
Abstract
The maintenance of genomic integrity is crucial to embryonic stem cells (ESC) considering the potential for propagating undesirable mutations to the resulting somatic and germ cell lineages. Indeed, mouse ESC (mESC) exhibit a significantly lower mutation frequency compared to differentiated cells. This could be due to more effective elimination of genetically damaged cells via apoptosis, or especially robust, sequence-conserving DNA damage repair mechanisms such as homologous recombination (HR). We used fluorescence microscopy and 3-dimensional image analysis to compare mESC and differentiated cells, with regard to HR-mediated repair of spontaneous and X-ray-induced double-strand breaks (DSBs). Microscopic analysis of repair foci, flow cytometry, and functional assays of the major DSB repair pathways indicate that HR is greater in mESC compared to fibroblasts. Strikingly, HR appears to be the predominant pathway choice to repair induced or spontaneous DNA damage throughout the ESC cycle in contrast to fibroblasts, where it is restricted to replicated chromatin. This suggests that alternative templates, such as homologous chromosomes, are more frequently used to repair DSB in ESC. Relatively frequent HR utilizing homolog chromosome sequences preserves genome integrity in ESC and has distinctive and important genetic consequences to subsequent somatic and germ cell lineages.
Introduction
M
As indicated above, there are different types of repair acting on DSBs, including NHEJ and HR [13]. Choice of repair pathway(s) is influenced by at least 2 factors: the source of damage and the cell cycle phase at the time of damage. In differentiated cells, the choice of pathway is constrained by the cell cycle stage; NHEJ may be active during the entire cell cycle but HR thought to be only active in S and G2. The choice of repair pathway is also influenced by the source of damage: DSBs associated with DNA replication are primarily repaired by HR, whereas ionizing radiation-induced DSBs are repaired by NHEJ [14 –17]. In mammalian cells, differences in accumulation of the DSB indicator, phosphorylated histone 2AX (γH2AX), in single versus double NHEJ and HR mutants were interpreted as cooperation of the 2 DSB repair pathways [18]. In fact, the absence or overexpression of proteins of the NHEJ pathway enhances or suppresses HR, respectively [19,20]. Therefore, the availability of pathway-specific repair proteins is also a factor that influences the choice of repair pathways and is likely to be correlated with cell differentiation and proliferation.
Proteins involved in the recombinational repair of DSBs were first characterized in Saccharomyces cerevisiae and are encoded by the RAD52 epistasis group of genes [21,22]. RAD51 is the primary eukaryotic recombinase responsible for initiating DNA strand exchange during HR, a function that is conserved from bacteria (RecA) to humans [23]. Upon treatment of mammalian cells with agents that produce DSBs, several proteins that participate to the DNA damage response undergo redistribution and concentration within the cell nucleus to form discrete foci that can be observed by immunofluorescence [24]. This is especially well documented for the DSB marker γH2AX and the recombinase protein RAD51 [24 –29]. However, even in the absence of external damaging agents cells also contain γH2AX foci, which are believed to result from replication fork collapse [30 –32]. It is known that RAD51 associates with chromatin as cells progress through S-phase, interacting with components of the DNA replication apparatus. There is consensus that RAD51 association with chromatin is essential for the repair of DSBs that originate during replication [31]. In vivo analysis of the recruitment of DNA repair proteins at the sites of laser-induced DNA lesions shows a transient assembly of NHEJ repair factors, which precedes a prolonged occupancy by HR factors, such as RAD51 [33]. These and other authors conclude that NHEJ and HR are not competing pathways, NHEJ being an immediate early repair pathway that precedes a more prolonged attempt to repair persistent DNA lesions by HR [33,34]. Thus, in both external and replication-induced DSBs, the cytologically visible stabilization of damaged-induced ssDNA by RAD51 polymerization is considered a marker of HR activity, even if the strand invasion process and the completion of DNA repair do not occur until a homologous template is available.
In this study we aimed to identify the contribution of HR to the repair of DSBs in mESC. We demonstrate that additional mechanisms to preserve mESC genome integrity are related to preferential utilization of error-free DNA damage repair pathways, specifically HR, throughout the cell cycle. Our results show that mESC utilize a unique DNA repair strategy, thereby preserving genome integrity compared to other mammalian cells.
Materials and Methods
Cell lines and X-irradiation
Primary fibroblasts were isolated from mouse ears as described by Shao et al. [5]. Mice were sacrificed at 2–4 months of age. Cells were initially plated at 106 in 100 mm plates (10 mL medium) and allowed to achieve 80% confluence (about 2–3 days after plating). Then, cells were split 1:3 and plated for immunocytochemistry analysis as described below.
mESC were isolated as described by Cervantes et al. [3]. They were grown in the presence of leukemia inhibitory factor and bone morphogenetic protein 4 as a substitute for feeder cells to maintain full pluripotency as described by Ying et al. [35]. Cells were not allowed to exceed 50% confluence. mESC lines were monitored for pluripotency by measuring expression of Oct4 and Nanog [36] (Supplementary Fig. S1; Supplementary Data available online at
Radiation was delivered at the rate of 1 Gy/min using an aluminum filter at 100 kVp (Faxitron Cabinet X-ray System; Faxitron X-ray Corp.).
Mitotic synchronization
mESC were synchronized in M-phase by incubation for 16 h with 200 ng/mL nocodazole (Sigma). The mitotic inhibitor was removed by repeated washes with regular ESC medium, and cultures returned to the incubator. Samples were taken at different time points. Cell synchronization was verified by flow cytometry using propidium iodide staining (Sigma) as described elsewhere [37].
Cell cycle profile assays
Cells were plated on poly-
Immunofluorescence
Cells were fixed in 1% paraformaldehyde for 10 min at room temperature. Membrane permeabilization was achieved by incubation in buffer A (0.1% sodium azide PBS, 0.5% Triton-X, 0.5%–1% BSA) for 10 min. EdU incorporation into chromatin was observed by chemical incorporation of a fluorescence dye, in this case Alexa Fluor 488, into the DNA containing modified nucleotides following manufacturer's specifications (Click-iT EdU imaging kit; Invitrogen). Primary and secondary antibodies were diluted in 0.1% sodium azide PBS, 0.2% Triton-X, and 0.5%–1% BSA. We used rabbit polyclonal anti-RAD51 (Santa Cruz Biotechnology), mouse monoclonal anti- γH2AX (Upstate), mouse anti-OCT4 antibody (Santa Cruz Biotechnology), and rabbit anti-NANOG polyclonal antibody (Cosmo Bio). As secondary antibodies, we used anti-mouse Alexa Flour 594 (red) and anti-rabbit Alexa Flour 488 (green) from Molecular Probes. Cells were counterstained with DAPI (Sigma) and slides mounted on Vectashield (Vector Laboratories).
