate into endothelial cells with distinctive Notch expression [104] and Prox-1-positive cells are strongly expressed in malignant tissue undergoing lymphangiogenesis [85]. The differentiation of cancer stem cells into endothelial cells is consistent with lymphangiogenesis, which indicates a role for Notch in lymphatic differentiation. Nevertheless, it remains to be shown how and when Notch signaling is maintained and activated in differentiating LECs, and whether same principles apply during transdifferentiation toward the lymphatic lineage. In summary, expression of pathways relevant to both arterial and venous development and the role of associated proteins such as EPHRIN-B2, EPHRIN-B4, neuropilin-1, -2, FOXc2, and Notch on LECs demonstrate considerable commonality with both venous and arterial pathways (Fig. 3). In this way conjoint venous and arterial pathways would align with the anatomical position of the lymphatic system that is in-between the venous and arterial vasculature.
Early and Late Regulators During the Differentiation Process of LECs
The factors that regulate cell differentiation toward the lymphatic lineage can be observed during embryonic development, where migration and differentiation are influenced by specific growth factors. In lymphangiogenesis LECs are generated/stimulated during the inflammation process. Cytokines released by immune cells (eg, dendritic cells and macrophages) play an important role by inducing cell proliferation, homing progenitor cells and stimulating the differentiation process.
Soluble factors
Vascular endothelial growth factors
Isoforms VEGF-A, -C, and -D have been identified as the most important factors in lymphangiogenesis. Lymphatic cells lack the VEGFR-1 receptor, which is expressed in BEC; therefore, LECs cannot be stimulated by VEGF-B [105]. Lymphangiogenesis requires activation of VEGFR-3 by VEGF-C/-D as reviewed by Lohela et al. [106], as well as a second requirement through the activation of VEGFR-2 by VEGF-A, as recently proposed by Kulkarni et al. [7]. VEGF-C signals through VEGFR-3 and thus can also induce secondary lymphangiogenesis in adult tissues [107]. The role of VEGFs for LEC migration, survival, proliferation, and inflammation is better understood than their relevance to transdifferentiation. Experimental evidence favors VEGF-C over VEGF-A as a primary inducer of LECs differentiation (Table 2) via VEGFR-3, but it is still uncertain which form of VEGF is critical and whether it induces transdifferentiation or simply drive the process once it is initiated. Kono and colleagues [108] demonstrated that VEGF-C and ang-1 were required but neither was sufficient alone to induce LEC differentiation from VEGFR-2 + cells in vitro. Further, it was shown that treatment of stem cells with VEGF-C was less potent at inducing a lymphatic phenotype than was the supernatant derived from LECs in culture [109]. Initially Kreuger et al. [78] suggested that “the process via VEGF-C alternatively has to be complemented by other signals for induction of lymphatic differentiation.”
Although the possible involvement of other as yet unknown factors cannot be ruled out in the initiation of differentiation toward an LEC phenotype, the importance of VEGF-C and -A is certain. As discussed earlier, the Notch and PI3K signaling pathways are activated to regulate sprouting and determine lymphatic cell fate. Activation of VEGFR-3 upregulates PI3K signaling in LECs [110]. Therefore, transdifferentiation toward the lymphatic linage via VEGFR-3 signaling cascades seems to be the most promising approach for induction and this has been successfully demonstrated in recent studies (Table 2).
Interleukins
Several ILs have been shown to trigger the differentiation processes toward LECs. Of the 35 known ILs to date, 2 are known to induce transdifferentiation in EPCs. IL-3 was found to be required for the maintenance of Prox-1 and podoplanin expression in blood endothelial cells (BECs) [111], whereas IL-7 has been shown to increase expression of lymphatic markers Prox-1, LYVE-1, podoplanin, and to stimulate tubulogenesis and capillary sprouting [112]. IL-7 is expressed by LECs acting in an auto-/paracrine manner [5]. IL-3 and IL-7 signal through the PI3K/Akt pathway [112,113]. Further, IL-7 increases expression of VEGF-D, which is similar to the main lymphatic growth factor VEGF-C, which in turn binds to VEGFR-3 and is transduced via a Wortmannin-sensitive (PI3K/Akt selective) pathway to promote lymphangiogenesis [112,114,115].
