
Abstract
Adult mesenchymal stem cells (MSCs) have been shown to spontaneously express cardiac proteins (CP) in vitro and to improve cardiac function after transplantation into experimentally induced acute myocardial infarction (AMI). However, if these effects are the result of MSC cardiac differentiation or a mere cooperative cellular interaction is a matter of active debate. Additionally, the molecular mechanisms involved in CP expression by adult stem cells in vitro and its possible benefit for cardiac regeneration and improved function remain unclear. Here we show that although MSCs effectively engraft in AMI tissue, this engraftment leads to downregulation of CP expression in the implanted MSCs. We also found that pretransplantation cardiac specification of MSCs by exposure of the cells to transforming growth factor beta 1 (TGF-β1) led to sustained MSC CP expression without altering engraftment efficiency. This increase in CP expression was associated with greater improvement in cardiac function 1 and 4 weeks after AMI with TGF-β1-pretreated MSCs. We discovered that the TGF-β1-enhanced cardiac potential of MSCs was mediated by downregulation of disabled-2 (Dab2) expression, suggesting an inverse correlation between Dab2 levels and CP expression/cardiac functional improvement after MSC engraftment. Our investigations further demonstrate that loss of Dab2 expression was sufficient to induce MSC CP expression and improve cardiac function after MSC engraftment after AMI. In summary, we define a novel role for the TGF-β1 receptor adaptor protein Dab2 as a regulator of CP expression in MSCs and its potential as a molecular target for the enhancement of stem cell cardiac specification for transplantation therapies.
Introduction
A
What is well known is that modulation of cardiac tissue repair after myocardial infarction (MI) can reduce ventricular remodeling [8 –10]. In this context, stem cell transplantation represents a promising therapeutic alternative to help minimize myocardial cell loss and optimize remodeling after MI. In experimental settings the use of stem cells from embryonic, fetal, and adult origins for cardiac tissue repair has been reported in mice and rat MI models [7,10 –12]. Most of these studies describe the ability of stem cells to survive, engraft, and, to varying degrees, improve heart function after transplantation. Nevertheless, the level of tissue recovery achieved by exogenous progenitors varies greatly depending on the source of stem cells, and several possibilities are being studied to enhance engraftment and function of these cells [8,13 –15]. A reasonable alternative to improve stem cell engraftment, survival, and functional differentiation is to direct the cells to the desired lineage in vitro before transplantation [16,17].
In vitro stem cell differentiation often requires stimulation with drugs, specific growth factors, or cytokines to direct the cells to a particular phenotype. Several growth factors have been described to play relevant roles in cardiac specification of progenitor cells during development and stem cells in a variety of study models [18,19 –23]. The intracellular signaling pathways activated by these growth factors, the regulatory mechanisms, and the interactions between them are under active investigation. We have focused our efforts to improve cardiac specification of stem cells on the adaptor protein disabled-2 (Dab2) as a mediator of the signaling induced by transforming growth factor beta (TGF-β). TGF-β family of proteins has been shown to participate in cardiac development [18] as well as cardiac myocyte differentiation in vitro [19 –23].
Interestingly, TGF-β and disabled-2 have been shown to mediate cell lineage specification. For example, TGF-β1 treatment of epithelial cells leads to an upregulation of Dab2 critical for mesenchymal transformation [24]. In addition, during heart development TGF-β1 regulates the epithelial to mesenchymal transformation essential for formation of the heart valves and septum [25,26].
In our studies with mesenchymal stem cells (MSCs), we observed that treatment of MSCs with TGF-β1 induced a rapid decrease in Dab2 expression, and this effect induced marked increases in cardiac protein (CP) expression in vitro and in vivo after transplantation into infarcted myocardium. Sustained expression of CPs by engrafted MSCs was associated with greater improvement in cardiac function. Importantly, we demonstrate that Dab2 is a key molecular regulator involved not only in cardiac specification of MSCs but also in the functional consequences of MSC engraftment after acute MI.
Materials and Methods
Animals
All animals were housed in the AAALAC animal facility of the Cleveland Clinic Foundation and maintained under standard conditions. This investigation conforms to the Guide for the Care and Use of Laboratory Animals published by the National Institutes of Health (NIH publication No. 85–23, Revised 1996) and was approved by the IACUC of the Cleveland Clinic Foundation.
MSC preparation
MSCs were isolated from adult Lewis rat bone marrow as we have described previously [8,27]. In brief, after anesthesia with a mixture of ketamine (100 mg/kg) and xylazine (5 mg/kg) (IP), femurs were isolated and flushed with 0.6 mL Dulbecco's Modified Eagle Medium (DMEM) (Gibco–Invitrogen). The collected clumps of bone marrow were gently dissociated with a 20-gauge needle. The cell suspension was separated by a Percoll density gradient, centrifuged for 10 min at 260 g, and washed 3 times with phosphate-buffered saline (PBS) supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin (Invitrogen). The cells were then re-suspended and plated in DMEM-low glucose (Gibco–Invitrogen) with 10% fetal bovine serum and 1% antibiotic and antimycotic (Gibco–Invitrogen) and maintained at 37°C. Nonadherent cells were removed by replacing the medium after 3 days. Cultures were fed every 3–4 days until 70% cell confluence was reached. At this point, adherent cells were detached by incubation with 0.05% trypsin and 2 mM EDTA (Invitrogen) for 5 min at 37°C and subsequently re-plated. As we have done in previous studies, we validated the MSC phenotype and stability of the MSC phenotype through in vitro differentiation of the MSC preparation into adipocytes, chondrocytes, or osteoblasts [8]. We validated MSC phenotype as late as passage 16 [8].
Surface antigen detection and further characterization of MSCs
To prevent nonspecific selection of monocytes and macrophages, MSC cultures were immunodepleted of CD45+, CD34+ cells by negative selection using 10 μL each of primary PE-conjugated mouse anti-rat CD45 (BD Biosciences), and CD34 antibodies (Santa Cruz Biotechnology, Inc.) using the EasySep PE selection kit according to the manufacturer's instruction (Stem Cell Technologies). The MSCs were then tested by fluorescence-activated cell sorting (FACS) and were positive for CD90 and CD29 and negative for CD34 and CD45. The multipotentiality of resulting cells was verified with the use of in vitro assays to differentiate MSCs into osteogenic (alkaline phosphatase activity), adipogenic (oil red O staining), and chondrogenic (Alcian Blue) lineages according to published protocols [28].
