
Abstract
Hematopoietic stem cells (HSCs) accomplish a complex task. On a daily base billions of the 8 different mature cells are delivered in the right proportions. HSCs are located in niches located at several locations in the body. Communication between these spatially separated niches is accomplished by stem cells that leave their niche and migrate to other niches guided by soluble factors. The niche itself comprises all major signaling pathways (Hedgehog, Notch, Wnt, and BMP) and an array of adhesion molecules. The interplay between these components keep HSC in a quiescent state but also speed up production in case of urgent need during infection or excessive blood loss. In this review, we focus on the molecular cues of the niche, functional adhesion molecules and describe recent data obtained with multiphoton microscopy. A vast array of molecules is described that display similar functions as HSC controllers. This points to redundancy in the system that enables HSC to respond to different cues essentially with the same functional response. Apparently, the hematopoietic system is so crucial that it is not dependent on a single cue. When one cue fails to initiate a response, another cue will take over leading to an almost similar response. Another explanation is that every cue adds to an integrated signal that results in reaching the threshold. This integrated signal might be reached from huge signaling by a single cue or the low but additive signals by several cues.
Introduction
H
Characteristic to stem cells is their unique ability to self-renew. In theory, these cells have unlimited proliferative potential. In practice, however, mechanisms, like telomere shortening, prevent unrestricted proliferation. To preserve the pool of stem cells, it is essential to keep them quiescent under normal circumstances [1]. On the other hand, proliferation of stem cells is up-regulated upon extensive cell loss. The stem cell niche provides external cues that regulate the delicate balance between stem cell quiescence and proliferation.
The cells that are responsible for the production of all mature blood cells are the hematopoietic stem cells (HSCs). During embryonic development, the first definitive HSCs are formed at several anatomical sites. These HSCs migrate to the fetal liver where they proliferate and differentiate. In late embryonic stages, these HSCs migrate toward the bone marrow (BM; reviewed in Ref. [2]). During postnatal life, the BM remains the major compartment in which HSC reside. However, localization of HSCs is not restricted to the BM alone [3,4].
In the past 2 decades immunohistochemistry studies revealed that BM HSCs reside at the inner surface of the bone, the endosteum. Recently published articles shed new light on the in vivo localization and trafficking of immature HSCs. The different components of the endosteal niche are highlighted and several articles that emphasize the importance of the osteoblasts at the endosteal surface are discussed. An emphasis is put on the molecules and principles that mediate intercellular signaling between the osteoblasts and the HSCs. The main points are summarized and some problems that still have to be overcome to resolve the HSC niche in future research are put forward.
Localization of HSCs in vivo
In the past several years, much progress has been made in the field of cell imaging methods. Recently, 2 independently published articles describe in vivo trafficking of HSCs by imaging in time. Lo Celso and colleagues [5] used 3-dimensional in vivo imaging to observe hematopoietic stem/progenitor cell (HSPC) localization. HSPCs injected into non-irradiated hosts were dispersed throughout the BM, whereas large amounts of HSPCs injected into irradiated hosts were observed in close proximity to the endosteal surface [5]. The endosteum is a layer composed of mesenchymal tissue that covers the inner site of the compact and trabecular bone. It appeared that localization of the HSCs was influenced by their differentiation state. Immature long-term (LT) reconstituting HSCs were found close to the endosteum, whereas the more mature subsets were residing at larger distances from the endosteal surface. In addition, cells that did not show high proliferation rates were significantly closer to the osteoblasts. These results suggest that immature HSC are maintained quiescent at the endosteal surface and differentiate upon translocation into the BM center.
Similar results were obtained by Xie and colleagues [6]. By using ex vivo imaging stem cells (EVISC), they were able to follow HSCs after injection in either nonirradiated or irradiated mice. Again, HSCs injected into irradiated mice migrated to the endosteal region at the bone and BM interface whereas HSCs injected into nonirradiated showed a random distribution throughout the BM. In addition, they found that HSCs favored homing to the trabecular bone in irradiated mice, whereas they did not show any preference to either the trabecular or compact bone in nonirradiated mice. A luciferase assay visualizing HSCs revealed a strong and persistent signal in trabecular bone after BM damage. In contrast to the trabecular bone, only transient signals were observed in the spleen as well as in the liver, indicating short-lived HSC proliferation at these locations. These data indicate that after BM damage—emptying the niche—HSCs are attracted to the endosteal region where they proliferate to replace the lost cells. This was endorsed by the fact that 6 months after injection these HSPCs did contribute to host hematopoiesis and a significant part of the hematopoietic cells was comprised of their progeny [5].
More data were recently published that suggest that the primitive LT-HSCs are kept quiescent at the endosteal surface. Haylock and colleagues report that HSCs that are tightly connected to the endosteum have greater in vitro and in vivo potential compared with HSCs that reside in the central marrow [7]. By flushing the bones central BM HSCs were obtained. However, a substantial part of all the HSCs remained within the endosteal region, which was isolated subsequently. A clonogenic assay revealed that the proliferative potential of endosteal HSCs was significantly higher compared with the central BM HSCs. Moreover, upon transplantation in nonablated mice both HSCs subsets re-migrated to the location from which they were taken. Analysis of donor cells 12 weeks after transplantation of endosteal HSCs and central BM HSCs revealed a significant greater contribution of the endosteal HSCs to donor hematopoiesis compared with central BM HSCs [7], indicating that their functional potential is higher as well. Moreover, these data suggest that immature LT-HSCs express a specific set of chemotaxis receptors and/or adhesion molecules that interact with molecules present at the endosteal niche. These articles confirm previous observations that immature LT-HSC reside at the endosteum and that more mature progenitors are further away from the bone [8]. This indicates that endosteal-specific signaling molecules are essential in maintaining the immature phenotype of HSCs. Upon extrinsic cues that enhance HSC proliferation, for example, blood loss, however, offspring of the proliferating HSCs will be forced to migrate further away from the endosteal niche. It is thought that the transmigration of HSCs into the central BM deprives them from signals that maintain them immature and as a consequence differentiation will occur. However, which components of the endosteum are critical for HSC maintenance, and how they function, is not fully explored.