These findings suggest that other members of the common Y-chain family (IL-2/15, IL-3, IL-4, IL-7, IL-9, IL-13, and IL-21) may also have a role in lymphatic transdifferentiation. Stimulation of lymphangiogenesis and tubulogenesis has been observed with IL-1β, IL-12 [112], and IL-20 [116], whereas most other cytokines show no direct effect or have not yet been considered (q.v. Table 2). The potential of these ILs to induce lymphangiogenesis is, as yet, unknown, but the lineage similarity between LECs and BECs suggests at least a potential role for a wide range of cytokines during lymphangiogenesis. For instance, IL-3 [117], IL-8 [118], and IL-15 [119] are well known to stimulate BECs. Both LECs and BECs respond to most angiogenic cytokines, and in a similar manner owing to their similar IL receptor expression, but with some differences: KS-associated herpesvirus infection, which reprograms BEC toward lymphatic lineage, upregulates IL-6 and downregulates IL-8 [120]. The downregulation of the angiogenic factor IL-8 suggests a reduced effect on lymphangiogenesis compared with angiogenesis; moreover, IL-12, though highly anti-angiogenic, did not affect LECs significantly [121], whereas it induced podoplanin expression in microvascular endothelial cells [112]. The IL-12 family (IL-12, IL-23, IL-27, and IL-35), especially IL-12 (anti-angiogenic) and IL-23 (angiogenic), acts through a common surface receptor [122]. This could indicate a fundamental difference in intracellular responses to the same receptor stimulation in these 2 endothelial cells types and is thus worthy of more detailed study. These findings identify the possibility to direct transdifferentiation and development toward LECs, and ways to explore a unique signaling pathway that sets LECs apart from BECs.
Cell–cell and cell–ECM interactions
Although the majority of studies on differentiation have focused on soluble factors that work in a paracrine manner, juxtacrine factors and involvement of the ECM deserve consideration also.
Cell–cell communication
LECs strongly express proteins involved in transport and vesicle formation for intercellular communication [25]. In coculture experiments, induced pluripotent cells when seeded with OP9 cells, a line of bone-marrow-derived mouse stromal cells [123], resulted in differentiation of the induced pluripotent cells toward the lymphatic lineage [124], which indicates para- and juxtacrine stimulation. Equally, during lymphatic development, platelets adhere to the vascular wall and initiate blood lymphatic separation in juxtacrine manner [125]. Platelets C type lectin-like receptor −2 interacts with transmembrane protein podoplanin on endothelial cells, which is thought to initiate blood lymphatic separation via Syk and SLP-76 signaling [19]. The juxtacrine pathways also play a role in tubulogenesis, similar to that observed with vascular endothelial cells. First, the paracrine effect of VEGF-A induces Dll4 expression in tip cells, which activates a Notch-mediated juxtacrine signal between adjacent cells [126]. Juxtacrine factors become more important during lymphatic maturation. Mural cells, SMCs, are involved in the formation and function of collecting lymphatic vessels; however, the mechanism that initiates the transition from lymphatic capillaries toward collecting vessel, as well homing and functionalization of contracting SMCs is still unclear in lymphatic morphogenesis. Other cell types also communicate with the lymphatic endothelium; immunological cells such as monocytes have been observed to interact with LECs during early development, as these cells adhere and are transported by the lymphatic capillaries to the lymph nodes. The lymphatic endothelium is an interface for cell transport, and cell cross talk with mural and other proximal cell types like adipocytes or dendritic cells, though the exact mechanisms are unknown.
Cell–ECM interactions
An important role for the ECM environment during differentiation is well established. For example, it is known that the transcriptional programs that define BECs and LECs in vivo are lost rapidly in vitro without the native ECM. Therefore, the importance of the ECM and their integrin mediators in directing cell differentiation cannot be underestimated.
Integrins
The communication between the LEC and ECM components (ie, fibronectin and collagen) is mediated mainly through different sets of matrix-recognizing receptor molecules, namely, the integrins. LECs are known to express α1β1, α2β1, α4β1, α5β1, α6β1, α9β1, and αvβ3 integrins [127–131]. Expression of these integrins depends on developmental stages and tissue characteristics. During embrionic development LECs lack αv, α1β1, α2β1, and α4β1, but do express α5β1, α6β1, and α9β1 [129]. Integrin α5β1 is active in inflammatory lymphangiogenesis [132], whereas integrin α1β1 and α2β1 that bind laminin and collagen are expressed during wound healing [128]. Integrin α4β1 plays an important role in tumor-induced lymphangiogenesis [133]. The major integrins, α9β1, α4β1, α5β1, and αvβ3, which are the lymphatic fibronectin-binding partners, have been shown to regulate lymphangiogenesis; hence, these integrins and associate ECM proteins may also support transdifferentiation toward lymphatic lineage.