Antibodies
In addition to those described above, additional antibodies used in our studies for western blot, immunocytochemistry, and immunohistochemistry analysis were Anti-TGF-β1 (5 ng/mL; Biovision); monoclonal primary antibody to α-sarcomeric actinin (Sigma); mouse anti-troponin I (Chemicon International, Inc.); mouse monoclonal anti-α-sarcomeric actinin (Sigma); rabbit polyclonal anti-Dab2 (1:1,000; BD Transduction Laboratories); mouse IgG1monoclonal anti-Akt1 (1:500; Cell Signaling Technology); rabbit anti-GATA-4 polyclonal IgG antibody (Santa Cruz Biotechnology); goat polyclonal anti-Nkx-2.5 IgG antibody (Santa Cruz Biotechnology); rabbit polyclonal anti-MEF-2 IgG antibody (Santa Cruz Biotechnology); and rabbit anti-connexin-43 polyclonal IgG antibody (Santa Cruz Biotechnology). Primary antibody detection was carried out using Alexa Fluor–conjugated secondary antibodies goat anti-rabbit, donkey anti-mouse, or donkey anti-rat (Gibco)
Immunocytochemistry
MSCs were fixed for 30 min with 4% paraformaldehyde in PBS and after washing 3 times with PBS, permeabilized with 0.1% Triton for 15 min. Nonspecific binding was inhibited with 3% bovine serum albumin in PBS for 1 h. Immunostainings were performed including incubations with primary antibodies only and secondary antibodies only to determine if there was any nonspecific binding and levels of autofluorescence. In the case of the antibodies selected there was no significant nonspecific binding in vitro or in vivo. Primary antibodies were incubated at the indicated concentrations overnight at 4°C. After 2 washes with PBS, the cells were incubated with secondary antibody for 1 h at room temperature. Fluorescence staining was observed using a upright spectral laser scanning confocal microscope (TCS-SP; Leica Microsystems) equipped with blue argon (for DAPI), green argon (for Alexa Fluor 488), and red krypton (for Alexa Fluor 594) lasers. Image processing and analysis were evaluated using Leica Confocal software. Optical sectioning was averaged over 4 frames and the image data set was 1,024 × 1,024 pixels.
Immunohistochemistry
Animals were sacrificed 1 and 4 weeks after MI. Tissues were fixed in formalin and embedded in paraffin blocks according to established protocols. Antigen was retrieved using 10 mM sodium citrate buffer (pH 6.0) and heat at 95°C for 5 min. The buffer was replaced with fresh buffer and reheated for an additional 5 min, and then cooled for 20 min. The slides were washed in deionized water 3 times for 2 min each. Specimens were then incubated with 1% normal blocking serum in PBS for 60 min to suppress nonspecific binding of IgG. Slides were incubated for 1 h at room temperature or overnight with the indicated primary antibodies. Optimal antibody concentration was determined by titration. Slides were then washed PBS and incubated for 45 min, with secondary antibodies (Alexa Fluor) diluted to 1.5 μg/mL in PBS with serum in a dark chamber. After washing extensively with PBS, coverslips were mounted with aqueous mounting medium (Vectashield Mounting Medium with DAPI, H-1200; Vector Laboratories). Tissues were analyzed using an upright spectral laser scanning confocal microscope and 15 random fields of the infarct border zone were obtained and saved by a blinded observer. The number of GFP+ MSCs and the number of GFP+ and cardiac marker-positive GFP+ cells was counted by independent observers blinded to treatment group.
Histological analysis
Seven days after MI/cell transplantation, the hearts were excised and divided into 3 transverse slices with mid section being the basal portion of the infarct scar. Collagen deposition was evaluated in 5 μm sections after Masson's trichrome stain as described previously [29,30] by blinded observers to the individual samples. Seven images of tissue whole cross of the left ventricle were acquired using an ArtixScan 4000tf (Microtek) slide scanner with PathScan Enabler (Meyer Instruments) at a resolution of 4,000 dpi. ImagePro Plus (Media Cybernetics) was used to quantify percentage of myocardial area that exhibited collagen deposition. Percent area of collagen deposition (%) is expressed as a percentage of total left ventricular (LV) area by cross section.
Flow cytometry
MSCs were trypsinized and then washed once in PBS supplemented with 1% bovine serum albumin and 0.1% sodium azide (FACS buffer). After permeabilization with 0.1% triton in PBS for 10 min, the cells were incubated with Fc-receptor blocker for 10 min and incubated 15 min on ice with specific primary antibodies as indicated. After washing the cells once again, they were incubated with secondary antibody conjugated with Alexa Fluor 498 or 633 (Molecular probes) for 30 min in the dark at 4°C. At this point the cells were washed, resuspended in FACS buffer, and analyzed using FACSCalibur and CellQuest software (BD Biosciences). Negative controls consisted of either isotype-specific IgG or Alexa Fluor 488– or 633–conjugated goat anti-mouse control in the absence of primary antibody.
Western blot analysis
After the treatments, the cells were scraped from the culture dishes, pelleted, and gently washed with ice-cold PBS. Cells were lysed by adding prewarmed (95°C) 125 mM Tris and 1% sodium dodecyl sulfate (SDS; pH 6.8) buffer to the cell pellets. Cell lysates were then centrifuged and the supernatant was used as whole protein cell lysate. After the proteins were electrophoretically separated in 10% SDS polyacrylamide gels and electrotransferred to blotting polyvinylidene fluoride (PVDF) membranes, the unspecific bonds were blocked with 5% skimmed milk in 1× TBST [25 mM Tris (pH 8.0), 125 mM NaCl, and 1% Tween 20] for 1 h at room temperature and then probed with primary antibodies over night at 4°C. After incubation with horseradish peroxidase-conjugated anti-mouse or anti rabbit secondary antibodies (1:5,000–1:10,000, 1 h, room temperature), antibody recognition was observed with chemiluminescence kit (Amersham Biosciences) according to manufacturer's instructions. Alternatively, secondary antibodies conjugated with IRDye (Li-COR/Odyssey) were used and immunoblot was detected with Odyssey infrared scanner following manufacturer's instructions.
Genomic DNA extraction and real-time polymerase chain reaction
Genomic DNA was isolated by incubating the tissue in a DNA extraction buffer [50 mM Tris (pH 8.0), 1 mM EDTA, 100 mM NaCl, 0.5% SDS, and 500 μg/mL preoteinase K] at 55°C overnight on a heater shaker. Next day, 700 μL of a combination of 25:24:1 phenol/choroform/isoamyl alcohol was added and the soluble phase collected after centrifugation (14,000 rpm, 10 min). An additional 700 μL of chloroform was added to this solution, mixed by inverting the tubes, and spun down (14,000 rpm, 10 min). Approximately 500 mL of the supernantant from the tubes containing the DNA was then mixed with NaCl at a final concentration of 0.25 M and subsequently precipitated with 500 μL of isopropyl alcohol. After centrifugation for 10 min DNA pellet was washed with 70% ethanol and resulpend in 100 μL TE buffer (10 mM Tris and 1 mM EDTA, pH 8.0).
Real-time polymerase chain reaction (RT-PCR) was performed as detailed below using primers to the male-specific Y-chromosome Sry3 gene in rats, previously used to quantify the number of male cells engrafted in females for Sry3 [31]: forward 5′-GCATTTATGGTGTGGTCCCGCGG-3′ and reverse 5′-GGCACTTTAACCCTTCGATGAGGC-3′. The cycling conditions were 10 min at 95°C for activation of polymerase, and then 30 s at 95°C for denaturation, 60 s at 62°C inducing annealing, and 30 s at 72°C for extension for a total of 45 cycles as previously described [6]. For loading controls we used 18S: Hs99999901 TaqMan®Gene Expression (Applied Biosystem).
