
Abstract
Mesenchymal stem cells (MSCs) are promising for cell therapy and regenerative medicine; but their lack of specific markers renders the cell culture at potential contamination risk with other cell types, in particular, fibroblasts. In this study, we mapped 2 differential transcriptome data of MSCs compared, one to mononuclear cells and the other to fibroblasts, onto the membrane proteome data, the analysis of which led to an identification of transmembrane 4 L6 family member 1 (TM4SF1) as a surface protein marker candidate that could discriminate MSCs simultaneously from blood cells and fibroblasts. Our analyses confirmed that TM4SF1 was abundantly expressed on MSCs but neither on other blood/tissue cells nor on fibroblasts. TM4SF1 immunoselection from bone marrow and adipose tissues yielded homogeneous cell populations that were highly similar to MSCs, in terms of morphology, immunophenotype, and differentiation potential. These findings indicate that TM4SF1 can serve as a surface protein marker which singly identifies MSCs from diverse cell sources, in particular, fibroblast-rich connective tissues.
Introduction
A
Over the past decades, advances in hybridoma technology have expedited the development of a series of MSC-reactive monoclonal antibodies, namely SH-2, SH-3, SH-4, SB-10, HOP-26, and Stro-1 [8 –12], which have turned later out to recognize several different surface proteins on MSCs, including CD105, CD73, CD166, and CD63 [13 –15]. Recently, 3 glycolipids, stage specific embryonic antigen (SSEA)-1 [16], SSEA-4 [17], and disialoganglioside (GD2) [18] as well as 3 transmembrane proteins, nerve growth factor receptor [19], CD200 [20], and fibroblast activation protein α (FAPα) [21] have been discovered as MSC-specific surface markers and proved effective in identification and enrichment of MSCs from bone marrow (BM). However, it remains largely unknown whether all these molecules can equally serve as MSC-specific markers in other tissue sources and, more importantly, whether they can reliably discriminate MSCs from their phenotypic twin cells, that is, fibroblasts.
Tetraspanins are a large family of membrane glycoproteins that display a characteristic structural feature made of 4 transmembrane domains. There is a separate group of tetraspanins, called a transmembrane 4 L6 family (TM4SF), that share limited sequence homology with other family members [22]. They are known to induce formation of membrane microdomains and play diverse roles in various blood cells (for review, see ref. [23]); but little is known about their expression and roles in MSCs, except the role of CD9 in cell proliferation and attachment [24].
In this study, we performed a combined analysis on previously obtained MSCs' membrane proteome data [25] and differential transcriptome data [26,27], in an attempt to discover novel surface protein markers that could simultaneously distinguish MSCs from blood cells and fibroblasts. Such surface markers would facilitate identification and enrichment of MSCs from diverse cell sources, in particular, fibroblast-rich connective tissues.
Materials and Methods
Cell preparation
Cryopreserved human BM-derived mononuclear cells (MNCs), BM-derived MSCs, adipose tissue (AT)-derived MSCs, and dermal fibroblasts were purchased from Lonza Inc. Cryopreserved AT-derived stromal vascular fraction (SVF) samples as well as umbilical cord blood (UCB)-derived MNCs and MSCs were obtained from local companies with informed written consents. Both MSCs and fibroblasts were cultivated as previously reported [27]. A hybridoma cell line (L6-20-4) producing anti-TM4SF1 antibody was a generous gift from Prof. Steve R. Roffler (Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan).
Flow cytometry
All analyses were performed on a Guava PCA-96 flow cytometer (Guava Technlogies), with mouse monoclonal antibodies against previously established MSC markers (Table 1) and Alexa532-conjugated goat anti-mouse IgG (Invitrogen). All measurements were analyzed with Guava Express software and De novo software.
Reverse transcriptase-polymerase chain reaction analysis
Human multiple tissue cDNA panels were obtained from Clonetech Laboratories Inc. For other cell samples, total RNA was extracted using Trizol Reagent and reverse-transcribed to cDNA, as previously reported [25]. Primer sets and amplification conditions for the genes were listed in Table 2.
BSP, bone sialoprotein; RUNX2, runt-related transcription factor 2; aP2, adipocyte fatty acid binding protein; PPAR-γ2, peroxisome proliferator-activated receptor γ2; COLX, collagen X; SOX9, Sry-box 9; ACTB, β-actin.
Omics data manipulation
The membrane proteome data of MSCs [25] consisting of 191 ectodomain-containing membrane-bound proteins were integrated with 2 differential transcriptome datasets of MSCs [26,27] and reduced into a subset of 148 different proteins (Supplemental Table S1, available online at
Fluorescence cell labeling
For efficient fluorescence labeling and tracing of MSCs, we used Cell Staker (Biterials Co.,
Immunoselection
About 0.5 μg of L6-20-4 monoclonal antibodies purified from the supernatant of hybridoma cultures were incubated with 25 mL (1 × 107) of Dynabeads Pan mouse IgG (Dynal Biotech) for 30 min. After removal of excess antibody, the coated beads were then mixed with 2.5 × 107 thawed BM-derived MNCs or AT-derived SVFs, incubated for 20 min at 4°C, and separated by applying external magnets.