Image acquisition and analysis
Labeled mouse fibroblast nuclei were imaged using a conventional epifluorescence microscope (Zeiss 200M) or a Zeiss LSM510 Meta Confocal Laser Scanning Microscope. Confocal image stacks (0.15 μm per slice) were acquired using a C-Apochromat water-immersion 63 × lens (numerical aperture [N.A.] = 1.2) and 3 × digital zoom (voxel size: 0.093 × 0.093 × 0.15 μm, X, Y, Z, respectively). Acquired image stacks were processed by Huygens deconvolution software (Scientific Volume Imaging) using the quick-MLE (Maximum Likelihood Estimation) function for confocal images. Three-dimensional reconstructions, computer-generated visualization of the signal, and image analysis were obtained using Imaris software (Bitplane, A.G.), after defining intensity thresholds set at 2 standard deviations above background for each of the involved channels. The number of foci was calculated using Spot Finder software by Imaris (Bitplane, A.G) after intensity segmentation of the images and size discrimination (objects >0.2 μm diameter). The percentage of colocalization between 2 color channels is the percentage of voxels (with intensities above background) that are superimposed for both channels.
Real-time polymerase chain reaction
Total RNA from cultured cells was purified using an RNeasy kit (Qiagen) and reverse transcribed using the TaqMan Reverse Transcription Reagent kit (Applied Biosystems). Expression of the RAD51 gene was measured with a quantitative real-time polymerase chain reaction (PCR) assay as previously reported [38]. Expression level was calculated as a ratio of the mRNA level for a given gene relative to the mRNA level for glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primer sequences are as follows: mRad51-F, 5′-TGA TGA GTT TGG TGT CGC AGT G-3′ and mRad51-R, 5′-CGA ACA TGG CTG CTC CAT CTA C-3′; mBRCA1-2F, CGC CTC ACT TTA ACT GAC GCA A and mBRCA1-2R, AGA CCG GAC CAC CCA TGA ATA G; mligase4-F, GTC AAG CCC GAG TGG CTT TTA and mligase4-R, CGG CAC GCA TGT TTT TGT CT; mXRCC4-F, TGG AAT GAT GTT CAA GGC CG and mXRCC4-R, TCC AAG GCT TCT TTG GCA CTC; mGAPDH-F, 5′-CTC CAC TCA CGG CAA ATT CAA-3′ and mGAPDH-R, 5′-GAT GAC AAG CTT CCC ATT CTC G-3′. For each RNA sample, the real-time PCR was performed in triplicate.
Immunoblotting
Cells were harvested and washed with ice-cold PBS. Nuclear protein extracts were prepared as described [38]. Equivalent concentrations of nuclear extracts were electrophoresed with sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels and transferred onto polyvinylidene difluoride membranes. Membranes were blocked at room temperature for 30 min in blocking buffer [3% nonfat dry milk in PBS containing 0.1% Tween 20 (T-PBS)] and incubated at 4°C overnight with primary antibody diluted in blocking buffer. After 5 washes with T-PBS, membranes were incubated at room temperature for 1 h with secondary antibody diluted in blocking buffer. Immunoblots were observed by enhanced chemiluminescence. Primary antibodies used in this study were rabbit-anti-RAD51 and Ligase 4 (Santa Cruz Biotechnology), and rabbit anti-tubulin (Sigma).
Cell-free HR assay
HR catalyzed by cell-free extracts was performed essentially as described previously [39], using a plasmid substrate pair: SalI linearized pSV2neoDL and single-stranded circular ssneoDR, which were kindly provided by Dr. Colin Campbell, University of Minnesota. Briefly, 500 ng of each of these substrates were incubated with nuclear protein extracts prepared from ear fibroblasts or ESC at 37°C for 2 h. Purified plasmid DNA was used to electroporate the recombination-defective Escherichia coli strain DH10B using a Bio-rad Gene Pulser Xcell™. Kanamycin-resistant colonies (resulting from the generation of an intact neomycin phosphotransferase gene via HR) and ampicillin-resistant colonies were enumerated. The ratio of kanamycin-resistant to ampicillin-resistant colonies was used to calculate the frequency of HR-induced recombinants in the individual extracts.
In vitro assay for NHEJ
The NHEJ in vitro assay was conducted as described by Liang et al. [40,41]. Briefly, the linearized DNA substrate was incubated with nuclear protein extract in 50 mM Tris-HCL (pH 7.6), 10 mM MgCl2, 1 mM dithiothreitol, 1 mM ATP, and 25% polyethylene glycol 8000 at 14°C for 2 h. Aftr the reaction, DNA was deproteinized and purified by using a PCR purification kit (Qiagen). The amount of end-joined products was measured by real-time PCR using 2 pairs of primers: a loading control primer pair, which amplifies a DNA fragment 68 bp away from the end-joining junction, and a second primer pair flanking the joining junction, selectively amplifying end-joined products. The NHEJ activity was calculated as the ratio of the amount of end-joined products to total amount of DNA substrate. Real-time PCR was carried out on a GeneAmp 7900HT Sequence Detection System using SYBR Green dye for signal detection. The end-joining experiment was performed in triplicate.
Results
HR repair proteins bind to sites of DNA damage throughout the cell cycle in mESC
We performed a quantitative analysis of the spatial and cell cycle distribution of γH2AX and RAD51 foci in exponentially growing mESC and primary mouse fibroblasts. Sites of active DNA replication during S-phase progression were identified by the incorporation of EdU, which can be observed by postfixation chemical incorporation of a fluorescence dye, in this case Alexa Fluor 488, to the modified nucleotide. In this way, replication-associated DNA damage and HR protein activity, evident through γH2AX and RAD51 foci observation during S-phase (EdU-positive cells), respectively, can be identified and quantified simultaneously (Figs. 1A and 2C). Consistent with previous data [42], our results show that mESC have a cell cycle profile differing from that of somatic cells. As shown in Fig. 1B, 60% of mESC are EdU positive compared with 20% of fibroblasts. These results are consistent with the mESC cycle profile obtained by FACS analysis (Figs. 2B and 6A). Therefore, in unsynchronized, exponentially growing mESC, there is a large fraction that is in S-phase (∼60%–70%). For mESC, 93% show γH2AX, as compared to 45% of fibroblasts (Fig. 1B). Previous studies in mESC and in mouse embryonic teratocarcinoma cells (mECC) also showed a high proportion of noninduced single and DSBs [43]. Both mESC and mECC are very rapidly dividing cells with similar cell cycle profiles and cell cycle duration, estimated at 10–12 h. It has been suggested that the high levels of DNA damage observed in mECC may be due to their rapid replication rate, perhaps as a consequence of incomplete maturation of replication forks [44]. Our data are consistent with these reports. However, the high replication rate does not seem to affect their capacity to detect and repair such damage, as indicated by the presence of RAD51 foci in γH2AX foci-positive cells (Fig. 1A). Indeed, in the absence of external damage, 82% of mESC exhibit RAD51 foci as compared with 20% of adult fibroblasts (Fig. 1B).