Above mentioned studies assign an essential role to the endosteal niche in maintenance of HSCs. However, some studies show contradictory results. For example, no endosteal niches are present in embryos and HSCs are present in extramedullary tissues during adult life. Therefore, it has been proposed that the endosteal niche is not exclusively responsible for maintenance of HSCs. In vivo imaging techniques have shown that endothelial cells are actively involved in the recruitment of HSCs to the BM in lethally irradiated mice [9]. In addition, Kiel and colleagues used a specific set of HSC markers that revealed that 60% of the BM HSCs localized adjacent to sinusoids, whereas only 20% localized to the endosteum [10]. Moreover, a recent study by Li and colleagues suggested that the role of these cells extends beyond the recruitment of HSCs [11]. Transplantation assays indicated that endothelial cells can promote self-renewal and regeneration of HSCs as well [12]. Nonetheless, how endothelial cells regulate the HSCs has hitherto remained elusive. Intracellular communication between the endothelial cells and HSCs mediated by cytokines might be responsible for this process as hematopoiesis is severely reduced in mice that lack the expression of the cytokine receptor gp130 in endothelial cells [13]. Although direct evidence is lacking, these results have led to a model in which endothelial cells provide specialized niches, so called “vascular niches,” that regulate HSCs maintenance either alone or in cooperation with the endosteal niche (reviewed by ref. [14]).
Numerous studies have indicated that the vascular niche are especially important when there is a increased demand for hematopoiesis. After radioablation or specific deletion of osteoblasts, the number of endosteal HSCs was reduced, whereas the number of early hematopoietic progenitors at extramedullary sites, that is, liver and spleen, was up-regulated. Consistent with an up-regulation of the number of hematopoietic progenitor cells, hematopoiesis in these peripheral organs was enhanced [15]. These results might be explained by a model in which quiescent HSCs reside at the endosteal niche, which translocate to the vascular niches upon stimulatory cues. Thereby, proliferation and differentiation are stimulated (reviewed by ref. [16]).
The endosteal niche
The endosteum is a layer of connective tissue consisting mainly of osteoprogenitor cells and their progeny, which covers inner site of the bone (Fig. 1). Different cell types at endosteum have shown to be involved in HSC regulation.
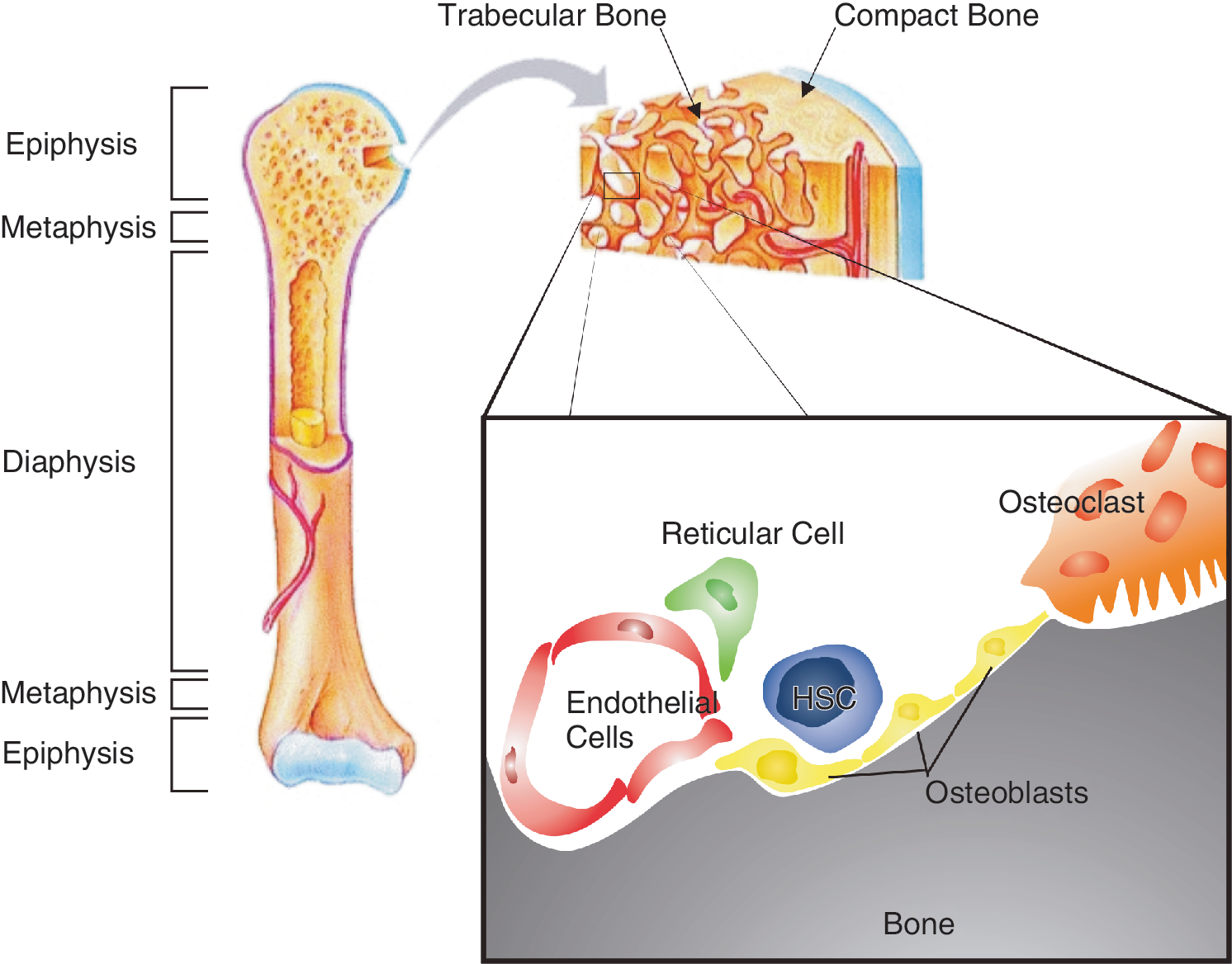
The hematopoietic stem cell (HSC) at the endosteal surface. The HSCs specifically home to the trabecular bone in the metaphysis and epiphysis. At the endosteal niche in the trabecular bone, different cell types surround the HSCs. These neighboring cells are all able to regulate HSC behavior. Moreover, the high Ca2 + concentrations at the bone surface attract HSCs to this specific site. Color images available online at
Osteoprogenitor cells originate from the mesenchymal stem cells (MSCs), which reside in the BM and are able to differentiate into osteoblasts besides other lineages. These osteoblasts are found lining the surface of bone. It has become clear that osteoblasts are essential for proper hematopoiesis. In the following part, the role of osteoblasts in creating the HSC niche and the molecules that are part of this process will be discussed.