Extraction of RNA and quantitative RT-PCR of Dab2
RT-PCR was performed after isolation of RNA from 6 × 10 6 MSCs using Rneasy Mini Kit (Qiagen, Inc.) according to manufacturer's instructions. Quantitative RT-PCR was performed by using the ABI Prism 7700 sequence detector (Applied Biosystems). The reaction mixture contained SYBR Green PCR master mix (Applied Biosystems), each primer at 300 nM, and 10 μL of cDNA. After activation of the AmpliTaq Gold (Applied Biosystems) for 10 min at 95°C, 45 cycles were performed with each cycle consisting of 15 s at 95°C followed by 1 min at 60°C. The dissociation curve for each amplification was analyzed to confirm that there were no nonspecific PCR products. GAPDH was used as internal control and to determine Dab2 relative expression. Primers used for Dab2 were forward 5′-GCT ATA ACA GGG C AA CAGG-3′ and reverse 5′-GTT CTG ATT GGT GTC GAT TTCA-3′; those used for GAPDH were forward 5′-TAC GAC AGG CTG GTA TCA TTGG-3′ and reverse 5′-ATC GAA GTC GTA CTG GATC-3′.
Dab2 gene silencing
Transfection was performed by electroporation with the Amaxa system according to the manufacturer's instructions for MSCs (Amaxa). Cells (3 × 106) were transfected in each experiment with a transfection efficiency of 55%–70% as assessed by GFP expression. To achieve effective inhibition of Dab2 expression, a combination of 3 Dab2 siRNAs was used (NCBI accession no. NM_024159).
Sequence 1 (5′ → 3′): sense GGAUUCUAUGAUGAAACUCtt; antisense GAGUUUCAUCAUAGAAUCCtg
Sequence 2 (5′ → 3′): sense GCACCAUCAAAGAAGGAAAtt; antisense UUUCCUUCUUUGAUGGUGCtt
Sequence 3 (5′ → 3′): sense GGUGAUGGUGUAAAAUACAtt; antisense UGUAUUUUACACCAUCACCtt
Left anterior descending ligation
Ligation of the left anterior descending (LAD) artery in Lewis rat was performed as previously described [8,29,32]. Briefly, animals were anesthetized with intraperitoneal ketamine and xylazine, intubated, and ventilated with room air at 80 breaths/min using a pressure-cycled rodent ventilator (RSP1002; Kent Scientific Corporation). Anterior wall MI was induced by direct ligation of the LAD artery with the aid of a surgical microscope (M500; Leica™ Microsystems). MSC transplantation was performed immediately after MI as described previously [8,9]. In brief, 2 × 106 MSCs were collected by trypsinization of the cell culture flask at P10, rinsed with PBS, and resuspended in 200 μL PBS. Cells were then injected into the infarct border zone multiple sizes.
GFP labeling of cells
We implemented a VSV-G pseudotyped lentivirus expressing EGFP or Dab2. The lentivirus was made using 4 plasmid vector system. The MSCs were transduced twice for 8 h with purified lentivirus in the presence of 8 μg/mL of polybrene at a multiplicity of infection of 30. The medium was changed 72 h posttransfection and replaced with the regular medium containing zeocin (EGFP), or zeocin and blasticidin (hSDF1 and EGFP). Thus, only cells that have incorporated the viral genome including the zeocin and/or blasticidin resistance gene survived.
Echocardiography
Two-dimensional (2D) echocardiography was performed at 7 and 28 days after LAD ligation and MSC transplantation using a GE Vivid 7 with a il3L linear probe operated at 14 MHz [8,32,33]. LV dimensions and wall thickness were quantified by digitally recorded 2D clips and M-mode images in the parasternal short- and long-axis views from the mid-LV just below the papillary muscles to allow for consistent measurements from the same anatomical location in different rats. LV-end diastolic dimension (LVEDD) and LV- end systolic dimensions (LVESD) were obtained from the short-axis views from the mid-LV just below the papillary muscles. Ejection fraction was calculated by quantifying the LV area at end systole and end diastole from the parasternal long view with the apex fully observed. Ejection fraction was calculated as follows: (left ventricular end diastolic area [LVEDa] − left ventricular end systolic area [LVESa])/LVEDa ×100. Measurements in each animal were made 6 times from 3 out of 5 randomly chosen 2D clips that contained at least 3 beats per clip that were recorded by an observer blinded to the treatment arm.
Statistical analysis
Data are presented as mean ± standard deviation. Comparisons between groups were by unpaired Student's t-test or by ANOVA with Bonferroni correction for multiple comparisons where appropriate.
Results
MSC CP expression is regulated in vitro as a function of passage
To determine MSC cardiac potential we derived MSC cultures from 6 individual Lewis rats [8,29,34] and assessed expression of relevant cardiac skeletal proteins [ventricular myosin heavy-chain (α,β-MHC) and troponin I] and cardiac-related transcription factors (GATA-4 and MEF-2). We found that each MSC culture exhibited a well-defined CP expression pattern, which in some cases showed distinct variations as a function of passage (P). For instance, α,β-MHC expression was detected at low levels at P4, increased between P8 and P23 (80% ± 7.5% of total cells), and was not expressed beyond P37. Troponin I, a cardiac and skeletal muscle protein indispensable for muscle contraction, was present in all passages being detected in 75% ± 8.0% of the total cells (Fig. 1). The transcription factor GATA-4 was predominantly expressed between P10 and P26 (90% ± 5.0% of the total cells), showing cytoplasmic localization with increased nuclear translocation with passage that peaked at P18. MEF-2, a transcription factor involved in muscle formation during development, also showed peak expression around P18 decreasing after P20 to undetectable levels (Fig. 1). We also analyzed the expression pattern of the gap junction protein connexin-43, which belongs to a family of proteins implicated in intercellular connections that are important for myocardial tissue architecture and function. Connexin-43 was detected at P10, subsequently showing low but stable expression in MSCs even at late passages (Fig. 1).

Mesenchymal stem cell (MSC) expression of cardiac protein (CP) varies relative to in vitro passage. MSCs showed expression of the indicated CPs, cardiac-related transcription factors, and gap junction protein at different levels of expression and with noticeable variation according to the passage (P). α,β-Myosin heavy-chain (MHC) and troponin I were highly expressed and were detectable throughout the culture. Conexin-43 (CX-43), Gata-4, and MEF-2 showed increased expression around P18. The arrow in the figures for Gata-4 and MEF-2 identify examples of nuclear expression of the transcription factor.