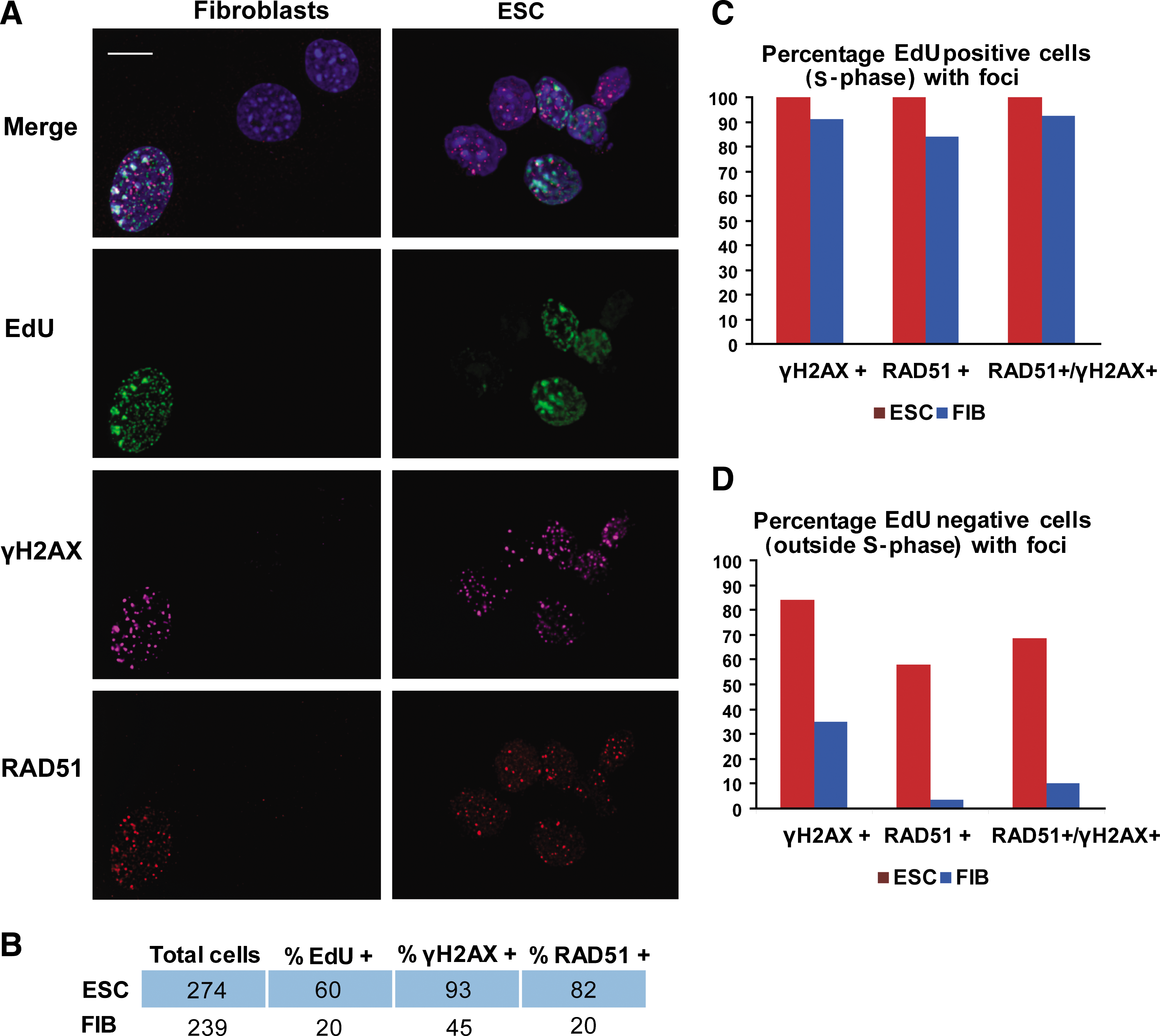
RAD51 proteins bind to double-strand breaks (DSBs) outside of S-phase. Mouse embryonic stem cells (mESCs) and primary fibroblasts (FIB) in log phase were pulse-labeled for 10 min with 5-ethynyl-2′-deoxyuridine (EdU), stained with anti-γH2AX and anti-RAD51 antibodies as indicated
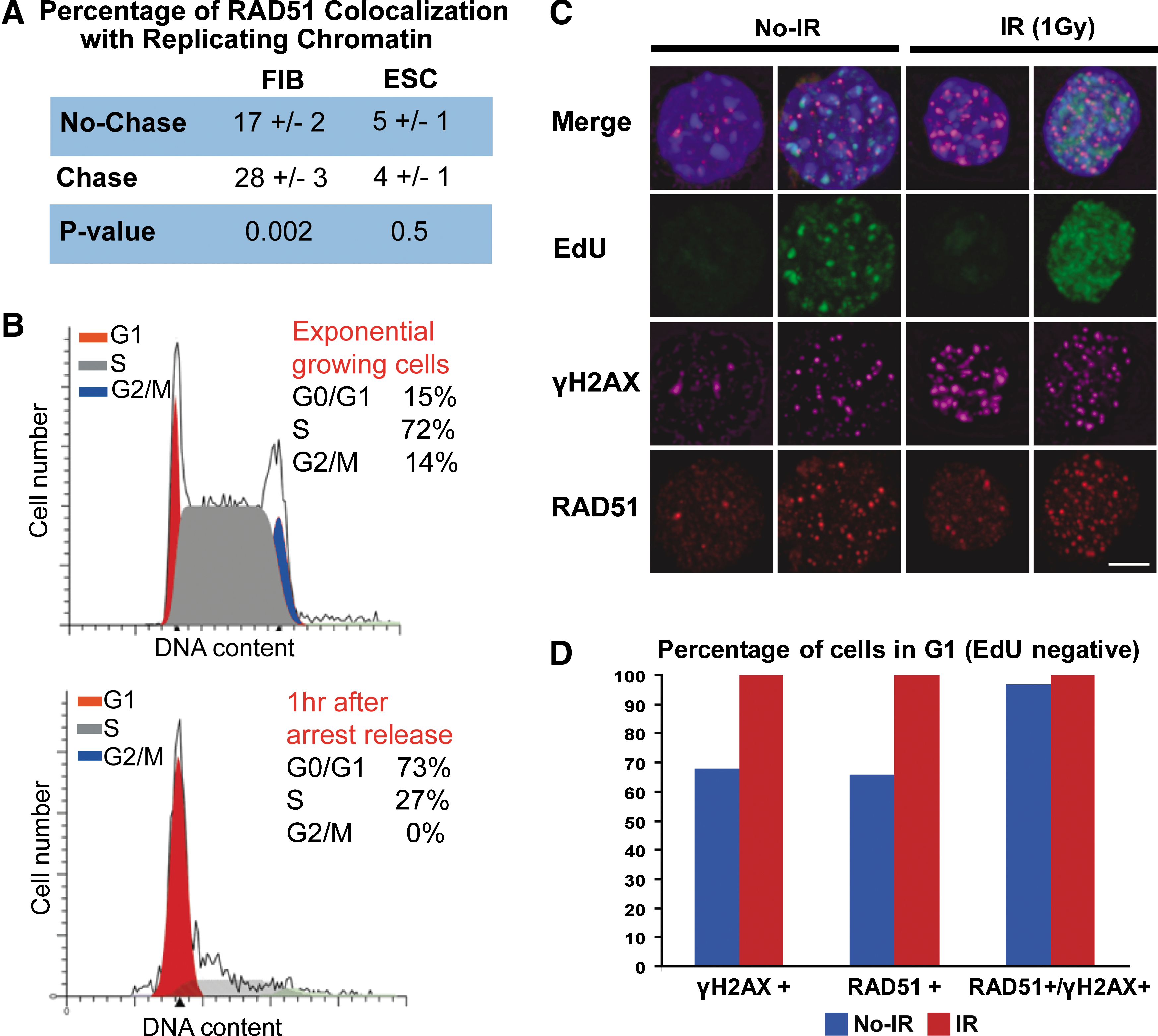
RAD51 foci are not associated with replicating chromatin in mESC.