Osteoclasts also line the endosteal surface. These cells are formed by fusion of multiple granulocyte–macrophage progenitor cells, a process mediated by osteoblasts. Osteoclasts are primarily responsible for degradation of bone. However, these cells are also implicated in mobilization of hematopoietic progenitor cells under conditions of hematopoietic stress [17]. Upon induction of stress, the number of activated osteoclasts at the endosteal region increased. In addition, these osteoclasts induced expansion of the number of hematopoietic progenitor cells in the blood stream. Enzymes secreted by osteoclasts were responsible for the release of HSCs from the endosteal niche. These enzymes were able to cleave factors that promote the interaction between HSCs and their niche [17]. On the other hand, results have been published that suggest that osteoclast activity can promote lodgment of HSCs at the endosteal niche. Osteoclast-mediated deconstruction of the bone leads to high levels of Ca2 + near activated osteoclasts at the endosteal surface. Adams and colleagues reported that HSCs carry the calcium-sensing receptor (CaR) that is a transmembrane G-protein-coupled receptor that is sensitive to the extracellular Ca2 + concentration. Moreover, they showed that in genetically engineered mice lacking the CaR the number of HSCs at the endosteal niche was significantly reduced and that physical association of CaR−/− HSCs with extracellular matrix (ECM) proteins at the endosteal niche was impaired [18]. These results imply that osteoclast activity mediates maintenance of HSCs at the endosteal surface by increasing the extracellular Ca2 + concentrations. Moreover, it has been hypothesized that proteases secreted by osteoclasts can release growth factors from the ECM, thereby influencing HSC maintenance. Taken together, these results suggest that osteoclasts have a dual role at the endosteal niche; both maintenance at as well as release of HSCs from the endosteal niche is mediated by proteins secreted by osteoclasts. As the expression pattern of proteases in osteoclasts depends on their activation status, the latter might be a critical factor in HSC regulation. The activity of osteoclasts is amongst others regulated by osteoblasts. Osteoblasts are able to promote proliferation, differentiation, and activation of osteoclasts through expression of the RANKL. Therefore, an osteoblast may indirectly regulate HSC maintenance and release through its interaction with osteoclasts. It has to mention, however, that activation of osteoclasts depends on multiple external stimuli.
Another important cell type at the endosteal surface is the reticular cell. These cells originate from the MSC and have osteogenic and hematopoietic potencies. In 2006 Sugiyama and colleagues showed that these cells can contribute to the maintenance of HSC at the endosteal niche as well [19].
The endosteal niche is a well-vascularized environment. Therefore, endothelial cells as well as signaling molecules provided by the blood stream may influence the endosteal niche [5].
Altogether, these data reveal that HSC maintenance at the endosteal niche is a complex process mediated by a diverse set of cellular and acellular interactors. Such a multiparameter regulation of HSC maintenance and growth ensures that a single factor cannot evoke total loss of the HSC pool because its action is also counteracted by other induced factors. Interaction of HSC with the niche points to a mechanism where binding to the niche and release is governed by integrating the on–off binding dynamics of several cell adhesion molecules (Fig. 2). This in contrast to adhesion molecules that act as a glue: removing the glue by soluble enzymes can lead to total release of HSCs.
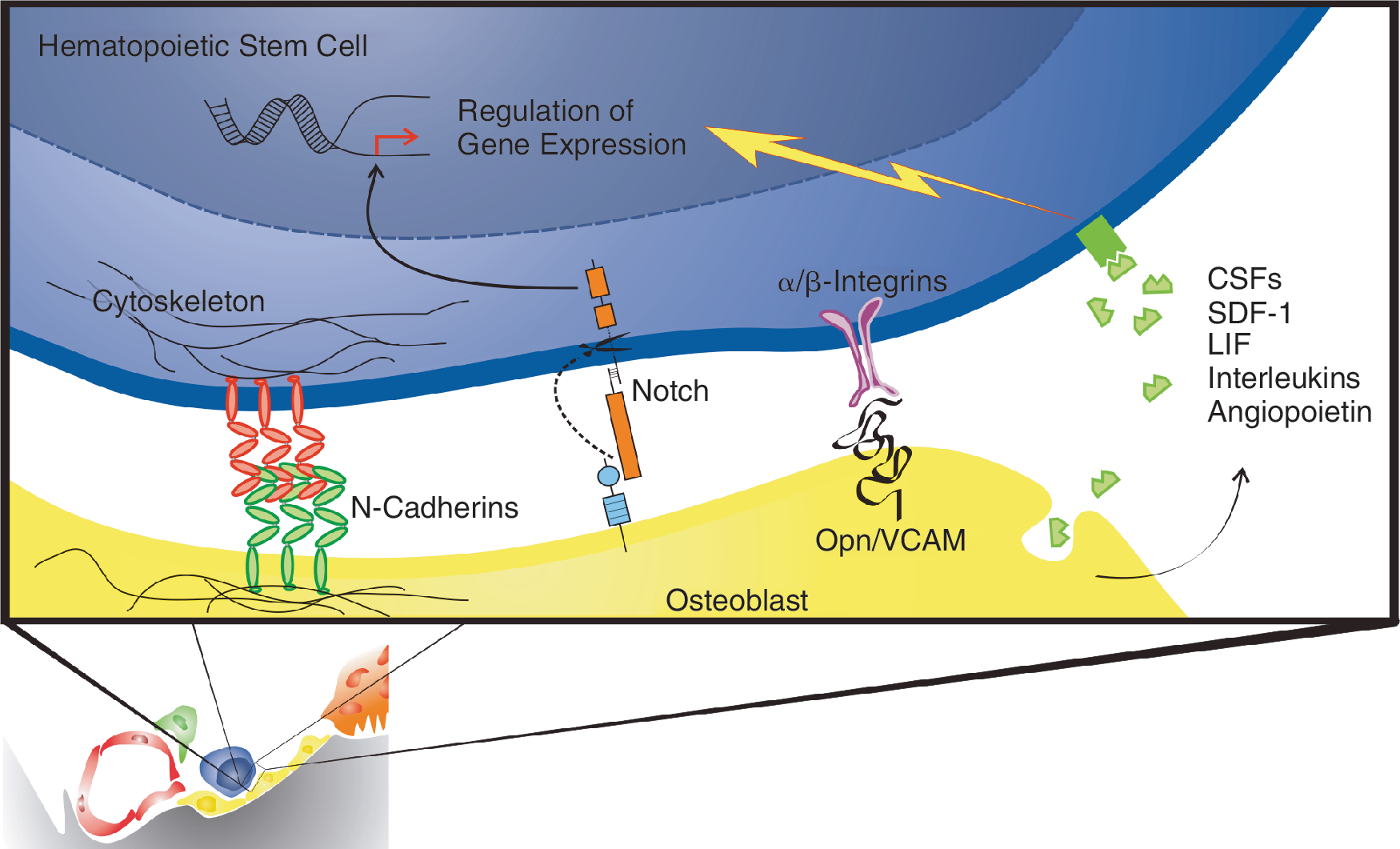
Molecular interaction between the osteoblast and the hematopoietic stem cell (HSC). Osteoblasts residing at the endosteal niche interact with HSC via soluble factors as well as adhesion molecules. Although it was proposed that physical interaction between both cell types is crucial for keeping the HSC quiescent, hard evidence is lacking. Interaction with the niche can also be explained by integrating on–off dynamics of cell adhesion molecules. Color images available online at
The role of osteoblasts in creating the HSC niche
