
Abstract
Huntington's disease (HD) is an autosomal dominant neurodegenerative disease caused by an expansion of cytosine-adenine-guanine (CAG) repeats in the Huntingtin gene Htt. To facilitate research into HD, we have derived 4 human embryonic stem cell (hESC) lines containing ≥40CAG repeats in exon 1 of Htt: SIVF017-HD (CAG40), SIVF018-HD (CAG46), SIVF020-HD (CAG48), and SIVF046-HD (CAG45). Additionally, we have derived a normal sibling-matched control for SIVF020-HD, cell line SIVF019. All 5 hESC lines had a normal karyotype, expressed pluripotency markers including Oct4, SSEA3, and Tra-1-81, and could be maintained in culture for multiple (>40) passages. Teratoma studies revealed that the hESC lines were capable of differentiating into cells representative of the 3 germ layers. Furthermore, in vitro neuronal differentiation experiments have confirmed that the hESC lines were able to generate MAP2-positive neuronal cells that express the Htt protein. Combined, these experiments confirm that the cell lines represent pluripotent stem cell lines. These HD-affected hESC lines will be made available to biomedical research laboratories and will provide a valuable tool to investigate the mechanisms and potential treatments for HD.
Introduction
H
Despite almost 2 decades of research, the normal function of the Htt protein and the pathogenesis of mutant Htt are not fully understood. The Htt protein is expressed ubiquitously in tissues throughout development, although cell loss in HD is primarily restricted to the brain. Gene ablation studies have shown that Htt is critical for early embryonic development, with constitutive knockout mice embryonic lethal prior to development of the nervous system [8,9], and plays an essential role in the postnatal brain, with conditional inactivation leading to progressive neuronal degeneration [10]. The over 200 proteins that interact with Htt suggest a role in transcriptional regulation, ubiquitin-mediated proteolysis, synapse formation, vesicle secretion, mitochondrial energy metabolism, and intracellular transport [11]. Mutant Htt also confers a toxic gain-of-function causing protease activation, protein misfolding, decreased transcription, and interrupted axonal transport [12]. At present there are no effective therapies to modulate or delay the disease, with treatment instead focused on the alleviation of symptoms.
Human embryonic stem cells (hESCs) are pluripotent cells derived from the inner cell mass of morula and blastocysts. They are capable of differentiating into cells of the 3 germ layers and as such have been proposed as a cell source for cell replacement therapies [13,14]. Additionally, hESCs have been suggested as tools for toxicity studies [15] and, when differentiated, provide a valuable in vitro model of embryonic development [16]. Disease-specific hESCs can also be used to investigate pathogenesis and facilitate drug development, which could otherwise not be performed in human patients. Recent efforts have focused on differentiation protocols to allow both investigative and therapeutic use of these cells [17]. This includes hESC differentiation to GABAergic medium spiny neurons of the striatum, the cell type primarily affected by HD [18].
Here we report the derivation of 4 HD-affected hESC lines, SIVF017-HD, SIVF018-HD, SIVF020-HD, and SIVF046-HD, as well as a normal sibling-matched control for SIVF020-HD, SIVF019. These hESC lines will provide a valuable resource to elucidate the pathophysiology of HD and to develop potential drugs to treat this fatal disease.
Materials and Methods
Embryo donation
Donated embryos were originally created by assisted reproduction technologies for the purpose of procreation and cultured to the blastocyst stage as described elsewhere [19]. Trophoectoderm biopsy and preimplantation genetic diagnosis was performed on blastocysts at day 5 or 6, to identify embryos inheriting the Htt gene mutation [20]. Twenty-nine embryos harboring the Htt gene expansion were obtained from 6 couples for stem cell derivation through an informed consent process [21]. Additionally, one sibling embryo unaffected by the Htt gene mutation but of poor quality, and therefore unsuitable for clinical use, was also declared as excess to reproductive needs and donated for investigation.
All relevant clinical procedures and research protocols were in compliance with the U.S. National Academies' 2008 Guidelines for Human Embryonic Stem Cell Research and were approved by the Sydney IVF Ethics Committee, an independent committee constituted according to the requirements of Australia's National Health and Medical Research Council (NHMRC). Cell lines SIVF017-HD, SIVF018-HD, and SIVF046-HD were derived under NHMRC license 309710. Cell lines SIVF019 and SIVF020-HD were consented for embryo cellular outgrowths for cytogenetic investigation because of unsuitability for clinical transfer and were subsequently consented for hESC line derivation, characterization, and distribution. The HD-affected hESC lines have received approval for importation into the United Kingdom from the Steering Committee for the United Kingdom Stem Cell Bank.