In both mESC and fibroblasts, detection of RAD51 foci is highly correlated with the number of cells that are in S-phase (EdU-positive cells): 84% of fibroblasts and 100% of mESC (Fig. 1C). In adult primary fibroblasts that are not in S-phase, as evidenced by lack of EdU incorporation, only a small fraction of cells show RAD51 foci (4%), which represents 10% of the γH2AX foci-positive cell population (Fig. 1D). These results suggest that HR in fibroblasts is initiated mostly during DNA replication (S-phase). In contrast to fibroblasts, 58% of mESC show RAD51 foci in the absence of EdU staining, representing 88% of γH2AX foci-positive cells. This indicates that HR activity occurs outside of S-phase in mESC (Fig. 1D).
RAD51 foci in mESC are assembled independently of DNA replication
We used confocal microscopy and 3-dimensional image processing for quantitative analysis of the subnuclear organization of RAD51 foci and their association with newly replicated chromatin (Fig. 2). Specifically, we investigated whether RAD51 foci were preferentially associated with postreplicative chromatin in mESC. mESC and primary fibroblasts were pulse-labeled with BrdU for 10 min and the association of RAD51 foci with newly replicated chromatin was measured before and after a chase period of 30 min. In primary adult fibroblasts, association of RAD51 foci with postreplicative chromatin increased significantly (P value =0.002, Fig. 2A), which it is consistent with previous reports [45]. Conversely, no increased association is seen in mESC (P value = 0.5) (Fig. 2A). Therefore, RAD51 foci in mESC can be assembled at DSB sites not originated during DNA replication. This suggests that in mESC, RAD51 protein is not only recruited at DSBs that originate from collapse of replication forks, but is also recruited at DSBs that originate from other sources.
Interestingly, bivariate fit analysis of the percentage of colocalization of γH2AX and RAD51 foci in mESC showed colocalization decrease with increasing nuclear volume (Supplementary Fig. S2). Thus, in contrast to differentiated cells, the percentage of colocalization decreases as mESC progress from G1- to M-phase. This result is consistent with the observed RAD51 recruitment at DSBs, independently of DNA replication (Fig. 2).
We consequently questioned whether HR was active during G1. mESC were partially synchronized in M-phase by incubation with the mitotic inhibitor nocodazole. One hour after nocodazole was removed from the cell culture, flow cytometry analysis revealed that 73% of mESC were in G0/G1 and 27% in S-phase (Fig. 2B). The cell cycle progression after mitotic arrest release is shown in Supplementary Fig. S3. Double staining with RAD51 and γH2AX antibodies, coupled with S-phase cell discrimination by EdU incorporation, demonstrated HR protein activity during G1 (Fig. 2C). RAD51 foci were present in 68% and 100% of G1 cells (EdU negative) before or after X-irradiation, respectively (92 of 135 cells before and 200 of 200 after irradiation). More than 90% of the scored cells showed DNA damage (γH2AX-positive cells) in the presence and absence of X-irradiation (Fig. 2D).
RAD51 foci in mESC are assembled independently of the source of damage
We investigated whether this apparent predominance of HR in mESC was maintained after X-irradiation, an agent known to cause DSBs, by analyzing γH2AX and RAD51 foci formation (Fig. 3 and Supplementary Fig. S4). Three-dimensional reconstruction and measurements of individual nuclei showed that in both mESC and adult primary fibroblasts the number of γH2AX and RAD51 foci per nucleus was initially increased after 1 Gy X-ray exposure (P value <0.001 in all cases). Compared to levels in unirradiated cells, the increase in number of foci as a response to X-irradiation was greater in murine primary fibroblasts than in mESC. However, the temporal dynamic of repair, as measured by the time required for a reduction in foci to nonirradiated levels, was similar in the 2 cell types (from 2 to 6 h after irradiation, Fig. 3B and Supplemental Fig. S4). Previous reports have shown a linear correlation between γH2AX foci formation and DNA damage [25]. Thus, the elevated number of γH2AX foci in fibroblasts suggests that they may sustain more DNA damage than mESC from the same level of X-irradiation.