Osteoblasts are responsible for the production of matrix, which they secrete at the site of the bone, and mineralization of the bone. Eventually, osteoblasts either get surrounded by matrix and end up as osteocytes or they become bone-lining cells, which is a reversible process. Since osteoblasts are key components of the endosteal surface, they are thought to play a pivotal regulatory role on HSCs. Therefore, the role of osteoblasts in HSC regulation has been extensively studied in vitro as well as in vivo. It was shown that osteoblasts support HSCs in vitro and that co-culture of osteoblasts improves HSC maintenance [20]. In vivo, the number of osteoblasts has also shown to correlate with the number of HSCs. In mice lacking the BMP receptor 1a, the formation of trabecular bone as well as the number of osteoblasts was increased. In addition, the number of donor-derived cells upon HSC transplantation was significantly higher in these mice as compared with littermate controls [21]. In line with these results, an up-regulation of the number of osteoblast due to the expression of a constitutively active parathyroid hormone (PTH) receptor was accompanied by an increase in HSC numbers [22]. Moreover, engraftment of HSC was improved when osteoblasts were co-transplanted into the donor [23]. On the other hand, disruption of osteoblastic development led to complete absence of the hematopoiesis [24,25]. Together, these results suggested that osteoblasts are crucial in regulating the number of HSCs. In order to study the importance of osteoblasts in hematopoiesis more directly, Visnjic and colleagues used a herpes virus thymidine kinase under the control of an inducible lineage-specific promoter. This construct allowed osteoblast-specific ablation of osteoblasts after treatment with ganciclovir (GCV). Upon administration of GCV, the BM cellularity decreased severely [15]. Together these data indicate that osteoblasts are essential for maintenance of the hematopoietic system in the BM.
The importance of osteoblasts in vivo in creating an environment in which HSC reside and contribute to the hematopoietic system has recently been confirmed by Dominici and colleagues [26]. Staining of osteoblasts after BM radioablation showed a significant up-regulation of the number of osteoblasts lining the endosteal surface in irradiated mice compared with nonirradiated mice. The uptake of bromodeoxyuridine (BrdU) indicated that the formation of multiple layers of osteoblasts was the result of enhanced proliferation of these osteoblasts. Donor HSC injected subsequently to engrafted radioablated mice in close proximity to the proliferating osteoblasts suggested that the contact with these osteoblasts is beneficial for HSCs engraftment. At later stages, however, donor hematopoietic cells were found throughout the BM and the number of osteoblasts was reduced to a normal level again. These data suggest that osteoblasts create an environment in which HSCs home.
To clarify the role of osteoblasts in HSC maintenance, proliferation, and differentiation, it will be important to map the intercellular signaling molecules that are known to have a regulatory function on HSCs expressed by osteoblasts.
Communication between osteoblasts and HSCs
HSCs proliferation and differentiation is regulated by a wide variety of cytokines, growth factors, and other signaling molecules. To determine the role of the osteoblast in creating the HSC niche, the proteins expressed by osteoblasts that have shown to influence HSC behavior are discussed.
First, several soluble signaling molecules that regulate HSC behavior are described. Some of these molecules enhance HSC proliferation whereas others keep HSCs quiescent. Among them, colony-stimulating factors (CSFs) and interleukins (IL) play a key role in regulating HSCs in the BM [27]. Although the expression on RNA as well as at the protein level of these molecules by osteoblasts was proven, it has to be taken into account that other cells express these molecules as well. Since these molecules are able to diffuse into the niche, it remains difficult to address the contribution of each cell type individually.
Besides soluble signaling molecules, there have been identified several transmembrane proteins with the potency to regulate HSC behavior. Since immature HSCs reside in close proximity to the endosteal surface, it is thought that these molecules mediate a direct connection between HSCs and osteoblasts essential for maintenance of immature HSCs.
Hematopoiesis is regulated by the adipocyte-osteoblast equilibrium
Recently, the role of adipogenesis as negative regulator in hematopoiesis was elucidated. Increasing the number of adipocytes in BM leads to less cycling hematopoietic progenitors and loss of short-term hematopoietic progenitors. The primitive HSC compartment remains largely intact. A 3-way co-regulation of hematopoiesis, osteogenesis, and adipogenesis was proposed [28]. Ovariectomy shifts the balance between the 3 interacting limbs. Changes in bone density, adipogenesis, and hematopoiesis support such 3-way co-regulation [29]. Since adipogenesis is prevented by endothelial cell contact and Wnt signaling [30], it is tempting to speculate that decay of the vascular niche precedes the gain in adipogenesis because loss of endothelial cells will shift the balance to more adipogenesis with subsequent impact on hematopoiesis.