Many integrins interact with growth factors and/or upregulate their expression, which provides further evidence for the link between ECM components and the differentiation process. In collagen matrices LECs responded to VEGF-A by upregulation of integrin α1 and α2, with increased migration and tube formation [131]. Collagen and fibronectin matrices also stimulate migration through integrin β1/VEGFR-3 co-operation in microvascular endothelial cells [134]. Integrin α4β1 promotes adhesion and migration of LECs on cellular fibronectin and is strongly stimulated by VEGF-C, VEGF-A, and FGF-2 [133]. Further, fibronectin supports proliferation and survival of LECs via VEGFR-3 through integrin α5β1 [135], whereas integrin α9β1 was shown to function as a receptor for VEGF-C and VEGF-D [136]. The signal cascade is initiated through Prox-1 inducing integrin α9β1, which has been shown to be essential for lymphatic differentiation [137], especially during lymphatic valve morphogenesis, with the deposition of fibronectin-EIIIA ligand [129]. It follows that matrices that incorporate fibronectin in combination with VEGF-C are likely to promote the differentiation process.
Structural proteins
LECs express ECM components such as laminin, fibronectin [129,138], and collagens, but generate relatively little matrix compared with other endothelial cell types [25]. The provision of ECM proteins in vitro improves cellular processes such as differentiation, proliferation, and tubulogenesis, but the ideal substratum for LECs in vitro has yet to be determined.
Studies of lymphangiogenesis have revealed that LECs prefer a matrix that contains only fibrin [139], but others have shown that collagen type I hydrogels and matrigel® accelerated lymphatic capillary tube formation (Table 2).
Collagen is the preferred substrate for many cell cultures; however, the type of collagen that facilitates the differentiation process with regard to LECs remains largely uncertain. Many endothelial cells respond to collagen type I with increased proliferation, migration, and tubulogensis via activation of integrin receptor α2β1, while remaining quiescent on collagen IV that interacts with integrin α1β1 [140]. Further, matrix metalloproteases-2 and −9 downregulate collagen IV in the EMC to support growth of lymphatics [141]. Hence, it can be argued that basal membrane substrates are less likely to promote the lymphatic differentiation process. Nevertheless, collagen type IV supports LEC attachment and growth in vitro (Table 2), but like galactin-8, which promotes adhesion and migration but inhibits tubulogenesis [21], collagen IV substrata might inhibit lymphatic tubulogenesis.
Another major component of the ECM is hyaluronan. LECs express LYVE-1 the receptor for hyaluronan trafficking [142], but immobilized hyaluronan on glass surface inhibited growth and migration of LECs [143]. However, hyaluronic acid hydrogels show promise as a tool to control cellular differentiation [144], but have not been applied so far. Equally novel biomaterials or synthetic polymer scaffolds may yet offer new insights in lymphatic research. The available, though limited, evidence suggests that a fibronectin/collagen matrix may provide the necessary cues for cellular transdifferentiation toward the lymphatic lineage, but further work on cell–matrix interactions and ECM dynamics is required.
Mechanotransduction
In vasculogenesis, it is known that mechanical properties of the ECM play an important role in vessel formation [145], and it is not unreasonable to expect that this holds true for lymphatic vessels also. LECs establish anchoring filaments through an irregular web of fibrillin deposition, eluain fibers, interwoven elastic, and outer collagen fiber bundles to maintain their function, mechanical stability, and stage of differentiation [35,36]. These cell–matrix interactions can be manipulated either through ECM-associated components, and microstructural topography of the underlying surface [146] or through matrix compliance [139] and mechanotransduction arising from interstitial stresses generated by flow and hydrostatic pressure [147].
The surface topography directs cell attachment, migration, and orientation. As shown in our laboratory, LECs recognize fiber dimensions and interact better on microstructures similar to the dimensions of native collagen fiber bundles compared with nanostructures (unpublished data).
In general, LECs organize themselves into tubular structures more effectively within matrices that are more compliant [139], whereas on coated surfaces with collagen type I and VI and fibronectin (unpublished data, Table 2), they do not initiate tubulogenesis. The reason for the poor organization observed on coated plates is the lack of compliance and the restriction imposed by a 2-dimensional environment (unpublished data).