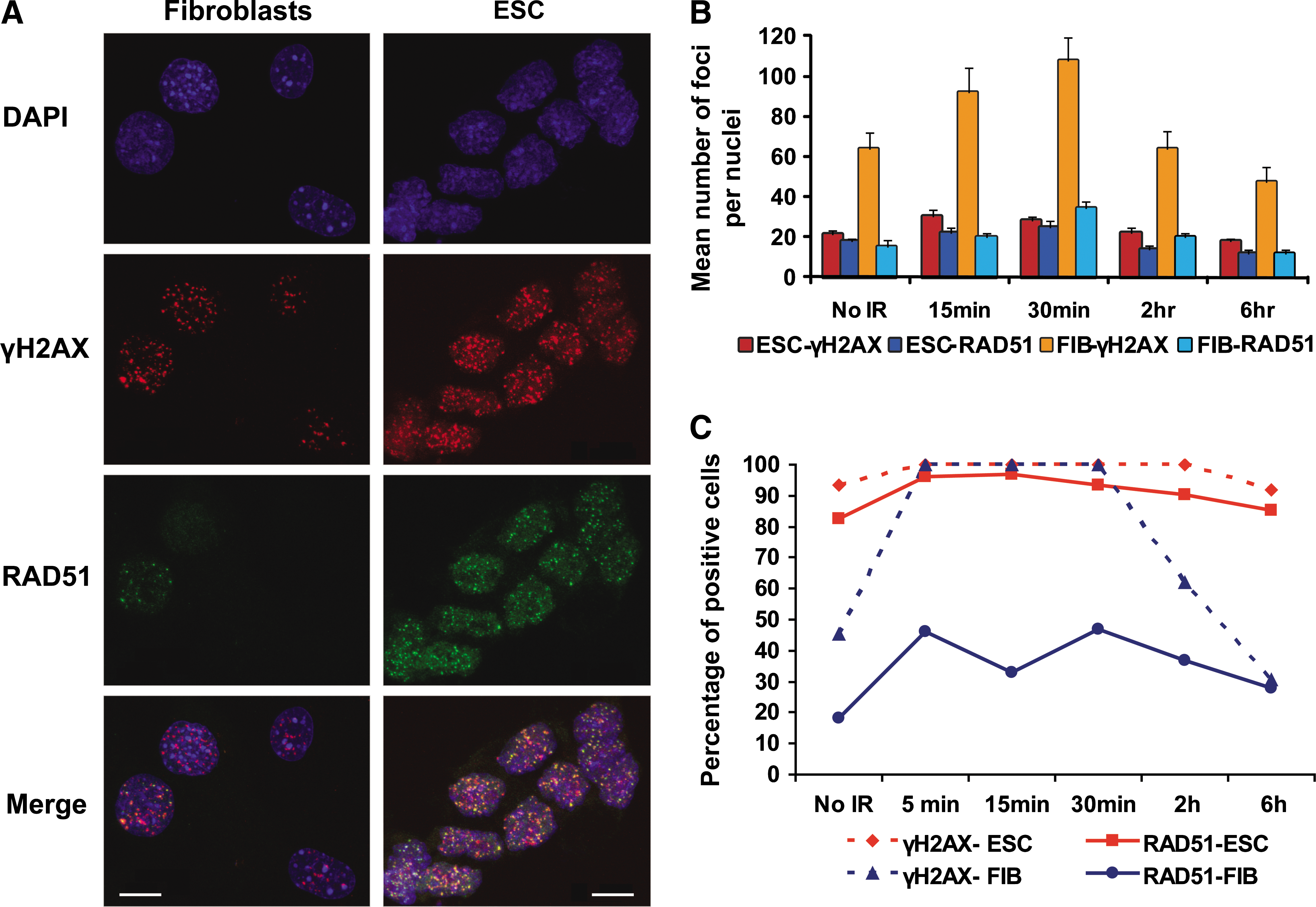
RAD51 binds X-ray-induced DSBs in mESC.
However, the most remarkable difference between the 2 cell types is that >95% of mESC that showed damage, as measured by the presence of γH2AX foci, also had RAD51 foci (Fig. 3A and C). The high correlation between γH2AX- and RAD51-foci-positive cells was maintained before X-ray exposure and at all subsequent time points. For fibroblasts, in contrast, the correlation between γH2AX- and RAD51-positive cells was dramatically lower after irradiation (Fig. 3C), suggesting that HR is predominantly repairing DSBs generated during replication, but not DSBs induced by irradiation.
We further analyzed the level of HR in the repair of X-ray-induced damage by analysis of colocalization between γH2AX and RAD51 foci in individual nuclei (Fig. 4). Colocalization of γH2AX and RAD51 foci was greater in mESC compared with adult fibroblasts in the absence of irradiation (P value <0.001). This difference in colocalization between cell types is remarkably enhanced (to as much as 20-fold) after X-irradiation (Fig. 4B). This disparity is mostly due to the fact that in fibroblasts the formation of large numbers of new γH2AX foci after irradiation was not accompanied by a proportional increase of RAD51 foci (Fig. 3). However, this observation cannot be explained by assuming differing degrees of damage inflicted to each cell type. While the number of foci increases, the fraction of colocalizing RAD51 and γH2AX foci remains greater (>20-fold) with higher doses of X-rays in ESC as compared with fibroblasts (Fig. 4C). Interestingly, there is a statistically significant reduction (0.42-fold, P value = 0.019) of the percentage of colocalization of γH2AX and RAD51 foci in 5 Gy versus 1 Gy X-irradiated fibroblasts. Although not significant, we observe the opposite tendency in ESC (0.64-fold increase). Overall, these data suggest that the HR repair in fibroblasts, compared to mESC, is saturated at lower levels of induced DNA damage.
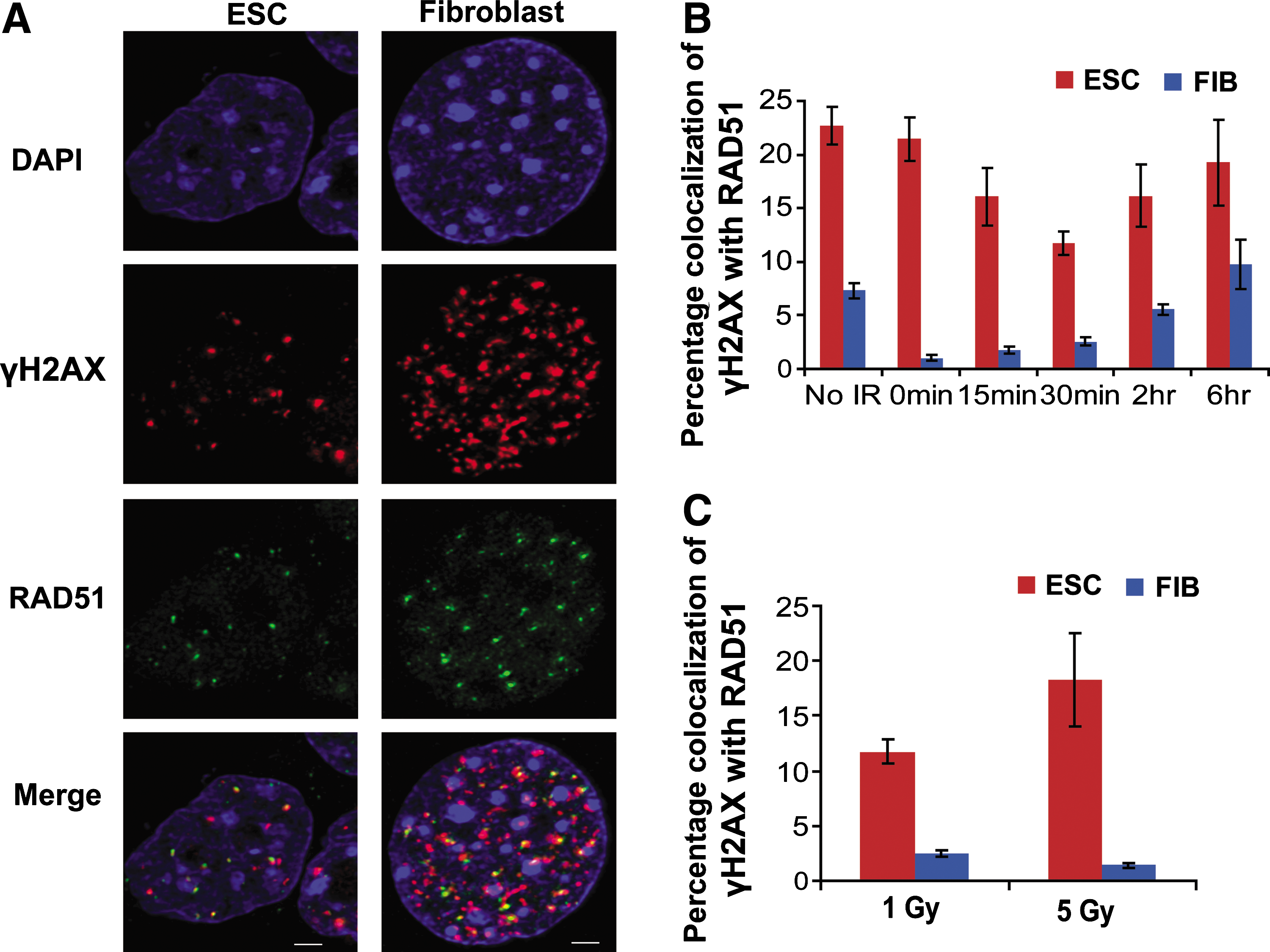
Increased recruitment of RAD51 protein at DSBs in mESC as compared with primary fibroblasts. mESCs and primary fibroblasts (FIB) in log phase were stained with antibodies against γH2AX and RAD51 as indicated
Most importantly, our results show that HR is active during the entire cell cycle, including G1, in mESC.