Stem cell derivation and culture
Derivation and culture of hESCs was performed as previously described [21,22]. Fresh or thawed embryos were plated either whole or bisected (cut into 2 parts with only the inner cell mass-containing part being plated) onto mitomycin-inactivated human fetal fibroblasts (produced in-house or commercially available CCL-110/Detroit 551; ATCC) in knockout-Dulbecco's modified Eagle's medium with 20% knockout serum replacement, 2 mM glutamine, 50 U/mL penicillin, 50 mg/mL streptomycin, 1× MEM-amino acids, 0.1 mM β-mercaptoethanol (Invitrogen), and 20 ng/mL basic fibroblast growth factor (Sigma). The embryos were incubated at 37°C in 5% CO2/6% O2 and cellular outgrowths were subsequently expanded by manual passaging and cultured as described above, with the exception that 4 ng/mL basic fibroblast growth factor was used in the culture media. The cell lines were cryopreserved at early passages using a modified vitrification procedure.
Karyotyping
Karyotyping was performed by incubation of hESC colonies with 0.22 ng/mL colcemid and 37.5 μg/mL BrdU for 17–19 h, followed by dissociation using Non-enzymatic Cell Dissociation Solution (Sigma). Metaphase spreads with G-banding were prepared using standard methodologies and analyzed by Sydney Genetics (a subsidiary of Sydney IVF; Sydney, Australia).
Molecular characterization
Identification of biopsied embryos harboring the Htt gene CAG expansion was performed by the Preimplantation Genetic Diagnosis Department of Sydney IVF (Sydney, Australia) via polymerase chain reaction (PCR)-based linkage analysis. The number of CAG repeats (as well as CCG repeats) in exon 1 of the Htt gene was determined by PCR amplification (5′-CCTTCGAGTCCCTCAAGTCCTTC-3′and 5′-TGAGGCAGCAGCGGCTGT-3′) followed by sequencing (5′-TGAGGCAGCAGCGGCTGT-3′). The number of TAA repeats in the GRIK2 gene was determined in the hESC lines by PCR amplification of the 3′UTR region [5′-(Fam) CAACACCTTTTCTCTAACCCC-3′and 5′-CTCGGCCAGTTTTTACAACTTG-3′] [23] and analysis on the 3100 Genetic Analyzer and software GeneMapper v4.0, with comparison to a sample of known (sequenced) size and standards GS500ROX (ABI Prism). The S18Y polymorphism in the UCH-L1 gene was determined by PCR amplification (5′-TTGTCTCCTCTCCGCAGGTGC-3′ and 5′-ACTCAAGCTGGGGAGCGGCGT-3′) followed by sequencing (5′-AGGCAGGCGCTGGCACCG-3′). Siblingship DNA testing of SIVF019 and SIVF020-HD was performed by DNA Labs (a subsidiary of Sydney IVF; Sydney, Australia) via analysis of X chromosomal polymorphic sites (17 markers) and mitochondrial DNA sequencing. DNA profiling was performed by DNA Labs via examination of tetranucleotide short tandem repeat loci using the AmpFℓSTR Identifiler PCR Amplification Kit (Applied Biosystems). The cell lines were negative for mycoplasma, as revealed by in-house monitoring using the MycoAlert Mycoplasma Kit (Lonza), as well as independent testing by the Victorian Infectious Disease Reference Laboratory (Melbourne, Australia).
Teratoma experiments
Teratoma inductions were performed by intramuscular injection of manually or collagenase-passaged stem cell clumps (∼1 × 106 cells) into the thigh of 6–8-week-old SCID mice. The resulting tumors were harvested when ∼1.5 cm in diameter (typically 2–3 months) and histopathology conducted to identify cell lineages (Symbion Pathology; Sydney, Australia).
Neuronal differentiation
Differentiation of hESCs towards neural cell types was performed by culturing on collagen-coated plates in Sydney IVF Neural Precursor media (includes 100 ng/mL Noggin), with passaging every 7 days for 6 weeks. For immunostaining, cells were plated on poly-ornithine (Sigma) and laminin (Cultrex)-coated 96-well plates at 8,000–10,000 cells/cm2 and cultured for a further 4 days in the presence of 20 ng/mL brain-derived neurotrophic factor before being fixed with 4% paraformaldehyde/1% gluteraldehyde.
