
Abstract
Cultivation of functional pancreatic cells isolated from adult mammalian pancreas remains difficult. We developed a differentiation protocol that gradually induced the formation of mouse pancreatic exocrine cells from embryonic stem cells (ESCs). This process mimicked in vivo pancreatic development by directing cells through definitive endoderm (DE), gut tube endoderm, and pancreatic progenitor cells to differentiated cells that expressed pancreatic exocrine enzymes. Mouse ESCs were cultured in hanging drops to form embryoid bodies. Treatment of embryoid bodies with activin A induced the formation of DE cells that expressed marker mRNAs Goosecoid and Mixl1 and that were double-positive with Foxa2 and Sox17 proteins. Subsequent treatment of the DE cells by retinoic acid induced the formation of gut tube endoderm cells that expressed the specific marker Hnf1b. Expression of Goosecoid and Mixl1 was downregulated during this period. Fibroblast growth factor 7 (FGF7) promoted differentiation of PDX1-expressing pancreatic progenitor cells that also expressed Foxa2 mRNA, an endodermal marker, suggesting derivation from the DE cells. Exocrine cell differentiation was induced with FGF7, glucagon-like peptide-1, and nicotinamide. The differentiated cells expressed mature pancreatic exocrine cell mRNAs, such as Amylase, Elastase, and Carboxypeptidase A. Additionally, they produced pancreatic elastase, amylase, carboxypeptidase A, and chymotrypsin proteins that were identified in cytoplasmic granules by immunocytochemistry. Active amylase was released into the medium. Moreover, FGF7 was associated with differentiation of pancreatic exocrine cells. The findings reported here offer a novel and effective process to develop pancreatic exocrine cells from ESCs.
Introduction
P
The molecular mechanism(s) that promote the commitment of stem cells into pancreatic acinar cells remain largely unknown [4]. Activin A, a disulfide-stabilized protein that belongs to the transforming growth factor β (TGF-β) superfamily, is important for early definitive endoderm development and can induce ESCs to differentiate into them [5,6]. All-trans retinoic acid (RA) is a well-characterized signaling molecule that acts in anteroposterior patterning of neuroectoderm and mesoderm in vertebrates [7]. RA is also necessary for formation of the pancreas in Xenopus laevis, zebrafish, and mice [8 –10]. A recent study demonstrated that sequential treatment of mouse ESCs with activin A and RA could induce pancreatic progenitor cells [11]. Finally, fibroblast growth factor 7 (FGF7) is expressed in the pancreatic mesenchyme and promotes the growth, morphogenesis, and cytodifferentiation of pancreatic exocrine cells [12,13].
In this study, we developed a stepwise protocol to promote differentiation of mouse ESCs into functional pancreatic exocrine cells by mimicking signals that occur during embryonic development. This protocol used molecular signaling by activin A, RA, and FGF7 to direct cells through progressive stages resembling the definitive endoderm (DE) and the gut tube endoderm (GTE). It also induced the development of cells expressing Pdx1, a typical marker of pancreatic progenitor cells. Ultimately, Pdx1-positive cells differentiated into cells that expressed exocrine enzymes.
Materials and Methods
Cell culture and differentiation conditions
Undifferentiated mouse ESCs (129/sv strain; Cell and Molecular Technologies, Philipsburg, NJ) were maintained on mitomycin C-inactivated SNL feeder cell layers in 0.1% gelatin-coated tissue culture dishes. SNL feeder cells are STO feeder cells transformed with neomycin resistance and murine leukemia inhibitory factor genes. The cells were incubated in Dulbecco's modified Eagle's medium (DMEM; Gibco BRL, Rockville, MD) with 15% knockout-serum replacement (Gibco BRL), 100 μM nonessential amino acids (Gibco BRL), 1 mM sodium pyruvate (Gibco BRL), 103 U/mL of leukemia inhibitory factor (Chemicon, Temecula, CA), 100 μM 2-mercaptoethanol (Sigma, St. Louis, MO), 50 U/mL penicillin, and 50 μg/mL streptomycin (Gibco BRL).