TGF-β1 treatment of MSCs increases expression of cardiac-specific cytoskeletal proteins, transcription factors, and gap junction proteins in vitro
According to our data and those from others [27], MSC CP expression presents great variability depending on several factors related to their maintenance in vitro. These characteristics diminish the potential of the cells to effectively integrate into the host tissue and result in poor functionality after transplantation. An approach to maintain a sustained CP expression in MSCs in vitro before their implementation for transplantation in injured hearts was therefore investigated. To this end, we relied on the known cardiogenic effects of TGF-β1 on stem cells [21,31,35,36]. After harvesting and culturing for 16–18 passages, MSCs were exposed to 5 ng/mL TGF-β1 for different periods and the effects of treatment on CP expression were evaluated. We observed that the treatment of MSCs with TGF-β1 was associated with a significant increase in expression of a number of structural CP, transcription factors, and gap junction proteins. The transcription factors GATA-4, GATA-5, MEF-2, and Nkx2.5, and the cardiac-associated structural CP α,β-MHC and troponin I, as well as the gap junction protein conexin-43, showed a significant upregulation (Fig. 2A, B). The effect of TGF-β1 on MSC CP expression was observed as early as 24 h after treatment.
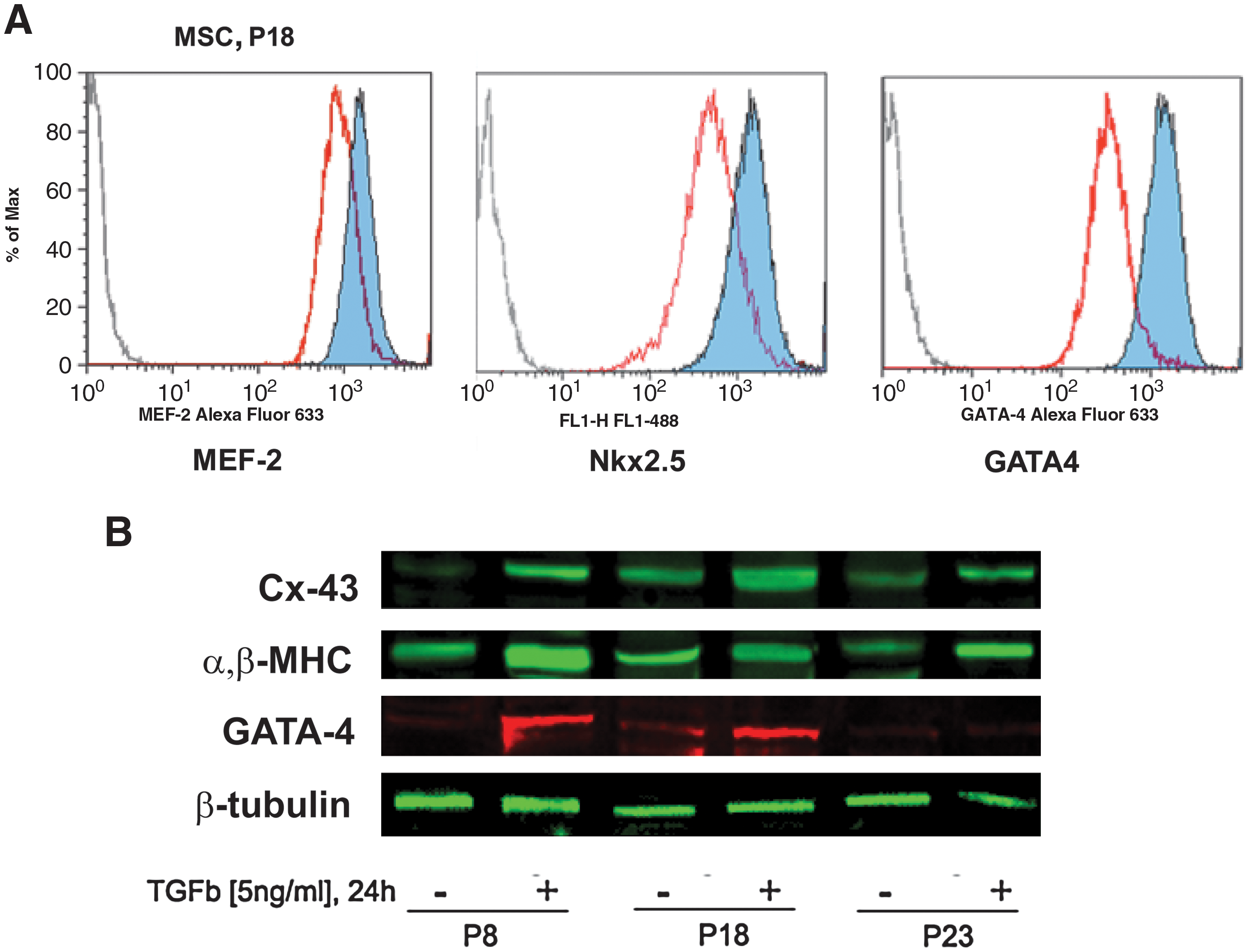
Transforming growth factor beta (TGF-β) treatment increases CP expression in MSCs. MSCs maintained in vitro during different passages were exposed to TGF-β (24 h, 5 ng/mL). Expression of the cardiac-related transcription factors MEF-2, NKx2.5, and GATA-4 was analyzed by flow cytometry
Role of TGF-β receptor adaptor proteins in MSC CP expression
To determine the molecular mechanisms activated by TGF-β1 in MSCs that led to the enhancement of CP expression in vitro, we analyzed expression of signaling molecules modulated by TGF-β1. TGF-β1 signals by activating the cell surface serine/threonine kinase receptors TGF-βRI and TGF-βRII. These receptors phosphorylate Smad proteins, leading to the activation and nuclear translocation of transcription factors. Additionally, TGF-βRI/II-activated intracellular signaling may be regulated by a series of adaptor proteins such as Dab2 or Smad-anchor for receptor activation. While we did not detect significant changes in expression of smad proteins or TGF-β receptors, we did observe a dramatic effect of TGF-β1 on expression of the adaptor protein Dab2. Expression of Dab2 was downregulated by TGF-β1 and this effect was observed through all the passages analyzed (Fig. 3A). MSC Dab2 expression was downregulated as a function of passage, and thus MSC Dab2 expression appears to be inversely correlated with expression of CPs independent of TGF-β treatment. Downregulation of Dab2 was rapid, with a significant decrease observed as early as 1 h after the addition of TGF-β1 and sustained at least up until 48 h (Fig. 3C). Further, this effect appears to be specific to TGF-β1 as it was not observed in response to other growth factors (bone morphogenetic protein-2 and fibroblast growth factor-2) and drugs (retinoic acid) that may impact cardiac specification in MSCs (Fig. 3D).