Soluble signaling molecules
Colony-stimulating factors
Granulocyte (G)- and granulocyte–macrophage (GM)-CSF have an important role in the inducement of HSC mobilization. Therefore, both cytokines are widely used in the clinic to harvest large numbers of HSCs from the blood. The expression of G-CSF leads to release of HSCs at the site of the endosteal niche, followed by translocation to the blood stream. Release of the HSCs is an indirect result of the expansion and activation of myeloid cells and granulocyte precursor cells by these CSFs. Activation of myeloid and granulocyte precursor cells results in an overwhelming release of neutrophil proteases, like elastase, cathepsin, and matrix metalloproteinase (MMP)-9. Moreover, the expression of serine proteinase inhibitors is down-regulated upon expression of G-CSF. Together, this leads to elevated levels of active proteases in the BM. These proteases cleave and inactivate certain molecules, for example, CXCL12 and CRX4, which control HSCs maintenance in their niche [31] (reviewed by ref. [32]). In addition, G-CSF inhibits the expression of CXCL12 by osteoblasts [33]. As a result, adhesion of HSCs to osteoblasts is abrogated and the number of circulating HSCs is increased.
Taken together, these data indicate that CSFs at the endosteal surface mediate HSC loosening, resulting in a decrease in the number of endosteal HSCs. It has to be taken into account, however, that the level of active CSFs expressed by osteoblasts is highly variable, probably depending on external cues that regulate osteoblast behavior. This is a plausible explanation for results that show no expression of either G-CSF or GM-CSF [34]. After all, these factors will probably be expressed when there is a demand for circulating HSCs. There seems no logical reason for G-CSF or GM-CSF expression under in vitro culturing conditions where no HSCs are present. Therefore, it would be worthwhile to look at the expression levels under normal circumstances and circumstances in which HSC mobilization is required.
Interleukins
Interleukins are a large family of cytokines that play a pivotal role in intercellular communication between cells of the immune system. Most of the family members have a role in the induction of hematopoietic cell proliferation, differentiation, or activation. The HSC has also shown to be susceptible to the presence of ILs. Therefore, it is essential to consider the role of ILs in HSC maintenance and proliferation at the endosteal niche. This is confined to a certain subset ILs based on their expression pattern and the presence of their receptors on HSCs. When discussing the expression of ILs in a particular cell type, one has to take into account that the expression of cytokines is highly variable. This means that the expression levels strongly depend on external stimuli like culture conditions.
In vivo, it has been shown that the IL-1 receptor is present on HSCs and that it is involved in HSC proliferation [35]. Several reports indicate that IL-1 is provided by osteoblasts at the endosteal surface [27]. Data of Kim and colleagues, however, showed no expression of IL-1 in osteoblasts, which is contradictory to results from previous reports [27,34]. However, Kim and colleagues measured the IL-1 levels in steady-state osteoblasts cultured from MSC in vitro, whereas it has been shown that IL-1 expression can be enhanced upon stimulation [36].
Besides IL-1, another IL commonly referred to as regulator of the HSC is IL-6. IL-6 mRNA as well as protein expression was observed in osteoblasts [37]. IL-6 promotes ex vivo expansion of umbilical cord blood (UCB)-derived HSCs. The authors suggest that IL-6 maintains the immature phenotype of HSCs [12]. Remarkably, it turned out that IL-6 expression by osteoblasts is regulated by the HSCs among others. In the presence of HSCs near the osteoblasts, the latter express a significant higher amount of IL-6 [38], suggesting that HSCs can promote hematopoiesis via a positive feedback loop involving osteoblasts.
Circumstances in which enhanced production of hematopoietic cells is crucial as well are created by radioablation. Radioablation leads to cellular loss in the BM that needs to be overcome. Kang and colleagues have shown that upon radioablation the expression of IL-10 in osteoblast lining of the trabecular bone is up-regulated. Moreover, they show that the IL-10 receptor is expressed by HSCs and that exogenous expression of IL-10 enhances the proliferative capacities of HSCs. These data suggest that cellular loss induces IL-10 expression and subsequent HSC proliferation to restore hematopoietic homeostasis. In this way, rebuilding of the hematopoietic system is promoted [39].
Altogether, it appears that ILs play a major regulatory role on the HSC during hematopoiesis. Whether HSCs are maintained quiescent or whether proliferation is stimulated seems to depend, at least partly, on the presence of the different ILs.
LIF
Another cytokine expressed by osteoblasts, which has shown to be involved in hematopoiesis, is leukemia inhibitory factor (LIF) [27]. LIF is widely expressed throughout the body and it is involved in controlling self-renewal and cell fate of several precursor cell types, like MSCs, spiral–ganglion neuron precursors, and embryonic germ cells [40 –42]. LIF has shown to play a regulatory role during hematopoiesis as well. In 2008, Schraml and colleagues showed that the colony-forming ability of hematopoietic progenitor cells relies on the presence of both stromal cells and LIF. Although LIF is expressed by a wide variety of cells, these data indicate that osteoblasts can use LIF to regulate HSCs behavior [43].
Angiopoietin
To maintain their long-term repopulating (LTR) potential, stem cells are kept quiescent and divide only occasionally. In 2004, Arai and colleagues reported that quiescent LTR-HSCs localize to the bone surface and that expression of Tie-2 is distinctive for these cells [44]. At the bone surface osteoblasts expressing angiopoietin localize in close proximity to these Tie-2 + HSCs, suggesting that these cells interact through Tie-2/angiopoietin route. Subsequent in vitro studies revealed that Tie-2/angiopoietin signaling inhibits expression of genes like CXCR-4, LFA-1, α4-integrin, VEGFR-1 and VEGFR-2 involved in HSC mobilization. In contrast, the expression level of β1-integrins was up-regulated, suggesting that Tie-2/angiopoietin signaling enhances cell adhesion via up-regulation of β1-integrins and inhibition of molecules involved in mobilization [44].