To induce differentiation, the ESCs were cultured in feeder-free gelatin-coated tissue culture dishes using the hanging drop method to form embryoid bodies (EBs; Fig. 1). After 5 days, EBs were plated on gelatin-coated tissue culture dishes in EB medium composed of Iscove's modified Dulbecco's medium (Gibco BRL) supplemented with 20% fetal bovine serum, 100 μM nonessential amino acids, 1 mM sodium pyruvate, 50 U/mL penicillin, 50 μg/mL streptomycin, and 100 μM 2-mercaptoethanol. The following day, the EBs were incubated in EB medium with 100 ng/mL activin A (Sigma). After 24 h, activin A was removed, and the EBs were incubated 6 h in EB medium alone, followed by treatment with 1 μM RA (Sigma). After 24 h, the medium was switched to serum-free medium: DMEM/F12 (1:1) supplemented with N2 supplement (Invitrogen, Carlsbad, CA), B27 supplement (Invitrogen), and 15 ng/mL FGF7 (R&D Systems, Inc., Minneapolis, MN). After 3 days, the EBs were re-plated on gelatin-coated 24-well plates and incubated in DMEM/F12 supplemented with N2 supplement, B27 supplement, 15 ng/mL FGF7, 10 mM nicotinamide (NA; Sigma), and 100 ng/mL glucagon-like peptide-1 (GLP-1, 7–36 amide; Bachem AG, Torrance, CA) to induce differentiation of pancreatic exocrine cells (Fig. 1). In control cultures, the EBs were treated similarly but without inducing factors.
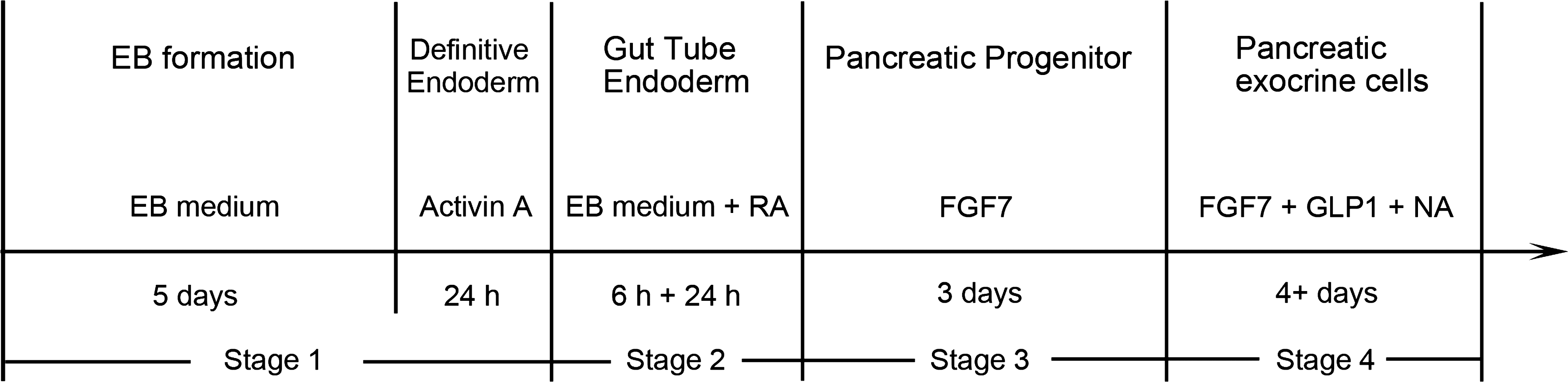
Experimental protocol for the differentiation of embryonic stem cells (ESCs) into pancreatic exocrine cells. The protocol was divided into 4 stages, including application of different growth factors at different periods. Undifferentiated mouse ESCs were cultured using the hanging drop method to induce formation of embryoid bodies (EBs). The EBs were incubated in EB medium supplemented with activin A, followed by treatment with all-trans retinoic acid (RA). Then EBs were cultured with fibroblast growth factor 7 (FGF7) for 3 days, and then re-plated and treated with FGF7, glucagon-like peptide-1 (GLP-1), and nicotinamide (NA).