Immunostaining
Expression of pluripotency markers in cell lines was evaluated in feeder-free hESC cultures using Alexa fluorophore–conjugated antibodies Oct4 Alexa Fluor 488, SSEA4 Alexa Fluor 488, SSEA3 PE, Tra-1-60 Alexa Fluor 555, and Tra-1-81 Alexa Fluor 555 (Bioimaging Certified; BD Bioscience). Additionally, hESC lines differentiated toward neuronal lineages were stained with primary antibodies MAP2 (Sigma; detects MAP2a, 2b, and 2c) or Htt (Abcam; detects both normal and mutant Htt) combined with goat anti-mouse IgG Alexa Fluor 488 (Invitrogen) as the secondary antibody. Cultures were counterstained with DRAQ5 (Biostatus), scanned using the IN Cell Analyzer 1000, and analyzed using IN Cell Investigator 1.5 software (GE Healthcare). Expression of alkaline phosphate was examined in manually passaged hESC cultures using the Alkaline Phosphate Substrate Kit (Vector Laboratories).
Results
Derivation of HD-affected hESC lines
Twenty-nine embryos revealed via PCR-based linkage analysis to harbor the mutant Htt allele were donated by 6 families in which one parent or grandparent was known to have the HD Htt CAG expansion. Plating of these embryos generated 4 cellular outgrowths, SIVF017-HD, SIVF018-HD, SIVF020-HD, and SIVF046-HD, each of which was obtained from a different family. Additionally, a nonaffected sibling embryo of SIVF020-HD, which was deemed unsuitable for clinical use because of poor embryo quality, was plated and the outgrowth SIVF019 was generated. The 5 cellular outgrowths demonstrated compact colony morphology consistent with hESCs (Supplementary Fig. S1, available online at
Karyotyping revealed that the cellular outgrowths were cytogenetically normal at both early (≤6) and later (>p30) passages, with SIVF017-HD determined as 46-XY and lines SIVF018-HD, SIVF019, SIVF020-HD, and SIVF046-HD revealed as 46-XX (Supplementary Fig. S1). DNA fingerprinting of the cells revealed a unique and consistent DNA profile for each of the hESC lines, which can be used as a permanent “barcode” identification to monitor the use and integrity of the cells (Supplementary Table S1, available online at
Characterization of polymorphisms affecting HD in HD-affected hESC lines
Sequencing of the Htt gene CAG repeat region confirmed that the HD-affected hESC lines contained one allele with ≥40CAG repeats (Table 1). This included 40CAG repeats for SIVF017-HD, 46 for SIVF018-HD, 48 for SIVF020-HD, and 45 for SIVF046. Both Htt alleles for SIVF019 contained <25CAG repeats and was therefore considered normal. The number of CCG repeats in the Htt gene, a polymorphism 12 bp upstream of the CAG repeat, which varies in length from 7 to 12 repeats [24], was also determined by sequencing (Table 1). This CCG polymorphism does not affect HD; however, because of its proximity to the CAG repeat it is typically amplified when evaluating the CAG expansion and is therefore useful for determination. Cell lines SIVF017-HD, SIVF020-HD, and SIVF046-HD contained 7CCG repeats on both alleles, SIVF019 had 10CCG repeats on both alleles, and SIVF018-HD had one allele with 7CCG repeats (with 46CAG repeats) and the other with 10CCG repeats (with 17CAG repeats). Furthermore, we examined polymorphisms in 2 genes thought to modulate the age of onset of HD: the S18Y polymorphism in UCH-L1 and the TAA repeat polymorphism in the 3′UTR of GRIK2 (Table 1). None of the cell lines were found to contain the protective S/Y or Y/Y genotype at amino acid position 18 in UCH-L1. For the GRIK2 TAA repeat, which varies from 10 to 18 repeats and appears to be inversely correlated with age of onset, all cell lines had low-to-medium TAA repeat numbers: ≤15TAA repeats.
This table shows the number of trinucleotide CAG and CCG repeats in exon 1 of Htt, as well as the number of TAA repeats in the 3′UTR of GRIK2 and the amino acid present at position 18 (S [Serine] or Y [Tyrosine]) in UCH-L1. Note that the Htt CCG repeat does not affect HD, but is included as it is typically amplified when evaluating CAG expansion and is thus useful to ascertain.
CAG, cytosine-adenine-guanine; HD, Huntington's disease.