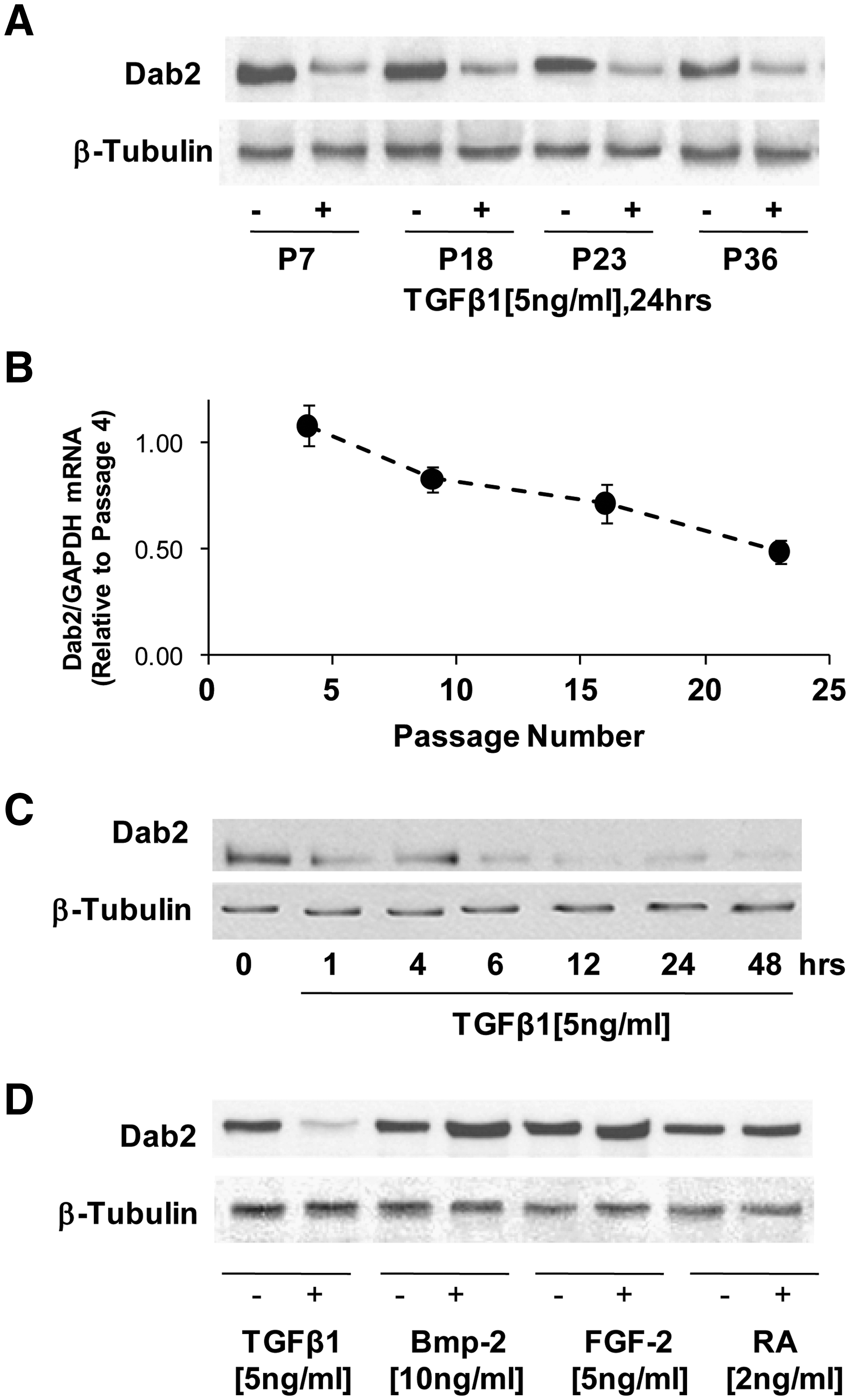
TGF-β specifically induces downregulation of expression of the adaptor protein disabled-2 (Dab2).
On the basis of these results, we hypothesized that Dab2 downregulation was required for CP expression in MSCs. To test this hypothesis, we knocked down Dab2 expression in MSCs by siRNA-mediated silencing and examined CP expression by quantitative RT-PCR. As shown in Fig. 4, effective abrogation of Dab2 mRNA and protein expression was achieved in MSCs transfected with a combination of 3 siRNA sequences targeting Dab2 mRNA (Fig. 4A). We observed a significant increase in GATA-4 mRNA expression in cells in which Dab2 expression was attenuated (Fig. 4B). GATA-4 mRNA levels reached expression levels similar to those induced by the treatment with TGF-β1 (Fig. 4B). The importance of Dab2 in CP expression in MSCs was further confirmed by increased expression of α-MHC mRNA in MSCs transfected with Dab2siRNA (Fig. 4C). No significant effect on Dab2, GATA-4, or α,β-MHC expression was observed in cells transfected with a scrambled siRNA sequence, demonstrating the specificity of the method. In addition, we found that overexpression of Dab2 in MSCs results in downregulation in expression of α,β-MHC (Fig. 4D). In MSCs in the presence of Dab2 overexpression, TGF-β fails to induce α,β-MHC expression (Fig. 4D). These results demonstrate that Dab2 expression is central in regulating CP expression in MSCs.
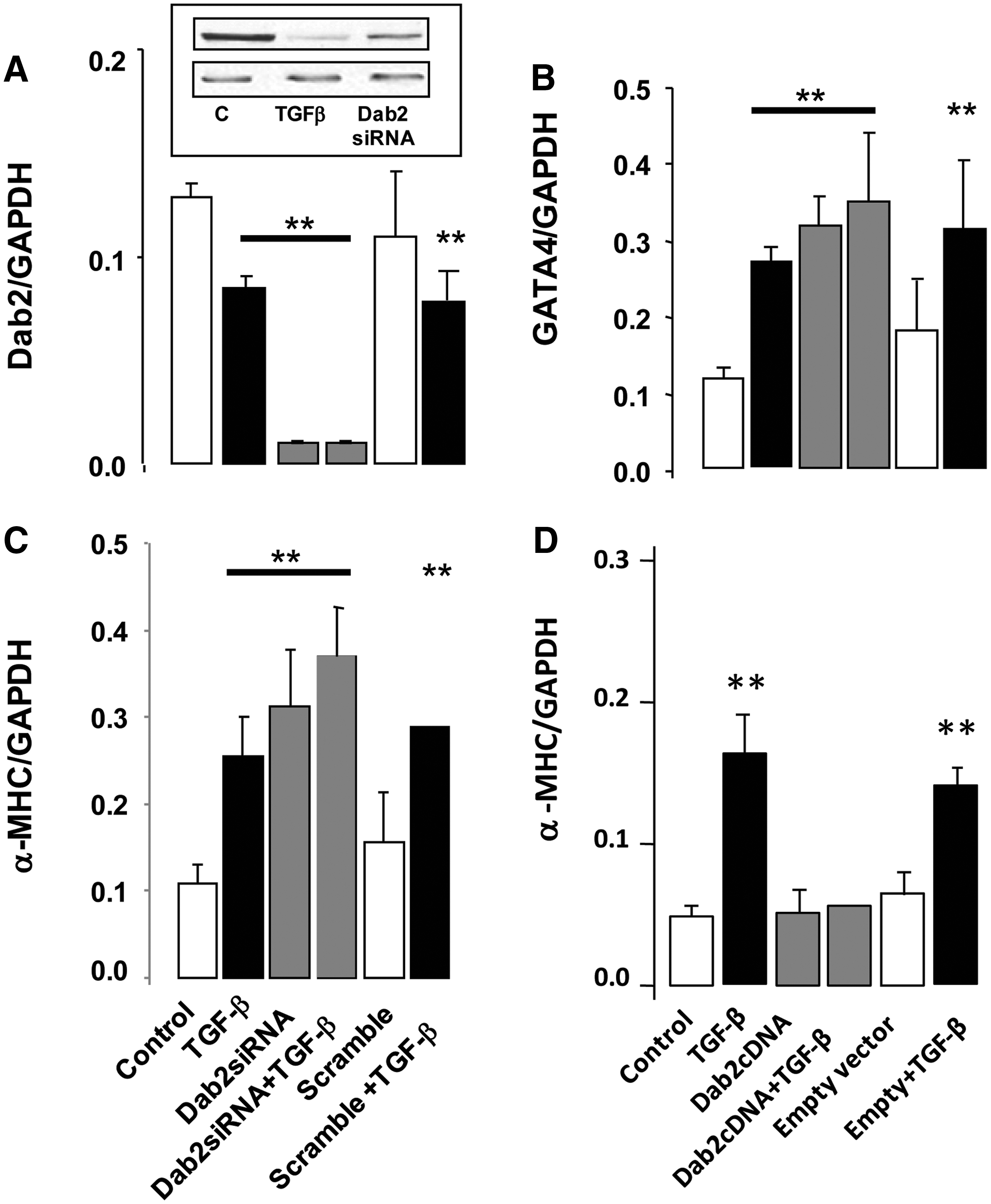
Dab2 expression regulates CP expression in MSCs. MSCs were transfected with 3 different Dab2 siRNA sequences to assure effective Dab2 repression that was confirmed by quantitative polymerase chain reaction
TGF-β1-pretreated MSCs survive, engraft, and sustain CP expression, and improve heart function after transplantation into infarcted myocardium