Besides promoting cell adhesion, Tie-2/angiopoietin signaling consolidates the immature phenotype of HSCs. The decrease in colony-forming ability of HSCs due to long-term culturing on stromal cells was diminished when the stromal cells expressed Ang-1. Moreover, addition of exogenous Ang-1 had a positive effect on colony formation by HSCs. In line with this, the number of donor cells contributing to host hematopoiesis was significantly higher upon transplantation of HSCs cultured in the presence of Ang-1 compared with HSCs cultured without Ang-1 [44].
Label retention experiments revealed that Ang-1 inhibits HSC proliferation. The fact that a significant larger percentage of the BM HSCs resided in G0 phase after addition of Ang-1 indicated that Ang-1 signaling blocks cell cycle progression. The usage of a dominant negative form of Tie-2 confirmed that Ang-1 acts via the Tie-2 receptor [44]. Finally, Arai and colleagues successfully used Ang-1 to protect HSCs from 5-FU and X-ray treatment, which leads to apoptosis of proliferating cells. Together, these data indicate that the Tie-2/angiopoietin signaling pathway plays an important role in HSC quiescence.
Thrombopoietin
In 2007, the function of thrombopoietin (TPO), a cytokine involved in HSC maintenance and proliferation, was clarified by Qian and colleagues and Yoshihara and colleagues [45,46]. Both works revealed that TPO has role in HSC postnatal maintenance.
Qian and colleagues showed that the relative numbers of HSCs and HSPCs in the BM were age-progressively reduced in TPO−/− mice (from 2-week-old mice), the most evident decrease was found for the LT-HSCs. Moreover, when used in a competitive transplantation assay, LT-HSCs derived from TPO−/− mice showed reduced repopulating capacities in radioablated WT mice compared with LT-HSCs retrieved from WT mice. Both experiments indicate that TPO is involved in maintenance of LT-HSCs. Subsequent molecular experiments revealed that the expression levels of 2 proteins involved in cell cycle regulation, namely p57Kip2 and CDK inhibitor p19INK4D, were affected in TPO−/− mice. Moreover, BrdU incorporation kinetics in cells enriched for HSCs in TPO−/− mice was accelerated compared with WT mice [45]. Together, these results indicate that thrombopoietin and its receptor, MPL, are important in keeping HSC quiescent by regulating the expression of cell cycle regulatory proteins.
Yoshihara and colleagues [46] report the localization of MPL + quiescent LT-HSCs at the endosteal niche. Immunohistochemical analysis of the BM revealed that quiescent MPL + HSCs co-localized with TPO-expressing osteoblasts lining the bone surface. Moreover, the use of MPL-neutralizing antibodies in 2 different cell assays indicated that TPO/MPL signaling is, at least partially, responsible for the immature phenotype of HSCs.
Altogether, the data of these articles suggest that HSCs interact with endosteal osteoblasts via TPO-MPL, thereby maintaining an immature and inactive status.
SDF-1
Osteoblasts regulate HSCs by the expression of SDF-1. The SDF-1 receptor, CXCR-4, is abundantly expressed by HSCs. Thus, SDF-1 is a chemokine that acts as a chemoattractant for HSCs and its expression by osteoblasts is therefore crucial in the onset of hematopoiesis in the BM [17]. Besides its role as a chemoattractant, SDF-1 has shown to activate cell adhesion molecules on HSCs as well. SDF-1 may therefore also promote tethering of HSCs in their environment [47]. However, endosteal osteoblasts are not solely responsible for the production of SDF-1 in the BM. Endothelial cells also express SDF-1, and SDF-1 is also present in bone [17,48,49]. Moreover, Sugiyama and colleagues report a subpopulation of reticular cells that express high levels of SDF-1. These so-called CAR cells are found at the endosteal surface and near the endothelial cells [19].
Although multiple signaling molecules, which influence HSC behavior, expressed by osteoblast were identified, to pinpoint their function in vivo remains elusive. This is largely because other components of the BM, like fibroblasts and endothelial cells, are also able to provide these, or functional-related, signals, indicating that there is much redundancy in the system.
Adhesion molecules
Cell adhesion molecules (CAMs) are transmembrane proteins that mediate the direct interaction between 2 cells or a cell with the ECM. These interactions not only are crucial for cell migration and proper assembly of distinct tissues, but also allow intercellular communication.
It is thought that the localization of stem cell in their niche is partly the result of the specific expression of adhesion molecules. It has been proposed that these molecules are responsible for keeping stem cell in an environment that regulates their stemness. In the Drosophila ovary and testis, for example, CAMs are essential for germinal stem cell (GSC) maintenance [50]. Data indicate that adhesion molecules play an important role in asymmetric division of stem cells.
Several types of CAMs have been identified on HSCs. Among others, integrins, immunoglobulin superfamiliy (IgSFs), selectins, and the sialomucins elicit important roles during homing and trafficking of HSC to the endosteal niche (reviewed by ref. [51]). Moreover, there have been identified numerous adhesion molecules expressed by HSC that can interact with ligands on the osteoblast and vice versa. In addition, some authors published results that indicate that certain adhesion molecules are critical for survival of hematopoietic progenitors on osteoblasts [52]. Some in vitro models, however, suggest that cell–cell contact is not required for proper hematopoiesis [53]. So the precise role of adhesion molecules on HSCs in regulating their maintenance remains elusive.
This review will only attempt to summarize a selection of CAMs, particular those that are involved in regulating the HSC at the endosteal niche. In the next paragraphs, the evidence that has been put forward to explain the role of CAMs at the endosteum will be discussed.
Integrins
Integrins are heterodimer receptors consisting of an α-chain and a β-chain. There are numerous different α- and β-chains that can assemble, thereby creating a broad range of different integrins. These receptors interact with ligands that include ECM proteins, immunoglobulin super-family members, and the vascular cell adhesion molecule (VCAM)-1, and therefore they play an important role in cell adhesion and signaling (reviewed in ref. [54]).