Pluripotency assessment of HD-affected hESC lines
The pluripotentiality of the cell lines was assessed by staining of cell cultures for hESC markers. The 4 HD-affected cell lines, as well as the unaffected SIVF019, expressed the pluripotency markers Oct4, SSEA3, SSEA4, Tra-1-60, and Tra-1-81 (Fig. 1). Additionally, all 5 lines were positive for alkaline phosphatase (Supplementary Fig. S1). Not surprisingly, differences were observed between the cell lines in growth rates and susceptibility to spontaneous differentiation during culture. We observed that SIVF019 and SIVF046-HD were the fastest growing hESC lines and least susceptible to spontaneous differentiation, SIVF018-HD and SIVF020-HD were the slowest growing and most susceptible to differentiation, and SIVF017-HD was in the middle for both growth rates and spontaneous differentiation.
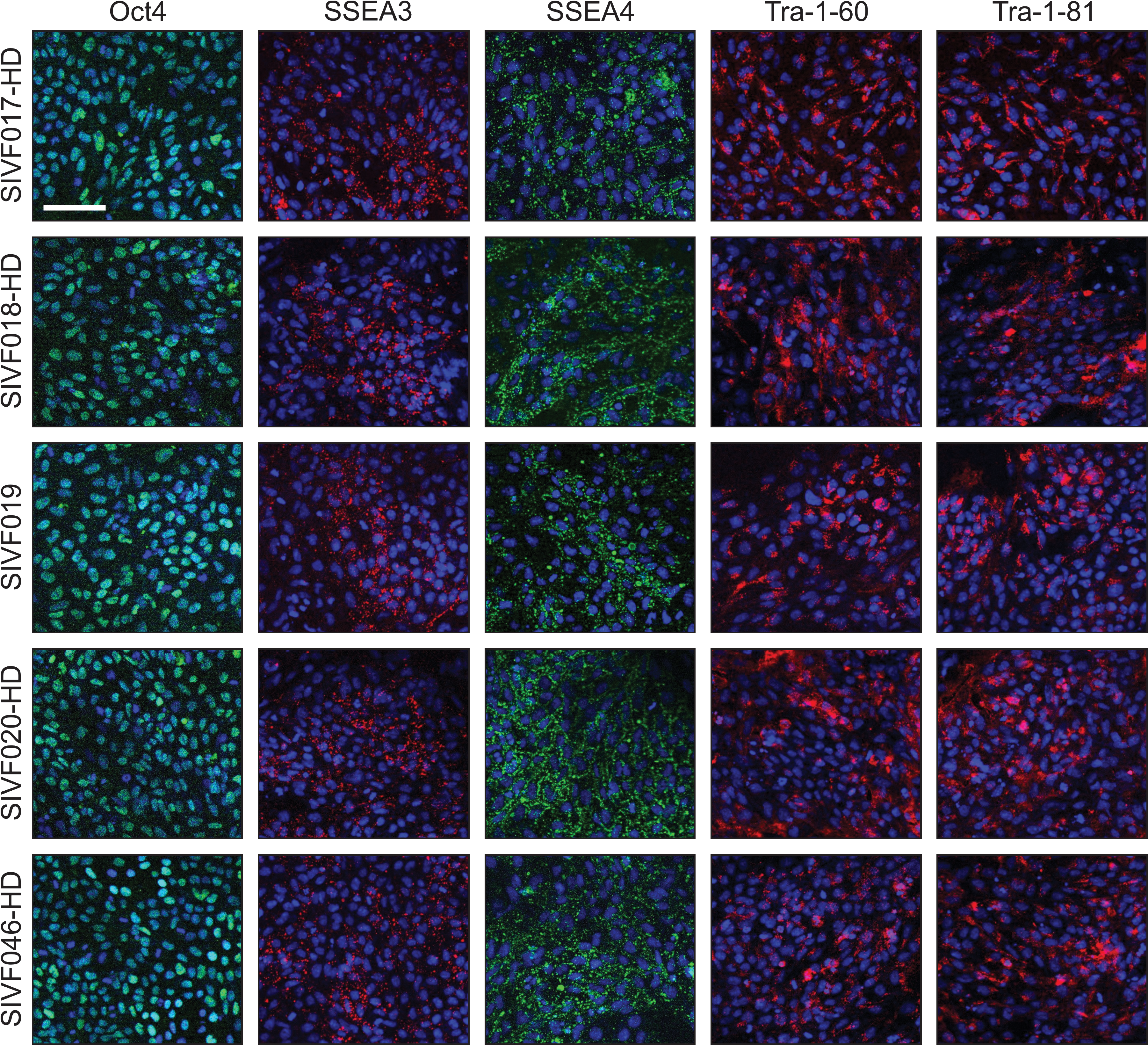
HD-affected hESC lines express pluripotency markers. Immunofluorescence staining of SIVF017-HD at p20, SIVF018-HD at p22, SIVF019 at p21, SIVF020-HD at p26, and SIVF046-HD at p7. Blue staining represents nuclear DRAQ5. Scale bar represents 100 μM. HD, Huntington's disease; hESC, human embryonic stem cell.