Our in vitro data demonstrate that MSCs spontaneously express cardiac lineage proteins in a passage-dependent manner, that cardiac commitment is enhanced by treatment with TGF-β1, and that Dab2 plays a critical role in the cardiogenic potential of these cells. We hypothesized that MSCs pretreated with TGF-β1 and/or Dab2 siRNA would maintain their CP expression profile after transplantation into infarcted myocardium. We further postulated that upregulation of MSC expression of Dab2 would have negative consequences on MSC protein expression and the functional consequences after transplantation after MI. Thus, we treated stably expressing GFP MSCs with TGF-β1 (24 h, 5 ng/mL) and transplanted 2 × 106 cells at passage 18 into the infarct border zone immediately after MI induced by LAD artery ligation (LAD; see Materials and Methods). We then quantified the number of engrafted cells and analyzed their CP expression pattern by fluorescence microscopy 7 days after transplantation (Fig. 5A–D). Additionally, to assess the functional impact of MSC transplantation we performed a parallel experiment in which we also injected untreated, TGF-β1-treated Dab2:siRNA cells (+/− TGF-β1) and Dab2:cDNA cells, and performed echocardiography before and immediately after the infarction in separate groups of animals at 7 and 28 days postinfarct/MSC transplantation (see Materials and Methods; Fig. 5 and Table 1). We observed GFP-positive cells in the infarct border zone surrounding necrotic tissue in the hearts injected with control and manipulated MSCs 7 days after LAD ligation (Fig. 5A, B). There was no difference in the degree of MSC engraftment as measured by GFP+ cells per unit area across groups with an average of 201.8 ± 20.9 MSC/mm2. After injection into the infarct border zone, MSCs appeared to migrate toward the ischemic myocardium and engrafted in proximity with native cardiac myocytes. This cellular response seems to be unique to the microenvironment of the ischemic myocardium because we did not observe any evidence of transplanted MSCs at remote, uninjured areas of the infarcted hearts.
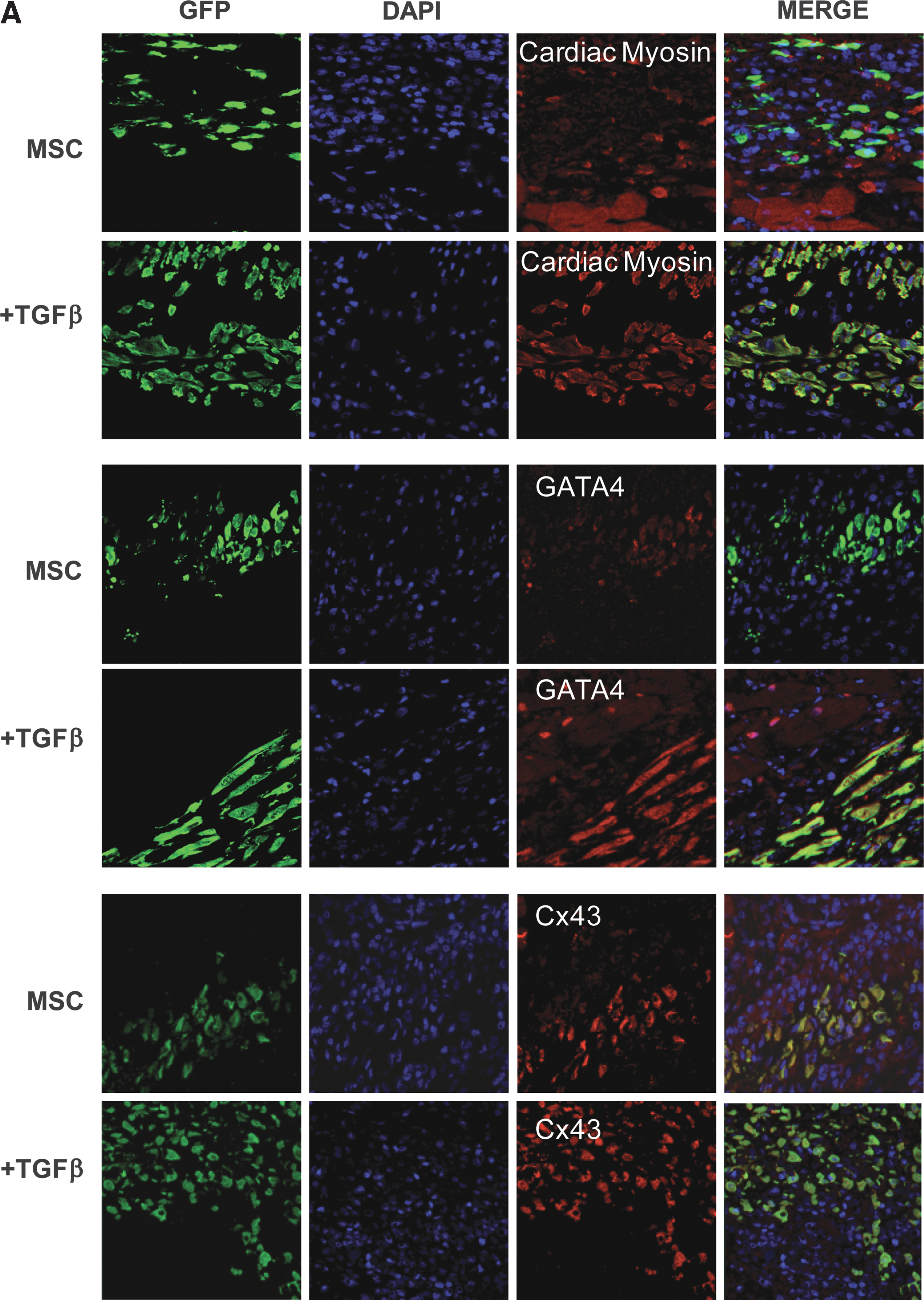
Inhibition of Dab2 expression enhances MSC-mediated improvement in cardiac function after transplantation.