Interstitial stresses cause reorientation of the collagen fibers, which are supposed to interact with the fibrillae elastin network of LECs [36]. Fluid and/or mechanical stimuli in the form of stresses—whether cell-mediated or externally applied—act through deformation of the ECM, to which cells are attached. In LECs these changes are transduced by the fibrillo-elastic apparatus via integrin-mediated focal adhesions to the actin cytoskeleton, which, in turn, activates ERK1/2, p38, and MAPK pathways to regulate the physiological activity (eg, lymph transport) [148]. Mechanotransduction regulates not only cell physiology, but also differentiation of stem cells [149]. For example, when embryonic stem cells were treated with VEGF and subjected to physiological fluid shear stress, they differentiated into cells with endothelial phenotype [150]. Lymphatic growth is stimulated by an increase in fluid flow after blood vessel sprouting has occurred [151,152]. Fluid transport and matrix deformation is regulated by gradients in interstitial pressure, which affect the surrounding ECM structure to a greater or lesser degree according to its porosity and viscoelastic properties; these properties, in turn, will influence cell orientation, protein expression, morphogenesis, and differentiation [153]. Although interstitial fluid flow/transport influences cells during morphogenesis, and is required for complex tissue functions, the vast majority of in vitro assays are static. To our knowledge, only one group so far has created a 3-dimensional in vitro flow chamber [154], which they used to demonstrate the influence of fluid dynamics toward the lymphatic morphology [153]. Clearly, further work of this kind is essential.
Summary with Implications for Regenerative Medicine
The differentiation process from uncommitted single cells toward the lymphatic lineage and its maturation depends on multiple factors from soluble biomolecules to native tissue dynamics. In an orchestrated way, cell signaling increases in proportion to cell commitment and differentiation, and this can be monitored by expression markers, matrix, and morphological changes (Fig. 1). Cells capable of commitment to the lymphatic lineage in vivo predominantly originate from venous tissue during embryonic development and from bone-marrow-derived EPCs in adults. The origin of lymphatic cells has been challenged by an alternative theory involving mononuclear cells, which have also demonstrated their plasticity to develop into a lymphatic lineage in vitro, but stronger evidence is needed to argue for a dual origin of the lymphatics. Several cell types, such as embryonic and adult stem cells, macrophages, and endothelial cells, have also been demonstrated to differentiate and trans-differentiate into lymphatics in vivo and in vitro. The variety of cell types capable of transdifferentiation into lymphatics is thus inconclusive and the mechanisms and relevance in vivo have still to be established. Some insights may be provided by infections of endothelial cells with KS-associated herpesvirus, which leads to development of the lymphatic phenotype. The virus upregulates Notch and PI3K/Akt signaling pathways, which has been proposed to be a mechanism behind lymphatic transdifferentiation. Evidence in support of this theory is provided by transdifferentiation induced by VEGF-C, via PI3K, in various cell types. VEGF-C, IL-3, and IL-7 have been convincingly demonstrated to drive cellular transdifferentiation toward the lymphatic phenotype. However, the essential paracrine stimulation that initiates transdifferentiation toward the lymphatic lineage is still unknown and the required signaling toward transcription factor Prox-1 has not been identified so far. Thus, the possibility of other as yet unknown factors remains high. In general, the trans-differentiation process has been shown to include multiple factors paracrine, autocrine, and as yet unknown juxtacrine factors and matrix environmental properties. In addition, the involvement of a 3D fibrous compliant matrix has been identified as a requirement for the morphological functionality of lymphatics and in vitro models should attempt to recapitulate this environment as closely as possible. Mechanotransduction has been shown to be an additional factor for lymphatic maturation and retention of its morphological character. Despite these requirements for a more complex in vitro model, approaches with biomaterials, ECM proteins, and environmental characteristics have been limited thus far and require greater attention to pave the way for successful tissue engineering applications, which can be translated toward medical treatments.
Despite the aforementioned limitations of studies to date, the first steps to translate fundamental research in lymphatic development and differentiation into treatment of pathological conditions are underway. Recently, 3 groups demonstrated amelioration of secondary lymphedema in mouse tails with the application of MSCs [109], VEGF-C [157], and collagen gels [158]. The elimination of secondary lymphedema, especially in postsurgical breast cancer patients, is only one such field of application. In addition, primary lymphedema of hereditary origin, such as Milroy's disease and Meige's disease, could be also addressed, for example, by transdifferentiation of autologous leucocytes toward VEGFR-3-expressing LECs, which could then be re-introduced into the developing tissue. Finally, lymphangiomas, which are characterized by lymphatic disconnection, could be stimulated to develop new connections, with appropriate growth factors.
Treatments of lymphatic diseases have not progressed to the same extent as cardiovascular disease management. This is in part due to the lack of basic scientific understanding of the development of the lymphatic system. However, taking advantage of the similarities between the cardiovascular and lymphatic systems, and combining this with a greater understanding of the factors controlling lymphatic development, could lead to the rapid application of basic scientific understanding to the development of treatments for a variety of lymphatic disorders, for which no therapeutic strategies currently exist.
Footnotes
Acknowledgment
The University of Strathclyde is a charitable body, registered in Scotland, No. SC015263.
Author Disclosure Statement
No competing financial interests exist.