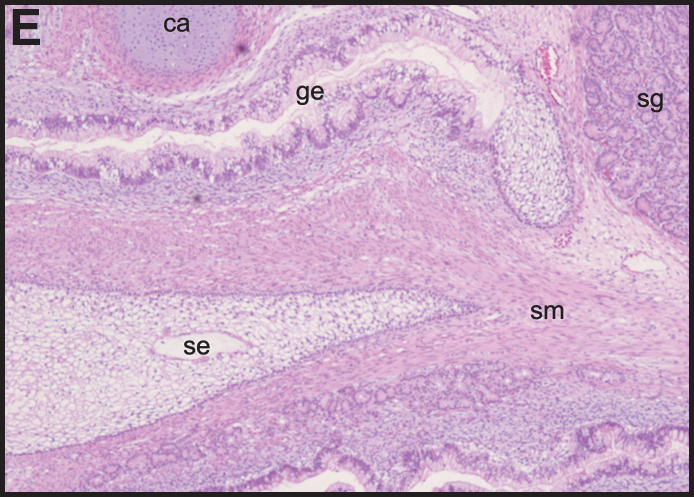
To confirm the pluripotency of the HD-affected cell lines, the potential to differentiate into the 3 germ layers was assessed by in vivo teratoma experiments. These studies identified cells representative of all germ layers in tumors from each of the 5 hESC lines (Fig. 2). This included ectodermal tissues such as immature squamous epithelium and neuroepithelium; mesodermal tissues such as cartilage, bone, smooth muscle, and adipose tissue; and endodermal tissues such as immature gastrointestinal and respiratory epithelium, and salivary glands. These experiments, combined with the expression of pluripotency markers, suggest that the hESC lines constitute pluripotent stem cells.
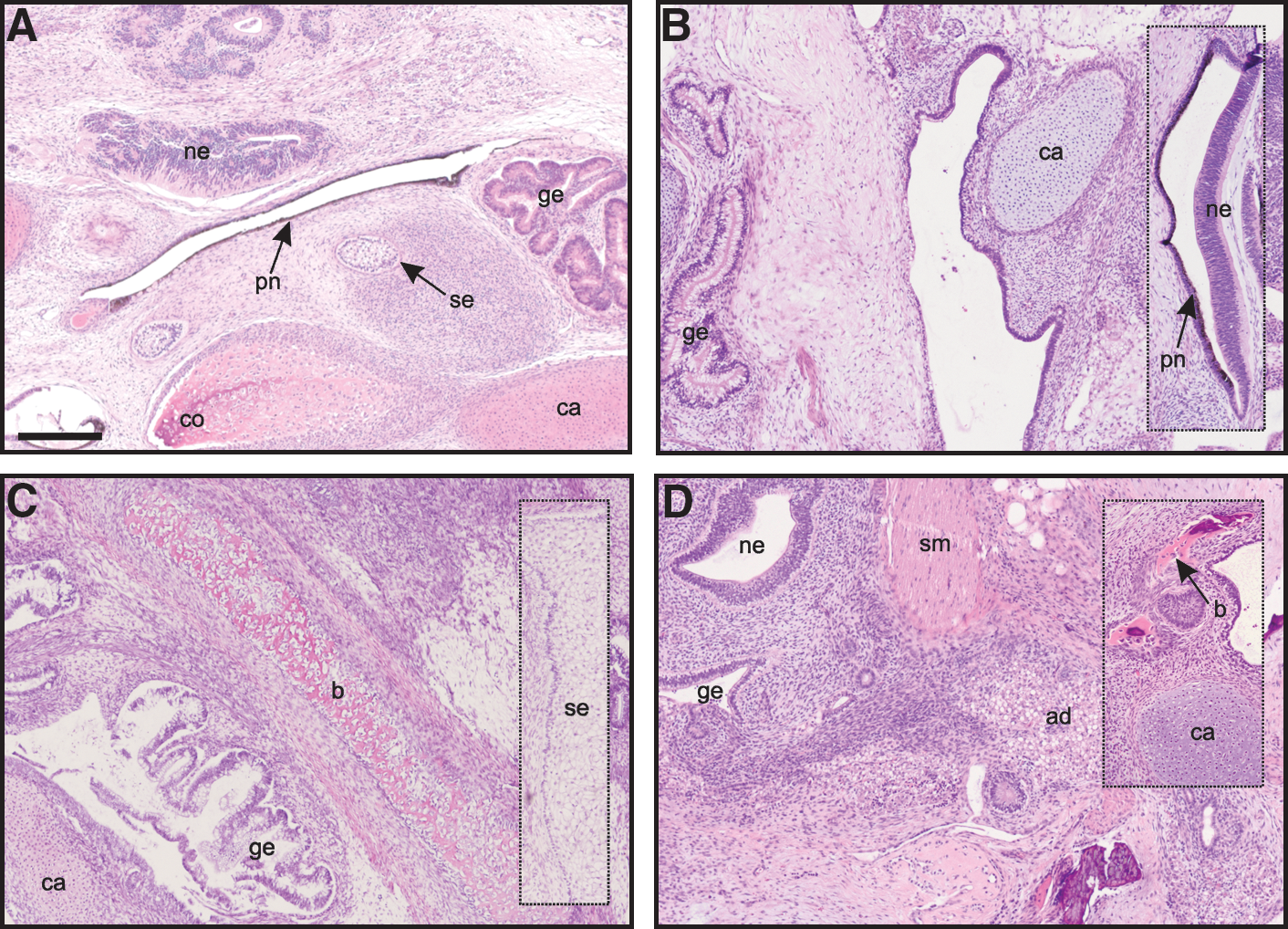
HD-affected hESC lines are capable of differentiating into cells of the 3 germ layers. Hematoxylin and eosin–stained teratoma sections from SIVF017-HD at p4 (
Neuronal differentiation of HD-affected hESC lines
To confirm that the HD-affected cell lines were capable of generating neurons, the hESC lines were differentiated to neural cell types. All of the stem cell lines generated cells with morphology typical of neurons such as neurites, expressed early neuronal markers including MAP2 (Fig. 3), and were negative for pluripotency markers Oct4 and Nanog (data