In vitro differentiation assays
TM4SF1+ cells were in vitro expanded to a second passage and incubated with mesenchymal differentiation media (Lonza, Inc.). Their differentiation into osteogenic, adipogenic, and chondrogenic lineages were measured by von Kossa, Oil Red O, and Alcian Blue staining, respectively.
In vivo osteogenic differentiation assay
The 0.4 μm-pore size transwell inserts (Corning, Inc.) were seeded with about 1 × 104 cells, sealed with parafilm, and subcutaneously implanted for 3 weeks in nude mice. After the inserts were harvested, their membranes were detached and stained by von Kossa method.
Results and Discussion
We first compared the expression pattern between MSCs and fibroblasts using previously established MSC markers (Table 1). The flow cytometry analysis showed that MSCs were strongly positive for CD105, CD73, CD166, CD63, SSEA-4, GD2, and FAPα and weakly positive for CD49a and CD146 (Fig. 1A), thereby certifying individual or combinatorial uses of these surface markers for positive identification of MSCs. Unfortunately, however, most of these markers were found to be expressed also on skin fibroblasts, mostly at comparable levels to those on MSCs, indicating that these markers might be unsuitable for discriminating unequivocally between MSCs and fibroblasts. Exceptionally, GD2 appeared to be differentially expressed in MSCs compared with fibroblasts, implying its potential use to discriminate MSCs from fibroblasts. However, its highly dispersed and heterogeneous expression pattern, which was also shown in a previous report [18], would limit the widespread use in basic and clinical research.
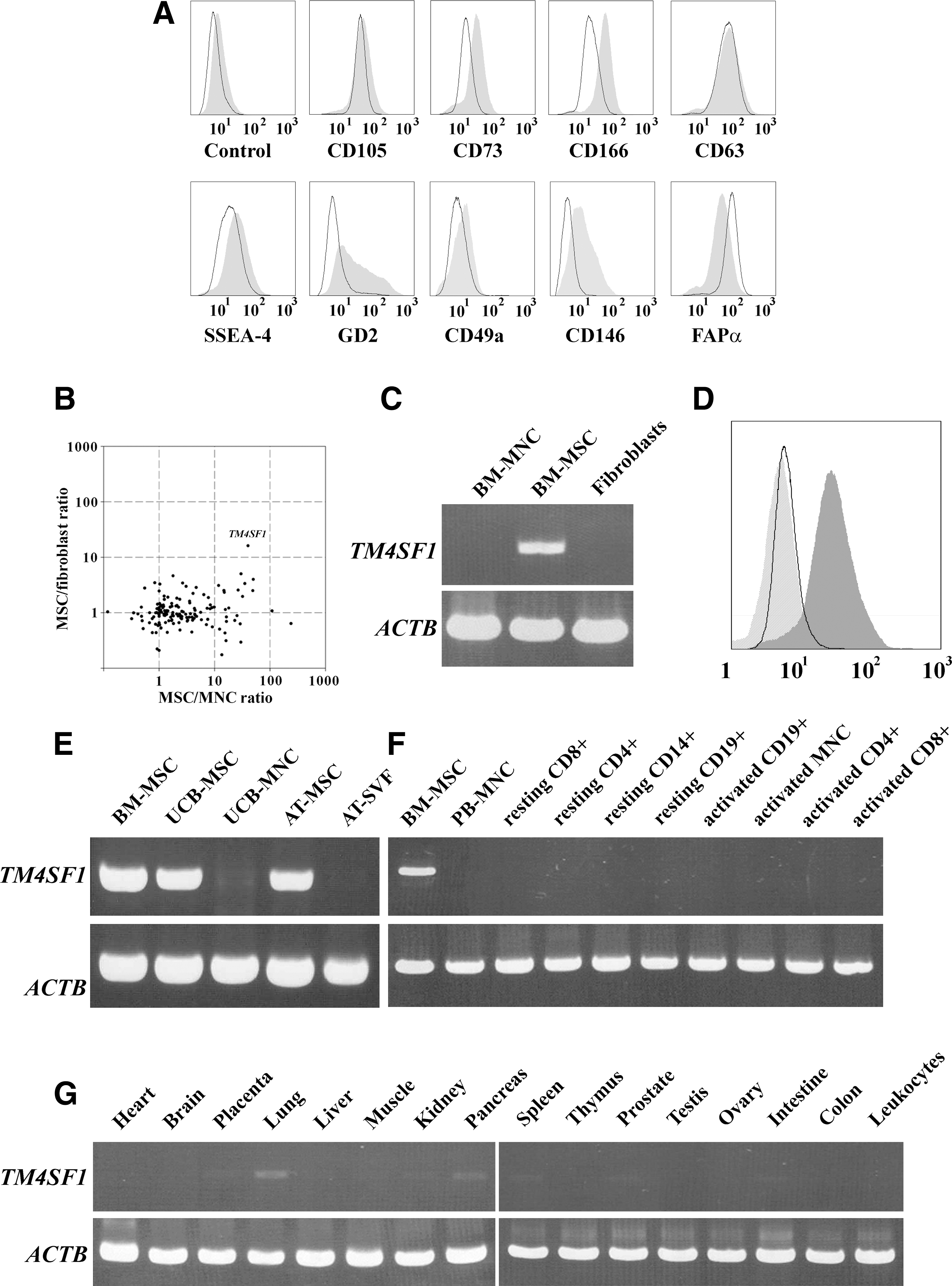
Identification of TM4SF1 as a novel MSC-specific surface marker.