Several integrins have been identified in the BM microenvironment and, more specifically, on HSCs. Expression of integrins alone does not warrant functional adhesion, since it was found that integrins did not express an activation epitope [55]. Functional integrins depend on the cytokine milieu in which they reside, making adhesive events regulatable by cytokines. Since there have been identified integrins on HSC, the role of these receptors in HSC homing is extensively studied (reviewed by ref. [56]). Among the different integrin subtypes, the β1-integrins have shown to play an important role in HSC migration and homing to the BM [57,58]. In the HSCs, the α4, α5, α6, α7, and α9 integrin subunits are expressed [59]. Clustering of the α4, α6, and α9 with a β1-subunit leads to assembly of specific receptors that are involved in HSC homing and lodgment in the BM [59,60]. The primary binding target of the α4–β1 and α6–β1 integrins on HSCs is laminin, which is found in the ECM. This explains the abrogated HSC homing capacities after addition of α4–β1 and α6–β1 blocking antibodies. Besides their role in migration, there have been identified integrins that are involved in HSC adherence to the osteoblasts. The work of Schreiber and colleagues showed that HSCs adhere to osteoblasts via the α9–β1 integrin [59]. Their binding assays revealed that the α9–β1 integrin binds to VCAM-1 and purified tenascin-C expressed by osteoblasts. Surprisingly, in contrast to data of Grassinger and colleagues [61], which will be discussed below, their data show no interaction between the α9–β1 integrin and osteopontin.
Moreover, the data of Schreiber and colleagues revealed that α9–β1 integrin antibodies are able to inhibit HSC proliferation, indicating that these molecules are able to induce signaling pathways that act on HSC maintenance. The mode of action of these antibodies is not clear yet. However, if these antibodies turn out to provide an agonistic effect on these receptors it would be interesting to look at the function of HSC-specific α9–β1 integrins in vivo. Perhaps, these integrins play a fundamental role in preserving the HSC pool at the endosteal niche.
Osteopontin
Another factor involved in HSC regulation expressed by osteoblasts is osteopontin (Opn). Opn is encoded by a single gene, but post-transcriptional modification steps, for example, phosphorylation, glycosylation, and cleavage, result in numerous different Opn subtypes. By binding to integrins, Opn modulates cell adhesion, motility, and migration. The best-known binding partners of Opn are integrins.
Although Opn is expressed not only by osteoblasts, but also by preosteoblasts, osteocytes, and hematopoietic cells, its expression in the BM is highly restricted to the endosteal surface. By attracting HSC to and maintaining them at the endosteum, Opn has shown to play an important role in stem cell homing.
Since integrins are involved in cell trafficking, it is not surprising that they were revealed as Opn-interacting proteins. Some β1-integrins expressed by HSCs have actually shown to interact with Opn [62] (reviewed in ref. [63]).
Recently, it was shown that a specific thrombin-cleaved form of Opn (trOpn) interacts with HSC via the α4–β1 and α9–β1 integrins. In addition, they show that trOpn is the predominant form present at the endosteal surface [61]. Their data revealed that this soluble form of Opn acts as a chemoattractant on HSCs, a function primarily dependent on α9–β1 integrins.
Strikingly, HSCs isolated from the endosteal surface were not inhibited in homing to their niche in Opn−/− transgenic mice, whereas HSC isolated from the central BM were [61]. These 2 HSC-subsets do so appear to express a distinctive set of chemoattractant-receptors. Although it seems that trOpn is exclusively responsible for attracting HSC originating from the central BM, other mechanisms are active in endosteal-HSCs.
Besides HSC homing, Opn has an additional role in maintaining HSCs quiescent. It has been demonstrated that Opn acts as a negative regulator of HSC proliferation in vitro as well as in vivo. In Opn−/− mice, HSCs showed an increased proliferation rate and the BM cellularity was increased [64].
Jagged–Notch signaling
The Notch signaling pathway is a highly conserved pathway involved in intercellular signaling. In the mammalian system, there have been identified 4 different receptors, Notch1–4, and 2 ligands, delta-like and Jagged. Upon ligand binding, the extracellular and cytosolic domain of the Notch receptor is cleaved and the latter translocates to the nucleus. In the nucleus, the Notch intracellular domain alters gene transcription by acting as a transcription factor. There is evidence that Notch signaling inhibits HSC differentiation in vitro and in vivo. Inhibition of differentiation is the result of symmetrically cell division and led to an expansion of the HSC pool [65]. Although osteoblasts express the Notch ligand Jagged1, the importance of the Notch signaling pathway at the endosteum is still unclear. Several works have been published with conflicting data regarding the role of Notch signaling in HSC maintenance. In 2003, Calvi and colleagues [22] report that cell–cell contact between osteoblasts and HSCs mediated by Jagged1–Notch is essential for HSC maintenance. However, subsequent studies from other groups showed that inactivation of the Notch signaling pathway had no effect on the HSC pool (extensively reviewed in [65]). These data suggest that Notch is not solely responsible for maintaining HSCs in an immature state. To what extent and under which conditions Notch signaling is important remains elusive.
N-Cadherins
The cadherin family is a diverse set of transmembrane molecules that play a pivotal role in cell–cell and cell–matrix interactions. More than 100 subtypes can be distinguished in vertebrates and invertebrates, which arise from multiple genes and post-transcriptional modifications. Although each molecule is able to establish a homophilic binding, that is, an intercellular binding between 2 adhesion molecules of the same type, they are often found in clusters. Clustering of adhesion molecules in specific organized structures, like adherence junctions and desmosomes, strongly enhances the binding strength. Cell–cell attachment through cadherins is mediated by catenins, cytosolic adaptor proteins that link the cadherins C-terminal cytosolic domain to the actin cytoskeleton. The extracellular domain of cadherins consists of multiple cadherin domains, each pair hold together by a shared Ca2 + ion. Therefore, the function of cadherins depends on the presence of extracellular calcium [66].
Several years ago, it was shown that N-cadherins are involved in HSC maintenance [21]. Upon the observation that the number of osteoblasts correlates with the number of HSCs, they investigated the expression of N-cadherin in both cell types. Histological analyses revealed that the localization of N-cadherin + osteoblasts along the endosteal surface is similar to the localization of HSCs expressing N-cadherin. Furthermore, they found that N-cadherin and β-catenin are both asymmetrically distributed throughout both cell types [21]. From these observations it was concluded that spindle-shaped bone-lining osteoblasts support HSCs through N-cadherin-mediated interactions.