not shown). Expression of the Htt protein was also examined (antibody detects both mutant and normal Htt) and was found to be present in both neural differentiated cells (Fig. 3) and undifferentiated hESCs, although the levels were greater in differentiated cells (data not shown).
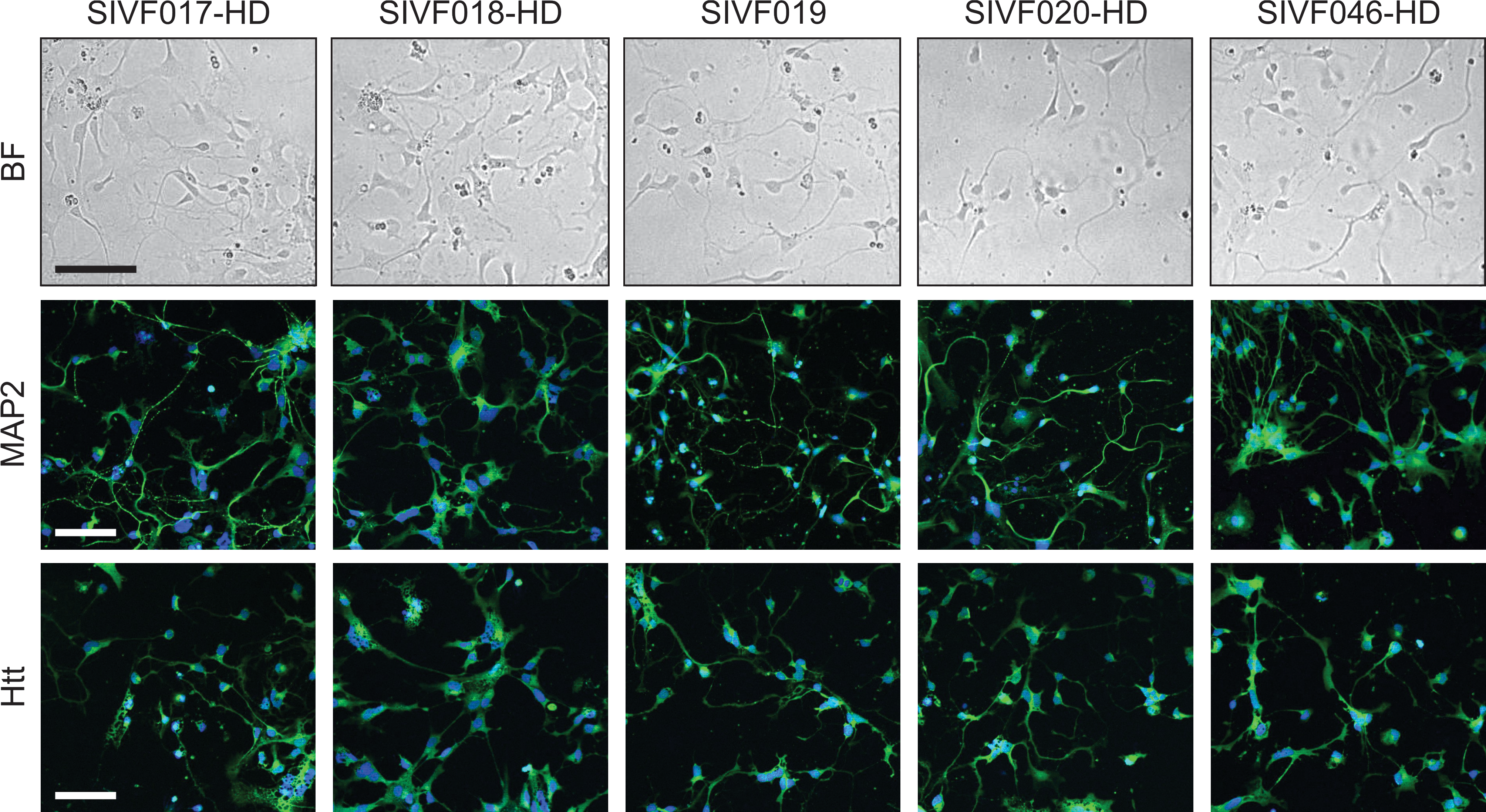
HD-affected hESC lines are capable of differentiating to neuronal lineages. Bright-field (BF) images and immunofluorescence staining with MAP2 and Htt (detects both mutant and normal Htt) of hESC lines differentiated toward neuronal cells. Blue staining represents nuclear DRAQ5. Scale bars represent 100 μM.
Discussion
The generation of therapeutically relevant compounds relies on models of neurodegenerative disease that accurately mimic neuropathology. The most commonly utilized animal models for HD are transgenic R6/1 and R6/2 mice containing 115 and 150 Htt CAG repeats, respectively [25]. Although these mice do demonstrate progressive motor and cognitive dysfunction, they do not suffer from the severe striatal atrophy paramount to the human disease [26]. Indeed, of the 4 potential therapeutics demonstrated to increase survivability in R6/2 mice that have progressed to phase II clinical trials, none have proven effective in humans [27 –30]. Because of the high cost of animal models and their unsuitability for high-throughput analysis, drug screening for HD therapeutics has mainly focused on biochemical (cell-free) assays of Htt aggregation or transgenic immortalized cell lines (reviewed in ref. [31]). Most often these cell lines show phenotypes unrelated to the tissue of interest and focus on a single aspect of a complex disease mechanism. In contrast, HD-affected hESCs theoretically provide an unlimited, uniform source of cells (including neurons) without any genetic manipulation and may more accurately reflect the human disease at a cellular level.
In this report, we describe the derivation of 4 new HD-affected hESC lines, SIVF017-HD (CAG40), SIVF018-HD (CAG46), SIVF020-HD (CAG48), and SIVF046-HD (CAG45), as well as a normal sibling-matched control for SIVF020-HD, SIVF019. Importantly, we have demonstrated that the hESC lines displayed the hallmark characteristics of pluripotent stem cells: stable growth rates beyond 10 passages, normal karyotype, expression of pluripotency markers, and the ability to form teratomas containing cells representative of all 3 germ layers. Additionally, we have demonstrated that the 5 hESC lines are capable of in vitro differentiation to neurons and will thus be a useful tool to investigate HD. The overall results of the experiments are summarized in Table 2.