Therefore, of practical value for safe and effective clinical applications of MSCs would be novel surface markers that could discriminate reliably MSCs not only from other blood cells but also from fibroblasts. To discover such doubly discriminating surface markers, we devised a combined omics approach, in which the membrane proteome data of MSCs [25] were integrated with 2 differential transcriptome datasets of MSCs, 1 against BM-derived MNCs [26] and the other against fibroblasts [27], and reduced to a subset of membrane proteins whose gene expression intensity ratios in MSCs compared with MNCs and fibroblasts could be mapped (Supplemental Table S1). A 2D scatter plot analysis of the combined data revealed TM4SF1, variously known as H-L6, L6, M3S1, or TAAL6, as a leading surface marker candidate that could simultaneously distinguish MSCs from MNCs and fibroblasts, because its gene expression was observed to be uniquely up-regulated at 10-fold or higher levels in MSCs compared with the other 2 cell populations (Fig. 1B).
To verify this finding, we compared TM4SF1 expression among BM-derived MSCs, MNCs, and fibroblasts at both gene and protein levels. The reverse transcriptase–polymerase chain reaction (PCR) followed by flow cytometry analyses showed that TM4SF1 was abundantly expressed on MSCs but neither on fibroblasts nor on MNCs (Fig. 1C, D), indicating that TM4SF1 expression might be exclusive to MSCs within BM stroma. Likewise, TM4SF1 expression also appeared to be highly confined to a MSC type within 2 other important sources for MSCs, that is, UCB and AT, because a high level TM4SF1 expression was observed in MSCs derived from UCB and AT but not in those tissues per se (Fig. 1E) where MSCs were very rare in number.
Then, we examined TM4SF1 expression in major body tissues by PCR screening of human multiple tissue cDNA panels. The results showed that TM4SF1 was not expressed at any significant level in either peripheral blood fractions (Fig. 1F) or major tissues (Fig. 1G). These findings were in line with previous reports that TM4SF1 expression, albeit often up-regulated in some carcinomas [29], was suppressed to a very low level in most of normal tissues [30]. Therefore, TM4SF1 is likely to serve as a surface protein marker that can uniquely identify MSCs from most of tissue sources, including fibroblast-rich connective tissues.