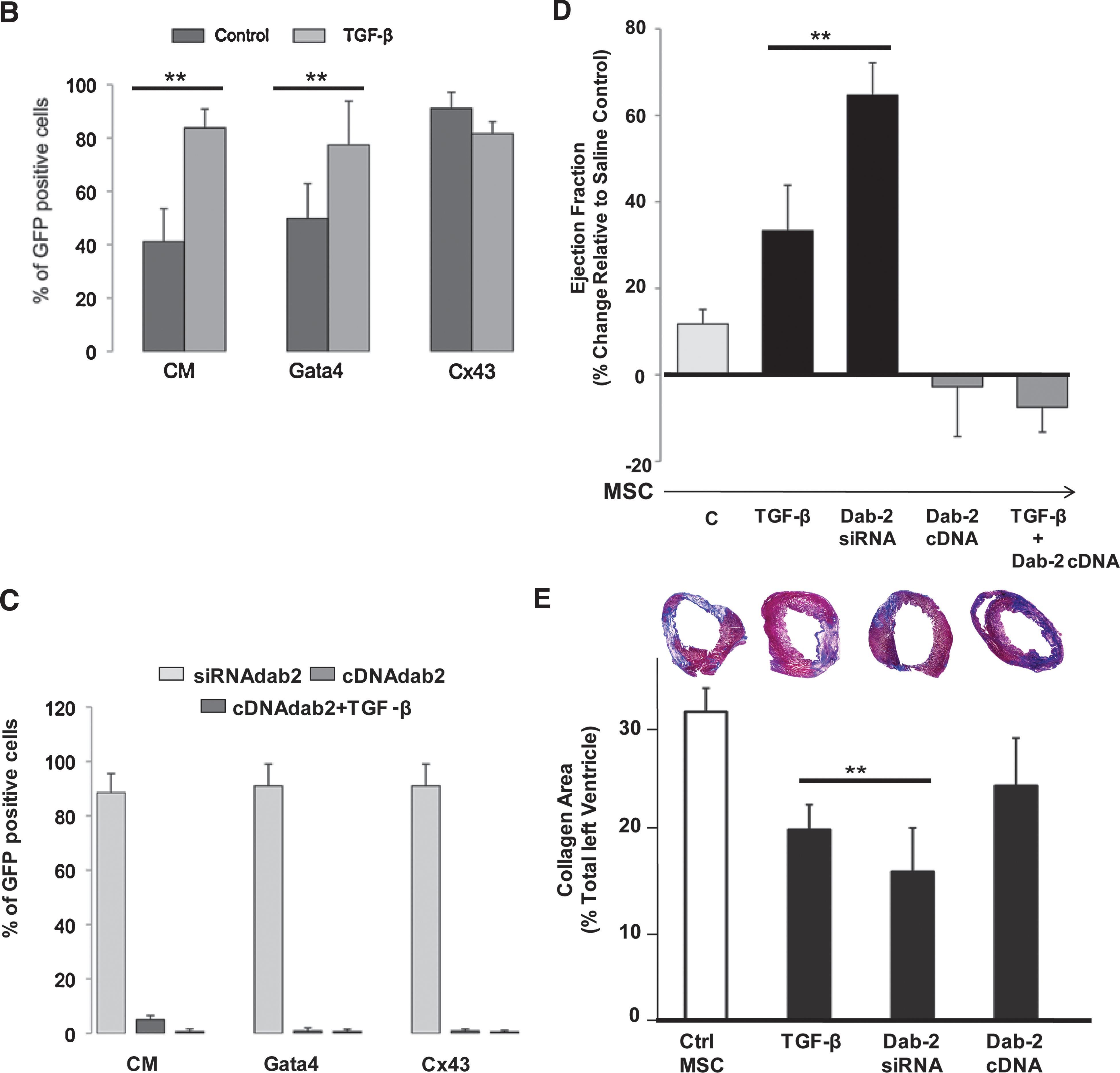
Echocardiographic measurements at 7 and 28 days after acute myocardial infarction in animals that received direction intramyocardial injections of saline, control MSCs, or MSCs with the defined pretreatments. The 7 and 28 day groups are different animals. Data represent mean ± standard deviation.
P < 0.05 compared with saline control.
Dab2, disabled-2; EF, ejection fraction; LVEDD, left ventricular end diastolic dimension; LVESD, left ventricular end systolic dimension; MI, myocardial infarction; MSC, mesenchymal stem cell; TGF-β, transforming growth factor beta.
As an additional measurement of the effect of MSC transplantation and to further determine whether TGF-β or Dab2:siRNA altered MSC survival relative to control MSCs, we performed additional studies injecting 2 × 106 control or pretreated male MSCs into female hearts at the time of LAD ligation. Two weeks after MI the whole left ventricles were harvested and genomic DNA was isolated. We quantified the relative engraftment and survival of control MSCs compared with TGF-β- and Dab2:siRNA-treated MSCs by quantifying the amount of male-specific Sry3 gene by quantitative RT-PCR [37]. Similar to our observations at 7 days after MI using immunohistochemistry, 2 weeks after MI there was no difference in the amount of Sry3 gene in the cardiac genomic DNA isolations between control and TGF-β- or Dab2:siRNA-treated MSCs. On the basis of quantitative PCR, the relative ΔCtSry3:18s for hearts that received TGF-β-treated (n = 4) or Dab2:siRNA-treated (n = 5) MSCs was −4.0% ± 1.3% and 6.1% ± 1.4%, respectively, and that observed in the hearts that received control MSCs (n = 5) was 0.0% ± 1.6% (P = not significant for either pretreated group compared with control MSCs).
Seven days after injection, in hearts injected with control, untreated MSCs, we detected that 40.9% ± 11.1% of the engrafted MSCs expressed cardiac myosin, 49.8% ± 13.1% expressed GATA-4, and 83.6% ± 14.5% expressed connexin 43. Additionally, we found that only 41.2% ± 12.3% of the transplanted cells retained expression of α-MHC and no expression of troponin I, α-sarcomeric actinin, GATA-5, Nkx2.5, or MEF-2 was detected by our methods. This represents a significant reduction in expression of CP compared with the percentage of cells expressing these cardiac markers in cell culture at the time of harvest for cell injection, which ranged between 75% and 95% (Fig. 5B). These data suggest that while MSCs express CPs in culture, the injured myocardial microenvironment does not support cardiogenesis and, in fact, leads to the downregulation of CP expression in transplanted MSCs.