Pluripotency marker expression was assessed by staining for Oct4, SSEA3, SSEA4, Tra-1-61, Tra-1-80, and alkaline phosphatase in undifferentiated hESC cultures. In vitro neural differentiation was assessed by staining for MAP2 in hESC cultures differentiated toward neuronal lineages.
hESC, human embryonic stem cell.
Each of the HD-affected hESC lines had a Htt CAG repeat of ≥40, the expansion length at which HD is completely penetrant in human patients, with a predicted age of onset of 32 years for SIVF020-HD, 36 for SIVF018-HD, 37 for SIVF046-HD, and 59 for SIVF017-HD [32]. It is however estimated that the Htt CAG repeat only accounts for 50%–70% of the variation in age on onset of the disease [33,34] and several genetic modifiers have been identified, including the size of the normal Htt CAG repeat [35]. The protective S18Y polymorphism in the neuron-specific deubiquitinating enzyme UCH-L1, which is evident in 17% of HD patients and has been attributed with 13% of the variance associated with the CAG repeat length and age of onset [36], was not present in any of the HD-affected cell lines. Additionally, we studied the 3′ UTR TAA repeat polymorphism of the excitatory neurotransmitter receptor GRIK2, which is thought to inversely correlate with age of onset and account for 7%–13% of age of onset variation [34,36]. All cell lines were found to be in the medium repeat range for this polymorphism and thus it is not likely to significantly alter their theoretical age of onset.
Four other hESC lines have been derived from HD-affected embryos, SI-186 (CAG37; partially penetrant), SI-194 (a sibling of hESC-186; CAG not documented), SI-187 (CAG51) [37,38], and VUB05_HD (CAG44) [39], although only limited characterization of the first 3 lines was reported. Cell line SI-186 and SI-187 have subsequently been differentiated in vitro to cells that express early neuronal markers, with the authors claiming to demonstrate an expansion of the CAG repeat for the partially penetrant line SI-186 in 2 of 6 small pools of neurosphere cells [38]. Unfortunately, the cell lines reported in ref. [37] have recently been rejected for U.S. federal funding by the NIH because of ethical concerns over the consent form signed by donating couples [40]. An induced pluripotency stem cell (iPSC) line with 72CAG repeats in the Htt gene has also been generated by reprogramming of a HD patient's skin cells [41]. It should be kept in mind that it is not clear if iPSC (even those made without viruses) are indistinguishable from ESCs [42 –44], and therefore, it is essential to have both iPSCs and hESCs available for research. Moreover, as hESCs (and presumably iPSCs) show a large degree of variability between different lines, including their propensity to differentiate into specific lineages [45], it will be important to have multiple HD-affected cell lines available to select the most suitable for disease modeling. These differences between stem cell lines should also be kept in mind when comparing HD-affected hESCs to nonaffected hESCs.
The differentiation of HD-relevant cell types from HD-affected hESCs should allow the exploration of HD pathology as it develops, rather than drawing conclusions from the state of brains that have run the course of the disease, as in autopsy studies, or from animal models that do not develop the key elements of HD. A recently published method for the generation of medium spiny neurons [18], the cells most prone to neurodegeneration in HD, could be combined with HD-affected hESC lines to study Htt protein aggregation and clearance, mitochondrial dysfunction, oxidative stress susceptibility, and excitotoxicity. The ability to generate mixed glial and neuronal cocultures from differentiated HD-affected and control hESC lines could shed light into the contribution of various cell types to HD pathology. Additionally, in vitro differentiation of HD-affected hESCs would be a unique tool to study the increased neural progenitor cell proliferation observed in HD-affected brains [46], as well as the cellular mechanisms by which postmitotic Htt CAG expansion occurs in neurons from HD-affected brain regions. Subsequently, any HD-related phenotype could be investigated for therapeutic intervention using hESC-based high-throughput drug screening assays.
In summary, we have derived 4 HD-affected hESC lines, which have a CAG repeat expansion in the Htt gene of ≥40 repeats, as well as a sibling-matched control hESC line. These HD-affected cell lines will allow further investigation of the pathogenesis of HD, as well as tools for HD drug screening and development, which could otherwise not be performed in human subjects. The hESC lines have been fully consented by donating couples, with no restrictions placed on their use, and will be made available to the research community along with any requested control hESC line (
Footnotes
Acknowledgments
The authors thank Prof. Peter Russell for histological examination of teratomas, Warwick Greville for molecular analysis of GRIK2 and UCH-L1 polymorphisms, and Paulette Barahona for preliminary Htt CAG analysis. This research was supported by the HP Therapeutics Foundation, Inc.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.