To prove such a possibility, we attempted to isolate MSCs from cryopreserved BM samples by immunoselection with TM4SF1-targeting antibodies. For this study, we used an L6-20-4 antibody [31], whose high specificity and affinity toward the ectodomain of TM4SF1 was demonstrated in our feasibility test with a cell mixture of TM4SF1+ MSCs and TM4SF1− fibroblasts (Supplementary Fig. S1, available online at
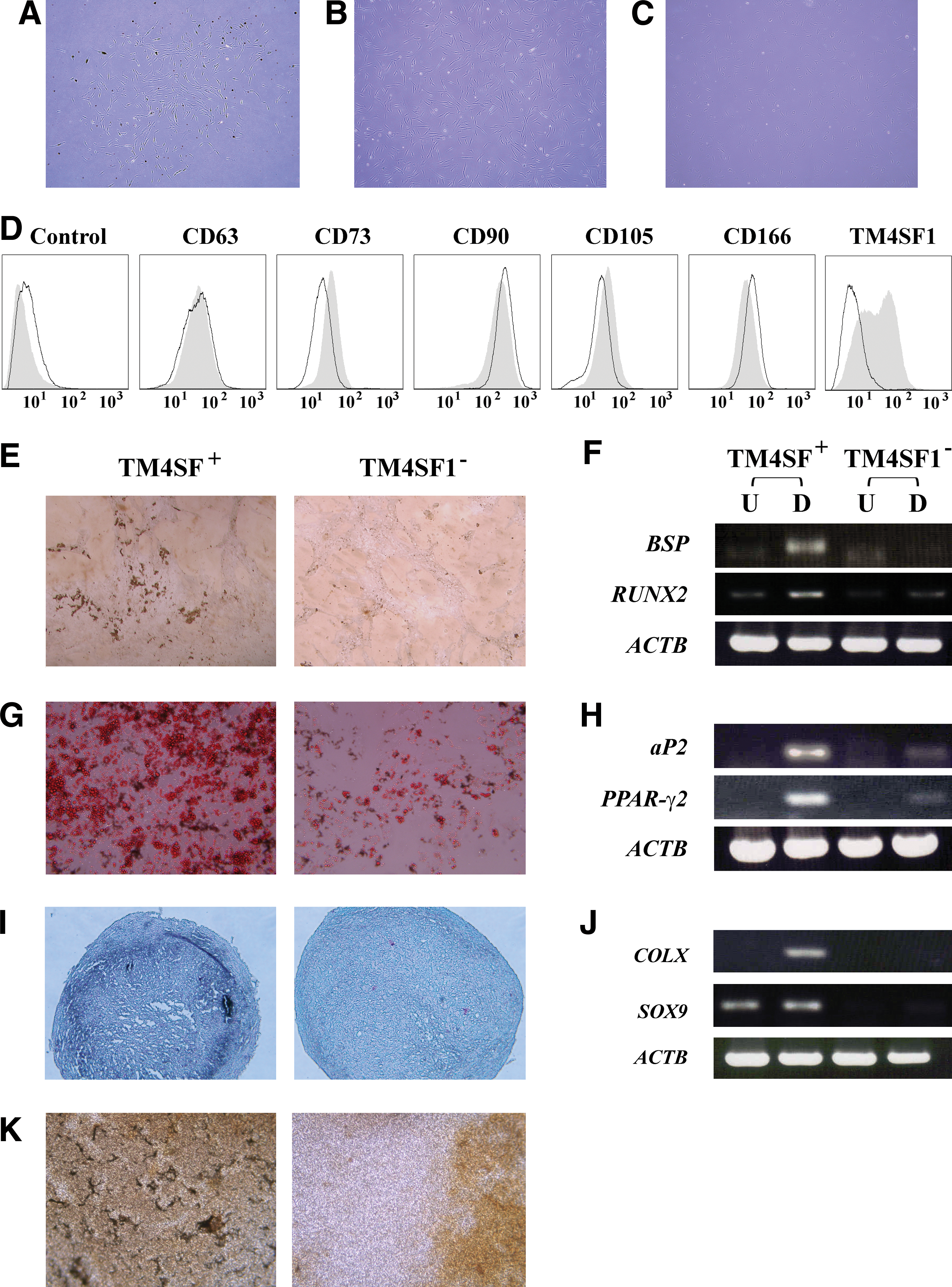
Comparative analyses of morphology, immunophenotype, and mesengenic differentiation potential between TM4SF1+ and TM4SF1− cells
Next, to demonstrate that TM4SF1 could serve as a surface marker discriminating MSCs simultaneously from fibroblasts and MNCs, we repeated immunoselection with a cryopreserved SVF sample, one of fibroblast-rich cell sources [32]. The results showed that both TM4SF1+ and TM4SF1− fractions yielded fibroblastic cells with frequencies of about 1 to 10 in 1 × 104 cryopreserved SVF cells. On continuous cultivation, TM4SF1+ cells yielded a homogenous fibroblastic cell population (Fig. 2B), whereas the TM4SF1− cells gave rise to heterogeneous cell types (Fig. 2C). Although TM4SF1− cells exhibited a slightly higher growth rate than TM4SF1+ cells (Supplementary Fig. S2, available online at
To further investigate the nature of TM4SF1+ and TM4SF1− cells, we incubated them separately under mesengenic differentiation conditions. Cytochemical staining and PCR analyses showed that TM4SF1+ cells, but not TM4SF1− cells, could readily differentiate into osteoblasts, adipoblasts, and chondroblasts (Fig. 2E–J; for MSC control, Supplementary Fig. S3, available online at
In the future, TM4SF1, due to its doubly discriminative nature against fibroblasts and blood cells, would be popularly used for identification and enrichment processes of MSCs from diverse cell sources, in particular, fibroblast-rich connective tissues.
Footnotes
Acknowledgments
This research was supported in part by a Korea Science and Engineering Foundation grant (M10641000037) funded by the Ministry of Education, Science, and Technology and in part by the grant (SC-2140) from Stem Cell Research Center of the 21st Century Frontier Research Program, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.