Differential activity of HR and NHEJ repair pathways in mESC and fibroblasts
Different levels of expression and/or activity of repair proteins in mESC and fibroblasts could alter relative contributions of different repair pathways. To examine this possibility, we analyzed the mRNA and protein levels of major players of HR and NHEJ pathways (Fig. 5A and B). We found that both RAD51 mRNA and protein levels were elevated in mESC compared with fibroblasts (Fig. 5). The mRNA level of RAD51-associated proteins BRCA1 and BRCA2 was also upregulated in mESC. Both proteins are well documented to be involved in the initiation of the recombination-mediated DNA damage repair process [46]. Conversely, both members of the XRCC4-ligase IV complex involved in NHEJ [47] were down-expressed in mESC.
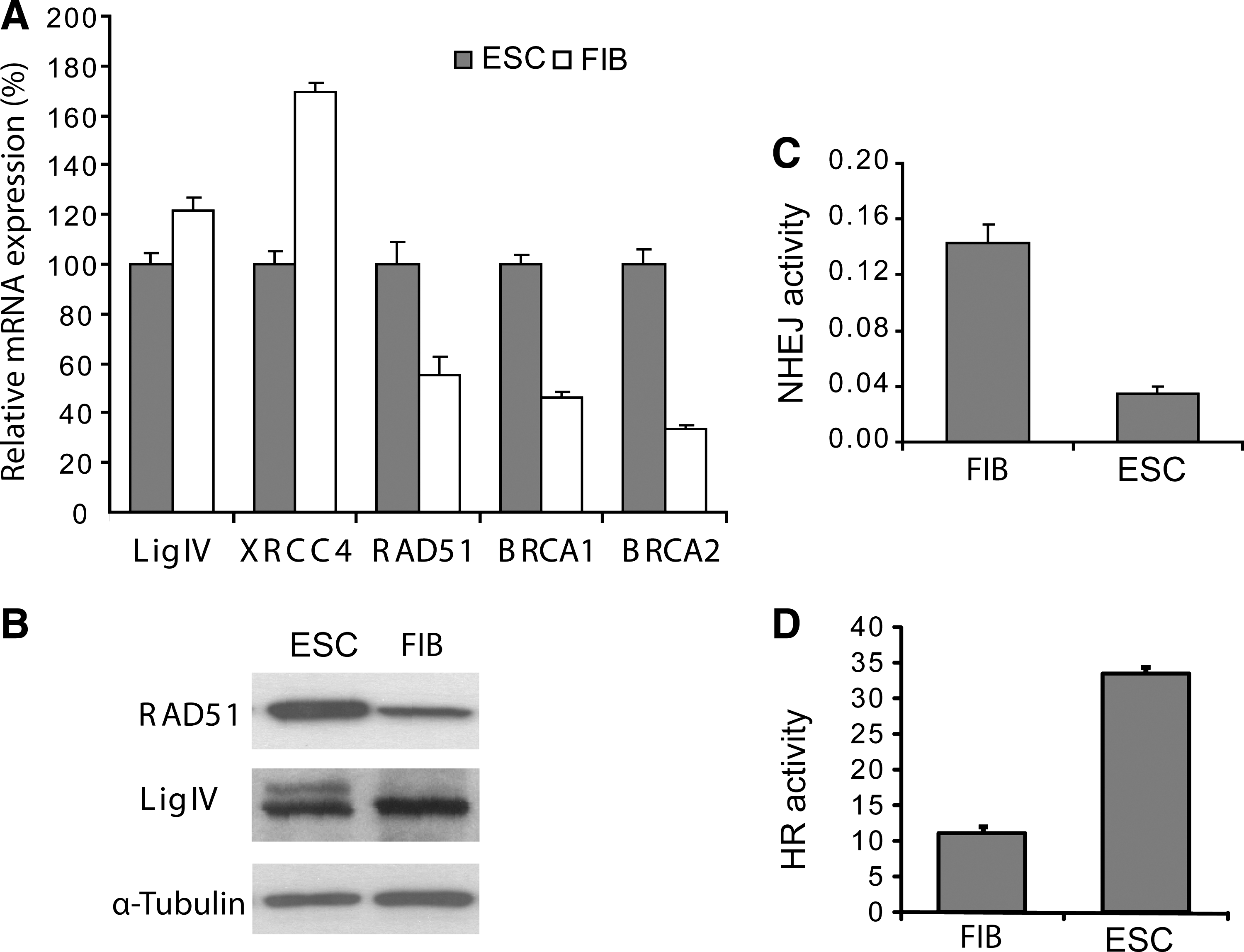
mESC express greater homologous recombination (HR) but less nonhomologous end-joining (NHEJ) activity than fibroblasts.
Further, we confirmed the differential activity of HR and NHEJ pathways in mESC and fibroblasts by in vitro functional assays [39 –41]. Recombination reactions using nuclear protein extracts prepared from exponentially growing cells showed a 3-fold increase in HR activity in mESC compared to primary fibroblasts (P value <0.001; Fig. 5C). Further, in vitro end-joining experiments showed a 5-fold decrease in the NHEJ activity in mESC (P value <0.001; Fig. 5D).