In hearts injected with MSCs pretreated with TGF-β1 (5 ng/mL, 24 h), we observed engraftment and survival of the pretreated MSCs 7 days after MI. However, TGF-β1 pretreatment before transplantation led to the maintenance of CP expression in the engrafted MSCs. We detected α,β-MHC expression in 83% ± 10% and GATA-4 in 80% ± 9% of the TGF-β1 pretreated MSCs as determined by cell counts in immunostained tissue with antibodies against these CPs (Fig. 5A, B). These numbers are similar to those found in cultured cells exposed to TGF-β1 (Figs. 5C and 2), which indicates that MSCs preconditioned with TGF-β1 result in a better inclusion of the transplanted cells into the host myocardium.
To correlate the positive impact of Dab2 loss or gain of function in transplanted MSCs in relation to the cells' ability to sustain CP expression, we counted GFP-positive cells expressing CPs 7 days after MI. We stained heart sections for GFP and α,β-MHC, Gata-4, or Cx-43 from animals that received MSCs transfected with Dab2siRNA or cells overexpressing Dab2 due to transient transfection with Dab2 cDNA with and without TGF-β pretreatment. We found a dramatic downregulation of CP expression in engrafted MSCs that had increased expression of Dab2 (Dab2:cDNA) regardless of whether the cells were pretreated or not with TGF-β, whereas in cells with little or no Dab2 expression (Dab2siRNA), there was CP expression close to 90% of the GFP-positive cells (Fig. 5C).
As stated above, heart function was analyzed by echocardiography at 7 and 28 days postinfarct/MSC transplantation. Seven days after MI, we observed a nonstatistically significant increase in cardiac function in animals that received 2 million control MSCs as compared with saline controls, whereas we observed a significant increase in ejection fraction in animals that received TGF-β1-pretreated MSCs (Fig. 5D). Transplantation of cells with downregulated Dab2 expression by Dab2:siRNA transfection also resulted in a significant increase in cardiac function compared with either control MSCs or TGF-β1-treated cells (Fig. 5D). Conversely, transplantation of MSCs with upregulated Dab2 expression through transfection of Dab2:cDNA resulted in no improvement in cardiac function compared with saline treatment. This negative effect was also observed in animals received Dab2 overexpressing MSCs that were pretreated with TGF-β1 (Fig. 5D).
Importantly, the improved cardiac function observed in animals receiving TGF-β1-treated cells and Dab2:siRNA-transfected cells 7 days after infarction appears to be sustained over time since a significantly enhanced heart function was still measurable 28 days after cell transplantation (Table 1).
To further investigate whether the transplantation of MSCs previously exposed to TGF-β, Dab2-siRNA, or Dab2-cDNA into infarcted hearts influenced remodeling of the left ventricle, we performed Masson's trichrome staining to quantify the extent of LV fibrosis [30]. These analyses demonstrated that the percent LV positive for collagen-based scar was significantly reduced 7 days after cell transplantation of TGF-β and Dab2-siRNA MSCs (P < 0.05 verse control MSCs; Fig. 5E).
Discussion
Stem cell transplantation into infarcted myocardium has been proposed as a feasible therapy to prevent and treat postinfarct cardiac dysfunction. However, the mechanisms and optimization of cardiac remodeling and improvement of cardiac function with cell therapy are still unclear. In general terms, cardiac tissue recovery is often low and the general outcome of cell transplantation is modest [38]. Some of the most challenging aspects of cardiac cell therapy include cell survival, migration, integration, and differentiation. In this series of studies we have defined the molecular signaling associated with adult MSC upregulation of CPs and its consequence on adult stem cell-mediated cardiac recovery.
Our investigations have led us to discover the pivotal role of the adaptor protein Dab2 on the regulation of CP expression in bone marrow-derived MSCs. The information we provide in this report demonstrate for the first time that Dab2 downregulation is central to CP expression in MSCs. This was further confirmed by our findings on Dab2 siRNA-mediated knockdown, which resulted in increased expression of CPs independent of TGF-β1. Our data further demonstrate that Dab2 negatively regulates MSC CP expression; therefore, Dab2 expression needs to be abrogated to induce cardiac specification. Thus, the functional cardiac response of MSCs appears to be inversely correlated with Dab2 expression and directly correlated with CP expression
Dab2 has been described in several studies as a regulator of a variety of cell surface receptor-mediated signaling pathways [39 –41]. Of special interest for studies on cardiogenesis and cardiac specification of stem cells are the well-known interactions of Dab2 and Wnt/β-catenin signaling pathways [40,42 –44]. Wnt/β-catenin appears to be highly relevant for cardiac fate of progenitor cells during development and embryonic stem cells in experimental settings [45,46]. On the other hand, it has recently been demonstrated that Dab2 directly interacts with 2 components of the β-catenin destruction complex, Dvl-3 and axin, and participates in regulation of the Wnt signaling pathway [43,44]. Overexpression of Dab2 was found to lead to a decrease in the Dvl-3/axin association while simultaneously causing an increased level of Dab2 to be found in association with axin, leading to decreased canonical Wnt/β-catenin-mediated signaling [43,44]. It was also observed that overexpression of Dab2 led to increased levels of axin and that loss of Dab2 caused a de-stabilization of axin protein [43,44]. Thus, future studies will need to determine whether downregulation of Dab2 in MSCs leads to upregulation of Wnt/β-catenin-mediated signaling, since increased Wnt/β-catenin-mediated signaling in turn has been associated with increased CP expression in stem cells [47,48].
Dab2 downregulation is usually correlated with increased proliferation and loss of differentiation. Despite the abundant literature on the signaling mediated by adaptor proteins, its role in cardiac cell differentiation and cardiogenesis is unclear [49,50]. Some adaptor proteins and particularly Dab2 have been shown to participate in key control points in stem cell differentiation [43,51 –53]. Our observations suggest that cell type-specific Dab2 response to TGF-β1 may determine the regenerative capacity of different progenitor cell populations.
Our data further the possibilities for cell transplantation for the treatment of cardiac failure. We propose that Dab2 regulates CP expression in vitro in MSCs, and modulating Dab2 levels may be a strategy for inducing CP expression in vivo after transplantation. Also, cells pretreated to maintain high expression of cardiogenic proteins led to significant greater improvement in cardiac function compared with control MSCs. Importantly, the functional consequences of pretreatment of MSCs with TGF-β or Dab2:siRNA occurred in the absence of alterations in MSC engraftment or survival as measured by 2 distinct and independent techniques. Whether this benefit is associated with the integration of CP-expressing cells in the infarct border zone leading to improved integrity of the infarct border zone leading to improved LV remodeling, as we have seen in other studies [30], or is due to an as yet unknown modulation of paracrine factor expression by the MSCs needs to be determined [54].
Footnotes
Acknowledgments
This work was funded by the State of Ohio and the Skirball Foundation. M.E. Mayorga is the recipient of an AHA postdoctoral fellowship form the Great Rivers affiliate.
Author Disclosure Statement
No competing financial interests exist.