Expression of N-cadherin by HSCs lining the endosteal surface was confirmed by others [67,68]. However, according to their data, these cells express only intermediate levels of N-cadherin and BM cells highly expressing N-cadherin + are not HSCs because these cells do not express the LT-HSC marker proteins. In line with this, reconstitution assays revealed that only cells with low or intermediate N-cadherin expression levels are able to fully reconstitute the hematopoietic system. HSCs that are negative for N-cadherin expression were identified as ST-HSCs. Visualization of N-cadherin expression using the LacZ reporter gene within the N-cadherin gene revealed a moderate expression of N-cadherin–LacZ in HSCs adjacent to osteoblasts with high LacZ expression. Moreover, analysis of BrdU incorporation revealed that HSCs expressing intermediate N-cadherin levels had a low cell cycle rate. These results indicated that LT-HSCs express intermediate N-cadherin levels and are kept quiescent in proximity of N-cadherin + osteoblasts at the endosteal surface [68]. Consistent with this model were the findings that up-regulation of N-cadherin in osteoblasts accompanied with inhibited HSC migration away from the endosteal niche and diminished differentiation [69].
In contrast to these studies, several articles were published that indicate that N-cadherins have no role in HSC maintenance at the endosteum [70]. Kiel and colleagues used 2 different antibodies, a monoclonal and a polyclonal, but they were unable to detect N-cadherin expression in very highly purified HSCs [70]. In addition, quantitative RT-PCR indicated that N-cadherin gene was not transcribed in HSCs either. Consistent with these findings, N-cadherin expression was not detected in HSCs isolated from gene trap mice [70].
These results seem to conflict with several other publications. Kiel and colleagues state that during earlier studies no proper HSC markers or reliable N-cadherin antibodies were used [70,71], which therefore might have led to false positive results. However, Kiel and colleagues used the same LacZ gene trap system as Haug and colleagues, nonetheless their results are contradictory. An important difference between the experimental setup used by Haug and colleagues and Kiel and colleagues is that Haug and colleagues looked at N-cadherin expression in HSCs at the endosteal niche, whereas Kiel and colleagues analyzed isolated HSCs. Since it has been shown that the expression of N-cadherin by HSCs varies and depends, among others, on external cues [44], it is possible that the loss of these extrinsic signals due to isolation of HSCs results in a decreased expression of N-cadherin. Moreover, physical stress or the use of digestion enzymes during harvesting might have an effect on the expression of cell surface molecules as well.
In order to investigate those controversies, the role of N-cadherin was studied in vivo using a mouse model, which allowed them to conditionally delete N-cadherin in HSCs [71]. Flow cytometric analysis of the BM did not show any effect on BM cellularity either immediately or 1–3 months after conditional deletion of the N-cadherin gene. Moreover, the number of HSCs and their colony-forming capacity was unaltered in N-cadherin-floxed mice [71]. In line with this, subsequent experiments revealed that N-cadherin is not required for in vivo maintenance of HSCs nor for reconstitution of the hematopoietic system of irradiated mice [71]. These data strongly suggest that N-cadherins are not required for homing and/or maintenance of HSCs at the endosteal niche.
However, the fact that these adhesion molecules are not required for HSC maintenance at the endosteal surface does not rule out that they have contributory role. After all, Arai and colleagues have shown that overexpression of N-cadherin in stromal cells enhances their capacity to nurture HSCs [44].
Moreover, the conclusions from Kiel and colleagues are based on homophilic interactions between cadherins. It has to be mentioned, however, that an other member of the cadherin family, E-cadherin, exhibits not only homophilic interactions but heterophilic interactions as well [72]. Based on the numerous articles that ascribe a role to N-cadherins in HSCs maintenance, and the multitude of adhesion molecules that can be expressed by HSCs, it might be worthwhile to address if similar heterophilic interactions, for example, involving N-cadherins and integrins, are formed between osteoblasts and HSCs.
Summarizing, the role of N-cadherins at the interface of osteoblasts and HSCs still remains debated. Although osteoblasts express high levels of these adhesion molecules, and HSCs tend to lodge near these stromal cells, the results of Kiel and colleagues put forward that N-cadherins do not influence HSC maintenance at the endosteum.
Concluding Remarks
Although the BM was first identified as a stem cell niche, the exact composition of it is still unknown. One of the problems is the identification and localization of the true HSCs in the BM. The use of stem cell characteristics, for example, LTR and label-retaining capacities, revealed several sites within the BM containing cell populations that highly exhibit stem cell properties. However, the fact that no marker that is expressed exclusively by true HSCs has been found hampers the localization of true stem cells and their direct niche. Future studies set up to identify that specific marker might therefore be very important in unraveling the composition of the stem cell niche.
To date the endosteal surface is one of the sites that accounts for a HSC niche. More specifically, the fact that osteoblasts are found in close proximity to HSCs and that they secrete factors that influence HSC behavior has led to the hypothesis that they are primarily responsible for maintenance of the HSC pool. It has to be taken into account, however, that surrounding BM cells secrete signaling molecules that act on HSCs as well.
Following clarification of certain stem cell niches in invertebrates, in which so-called hub cells are responsible for retention and nurturing of adjacent stem cells, a similar configuration has been proposed for mammalian stem cell niches. In these niche models, stem cells stay in direct contact with hub cells via adhesion molecules. It has been proposed that osteoblasts function as hub cells in the BM. Despite the identification of a broad range of adhesion molecules on osteoblasts as well as HSCs, hard evidence for requirement of direct contact between osteoblasts and HSCs is lacking. The presence of several different types of adhesion molecules in fact may be the problem in solving this issue. If these adhesion molecules work together in keeping the HSCs adjacent to osteoblasts, it could be that depletion of one of them is rescued by (enhanced) expression of others. To clarify the function of the HSC niche, not only the expression but also the functional status of adhesion molecules [73] and cross talk between cytokines and signaling by functional adhesion molecules have to be addressed in future research.
Footnotes
Acknowledgments
This study was sponsored by the Netherlands Initiative for Regenerative Medicine (NIRM) and the Dutch Program for Tissue Engineering (DPTE).
Author Disclosure Statement
No competing financial interests exist.