The dynamics of RAD51 levels through the cell cycle were analyzed by flow cytometry (FACS). BrdU-labeled exponentially growing cells were incubated with a marker for DNA content (7AAD) and with antibodies specific for RAD51. First, we used the 7AAD and BrdU signals to generate a horseshoe-shaped distribution to accurately distinguish cells in the different stages of the cell cycle: G1, S, and G2/M (Fig. 6A). Then, we determined the percentage of cells that showed RAD51 levels above background and the mean levels of RAD51 in these 3 cell populations (Fig. 6B and C). Consistent with our imaging studies, 100% of mESC show RAD51 protein levels above background at all phases of the cell cycle (Fig. 6B). In contrast, in fibroblasts RAD51 protein was almost absent in G1, peaked in S-phase, and decreased somewhat in G2 (Fig. 6B), which is in agreement with previous reports [17]. The overall RAD51 protein level, measured by the mean fluorescence intensity in RAD51-positive cells (Fig. 6C), was higher in mESC as compared with fibroblasts in all phases of the cell cycle, correlating with our western blot analysis (Fig. 5). In mESC, the levels of RAD51 are the same for cells in G1- or S-phase and slightly elevated during G2/M (Fig. 6C). These data strongly support the immunofluorescence imaging data described above, showing RAD51 activity throughout the mESC cycle, including G1.
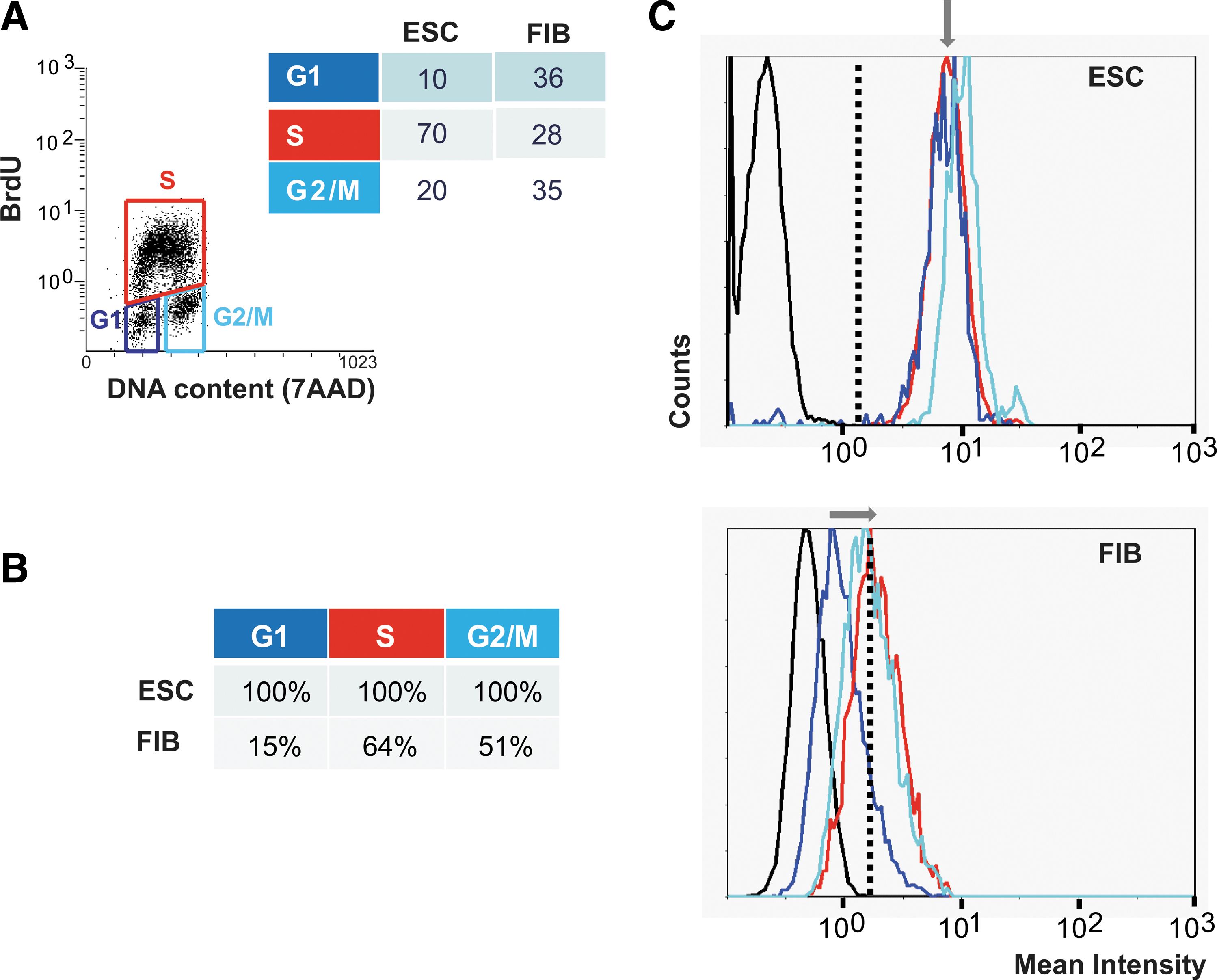
Sustained expression of RAD51 protein throughout the cell cycle in mESC. Exponentially growing mESCs and fibroblasts (FIB) were pulsed with BrdU, incubated with RAD51 antibody and 7-amino-actinomycin D (7AAD) (DNA content marker), and analyzed by FACS to quantify the levels of RAD51 protein at each stage of the cell cycle.
Discussion
Previous reports showed that there is a constant activity of HR during replication thought to be, in part, protecting fragile DNA sites and thus preventing chromosomal rearrangements [48]. Indeed, we demonstrated that replication-associated damage and HR protein activity, evident through γH2AX and RAD51 foci colocalization, occurs in a high percentage of replicating cells. We found that in primary fibroblasts, with intact cell cycle checkpoints, HR is restricted to S-phase. In these cells, the coordination of DNA damage-sensing proteins and mediators of DNA damage check points would promote cell cycle arrest and allow the completion of HR before the cell enters G2 [49]. After externally induced DNA damage, and in common with teratocarcinoma cells, human and murine ESC traverse G1-phase progresses through S-phase and accumulate in at the G2/M boundary [4,37,44,50]. In principle, a lack of damage-induced checkpoint at the G1/S boundary would limit the time and resources available for DNA damage repair and compromise genomic integrity. However, mammalian ESC exhibit the robust induction of a DNA damage response that facilitates the elimination of damaged cells by apoptosis [4,37,44,50]. Consistent with previous reports [43], our results demonstrate that DNA damage response mechanisms in mESC include the recruitment of DNA repair components and the activation of γH2AX foci. Moreover, our results indicate that an additional mechanism for the maintenance of mESC genomic integrity relies on the increased activation of HR throughout the cell cycle, in marked contrast to fibroblasts.
Previous studies have reported an enhancement of HR in mESC compared to a variety of somatic cell types, as measured by gene targeting of an inserted construct [10] or by the analysis of spontaneous chromosomal aberration frequency in various repair-gene knockout mice [12]. Indeed, the early embryonic mortality of mouse strains carrying mutations in HR-related genes suggests that HR has an important role during early development [51]. Our in vitro assays showed 3-fold increased HR activity in mESC compared to primary fibroblasts, which correlates with the mRNA and protein levels of HR-related proteins in these cell types. Further, RAD51 recruitment to DSBs occurs in both the absence and presence of X-ray-induced damage in a greatly elevated proportion of mESC compared with fibroblasts.
The recruitment of RAD51 at DSBs indicates that repair pathway choice toward HR is made even if HR is not completed or the damaged cells do not make it through the cell cycle [14 –16,33]. The formation of nuclear RAD51 foci in response to DNA damage is due to the redistribution and concentration of RAD51 protein at DSBs and not due to de novo protein synthesis [52]. However, RAD51 overexpression reduces the number of DSBs and chromatid-type aberrations after DNA damage [52]. Thus, the lower level of RAD51 protein in fibroblasts could explain why these cells sustain more DNA damage than mESC with the same level of X-irradiation, and why the HR repair response in fibroblasts, compared to mESC, is saturated at lower levels of induced DNA damage. On the contrary, the elevated levels and sustained expression of RAD51 protein throughout the cell cycle in mESC is consistent with the reported greater resistance of both human and murine ESC to oxidative stress, as compared with differentiated derivatives [53,54]. Continuous activity of the mechanisms for the detection and repair of DNA defects throughout the cell cycle may explain why mESC are apparently more resistant to oxidative stress and external X-ray-induced damage. Nevertheless, while the higher amount of RAD51 protein in mESC would likely affect the recruitment of RAD51 to DSBs and therefore RAD51/γH2AX colocalization, it is likely that other factors also contribute to the elevated and sustained DNA repair in these cells.
The state of cell proliferation could be the driving force for choice of DNA repair pathway, because the availability of a sister chromatid for HR is limited to S-phase. During mammalian nervous system development the 2 major pathways of DSB repair act in a spatiotemporal nonoverlapping manner showing clear compartmentalization; HR is more prominent in proliferating neural precursor cells, whereas NHEJ predominates at later developmental stages [11]. In yeast, where constant proliferation is the life strategy, HR is the predominant pathway of DNA repair [2].
Our in vitro functional assays show that HR activity is elevated in mESC compared with fibroblasts, which may be in part explained by differences in cell cycle profiles between both cell types: mESC divide faster and consequently, in exponentially growing mESC, there is a large fraction of cells that are in S-phase. However, in somatic cells NHEJ is an equal pathway of DSB repair, including during S-phase [17,55]. Moreover, this would not explain why RAD51 protein is recruited at DSB sites throughout the mESC cycle, including G1. This is dramatically different from what we observe in primary fibroblasts, and from what has been reported in other adult mammalian cell types, where HR is essentially confined to S-phase [14].
Most interesting, our results show that, as in yeast, HR in mESC can be independent of DNA replication. Therefore, in mESC RAD51 nucleoprotein filament formation and the search for a homologous DNA sequence to repair DSBs can be performed in the absence of a spatially proximate sister chromatid for use as a template. Johnson and Jasin used a recombination reporter to show that a sister chromatid is the most frequent template for HR in differentiated mammalian cells [56]. The choice of a donor DNA sequence for use as a repair template could be determined by spatial proximity and sister chromatids would be favored by such a positional effect. However, HR mainly between sister chromatids would predict preferential association of HR proteins with newly postreplicative chromatin, which is not the case in mESC.
HR in the absence of a sister chromatid would necessitate the use of the DNA sequence on the homologous chromosome as a template for recombinational repair, possibly resulting in a crossover that can potentially lead to LOH. Indeed, in mESC, 27% of somatic mutation events are due to MR [3,4]. However, mESC LOH as a consequence of MR is reported to be 2 orders of magnitude lower than that of adult cells [3,4]. Mitotic segregation of old and new homologous chromosomes to daughter cells is presumed to be random. In principle, after S/G2-phase recombination between homologs, both recombinant chromatids may segregate to the same daughter cell (z segregation), or away from each other, each to a different daughter cell (x segregation), and only the former leads to LOH [57]. In Drosophila, however, recombination in S/G2 produces only z segregation, whereas G1 recombination is always followed by x segregation [58]. G1 recombination of chromosomes with heterozygous loci alters linkage relationships compared with parental cells but does not lead to LOH. Thus, different frequencies of LOH in particular cell types may be the consequence of the relative fraction of z segregation and may be related to the cell cycle compartment in which recombination occurs. Also, it could be related to variability of nonrandom segregation of original and newly replicated DNA strands [59] and may even vary between individual chromosome pairs within a specific single cell type. Thus, patterns of LOH might be directly related to organismal development and cell differentiation. An intriguing point of possible similarity to yeast is that 70% of yeast MR initiates in G1 [60,61].
The relatively low level of LOH seen in mESC, despite elevated HR and the consequent potential for elevated LOH, might be explained by our data indicating that much of the HR is initiated before DNA replication. In this way mESC would maintain a low mutation frequency through an error-free DNA repair pathway (HR), while assuring that at least one parental allele passes to the next cell generation without a new mutation in the DNA proximal to a DSB. In addition, MR in early embryonic cells could have 2 evolutionarily positive consequences: first, MR alters cis linkage of alleles (as in meiosis), thus increasing genetic variability in germ cells; second, LOH could restore homozygosity leading to restoration of normal cellular phenotype in cases of haploinsufficiency. We have previously speculated that such newly homozygous cells in some organ anlagen might then have a selective growth advantage during development [9].
Based on physical evidence coupled with the in vitro assays, our results suggest that HR in mESC compared to adult fibroblasts is the predominant DSB repair pathway, independent of both the source of DSB and the cell cycle phase at the time of DNA damage. This is dramatically different from what has been reported for other mammalian cells and intriguingly similar to reports from yeast. Selection during evolution seems to have tailored DNA repair strategies in a way that optimizes genetic outcome to subsequent cell fate.
Footnotes
Acknowledgments
We thank Dr. M. Brenneman and Dr. Shao for comments on the article. We thank Noriko Kane-Goldsmith for technical assistance. This work was supported by grants from the National Institutes of Health (ES011633 and P30ES005022), the National Aeronautics and Space Administration (NNG05GN24G), and a New Jersey Stem Cell Research grant from New Jersey Commission on Science and Technology.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.