Reverse transcriptase–polymerase chain reaction analysis
Total cellular RNA was isolated using TRIzol reagent (Invitrogen). Complementary DNA (cDNA) was synthesized from total RNA using a PrimeScript RT reagent Kit (Perfect Real Time; TaKaRa Bio Inc., Yokkaichi, Japan) for reverse transcriptase–polymerase chain reaction (PCR). The cDNA was amplified by PCR using specific primers (Table 1) for Foxa2, Goosecoid, Hnf1b, Hlxb9, Nkx6.1, Hnf6, Pdx1, Amylase, Elastase 1, Elastase 2, Carboxypeptidase A, Chymotrypsinogen B, P48, Mist-1, and β-actin.
The PCR was initiated by a 10 min denaturation at 94°C. This was followed by 35 cycles of heating at 94°C (30 s), 53°C–65°C (30 s), and 72°C (30 s). The final extension was for 7 min at 72°C. β-actin was used as the invariant control. Water was used as a negative control in place of the cDNA template.
Real-time PCR
Real-time PCR was carried out with primers (Table 2) for Foxa2, Goosecoid, Brachyury, Mixl1, Hnf1b, Pdx1, P48, Amylase, Carboxypeptidase A, Chymotrypsinogen B, Elastase 1, Ins2 (Insulin 2), Glucagon, and β-actin using a SYBR Premix Ex Taq (Perfect Real Time; TaKaRa Bio) in a Thermal Cycler Dice Real Time System (TaKaRa Bio). The cDNA was amplified by a 10 min initial denaturation at 95°C, followed by 40 cycles of heating at 95°C (5 s) and 60°C (30 s). The total RNA levels were normalized to levels of β-actin mRNA.
Immunofluorescence
For immunofluorescence staining, cultured cells were fixed in 4% paraformaldehyde in phosphate-buffered saline (pH 7.4) for 30 min, permeabilized with 0.1% Triton X-100 in phosphate-buffered saline, and treated with 1.5% normal donkey or goat serum. Then the cells were incubated with primary antibodies at 4°C overnight. Localization of antigens was observed with IgG secondary antibodies conjugated with fluorescein or TRITC (Alexa 488 and 568; Molecular Probes, Leiden, The Netherlands). Afterward, the cellular nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; Molecular Probes). The following antibodies were used for immunostaining: rabbit anti-Foxa2 (1:800, Cell Signaling Technology, Inc., Beverly, MA), mouse anti-Sox17 (1:100; R&D Systems, Inc.), rabbit anti-HNF1b (1:100; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), goat anti-PDX-1 (1:200, R&D Systems, Inc.), rabbit anti-α-amylase (1:500; Sigma), rabbit anti-pancreatic elastase (1:200, Elastin Products Co., Owensville, MO), goat anti-carboxypeptidase A1 (CPA1, 1:100; Santa Cruz Biotechnology), and mouse anti-chymotrypsin (1:500, Abcam, Cambridge, United Kingdom). Nonimmune serum was used as a negative control in place of the primary antibodies. Mouse fetal foregut and adult pancreas were used as positive controls to test the specificity of these antibodies.
To estimate the efficiency of differentiation at the cell level, clusters were randomly selected, and the numbers of positive cells were counted at different stages of differentiation. The total cell number was determined by DAPI nuclear staining. When appropriate, data are given as mean ± standard deviation obtained from at least 5 independent experiments.
Amylase activity assay
To test for the presence of secreted amylase activity, the induction medium was changed with a fresh induction medium. After incubation for 24 h, the conditioned medium was collected and centrifuged. Secreted amylase activity in the supernatant was measured with Liquid Amylase (CNPG3) Reagent Set (Pointe Scientific, Canton, MI) using α-amylase activity as the standard.
Results
Characterization of ESC-derived cells by early differentiation (Stages 1–3)
We developed a 4-step protocol for differentiation of ESCs to pancreatic exocrine enzyme-expressing cells. We focused first on generating DE, followed by PDX1-expressing cells, and finally, pancreatic exocrine cells. In Stage 1, mouse ESCs were transitioned through mesendoderm to DE by treatment of EBs with high concentrations of activin A (100 ng/mL). The mouse ESC-derived DE expressed Foxa2 and Goosecoid, and the expression levels of these markers were increased by treatment with activin A compared with nontreated cells (Fig. 2A, B). In addition, real-time PCR analysis indicated that Brachyury, a mesendodermal marker, was highly expressed in Stage 1 and then sharply decreased afterward. DE cell markers Sox17 and Foxa2 were also demonstrated by immunofluorescence (Fig. 2C).
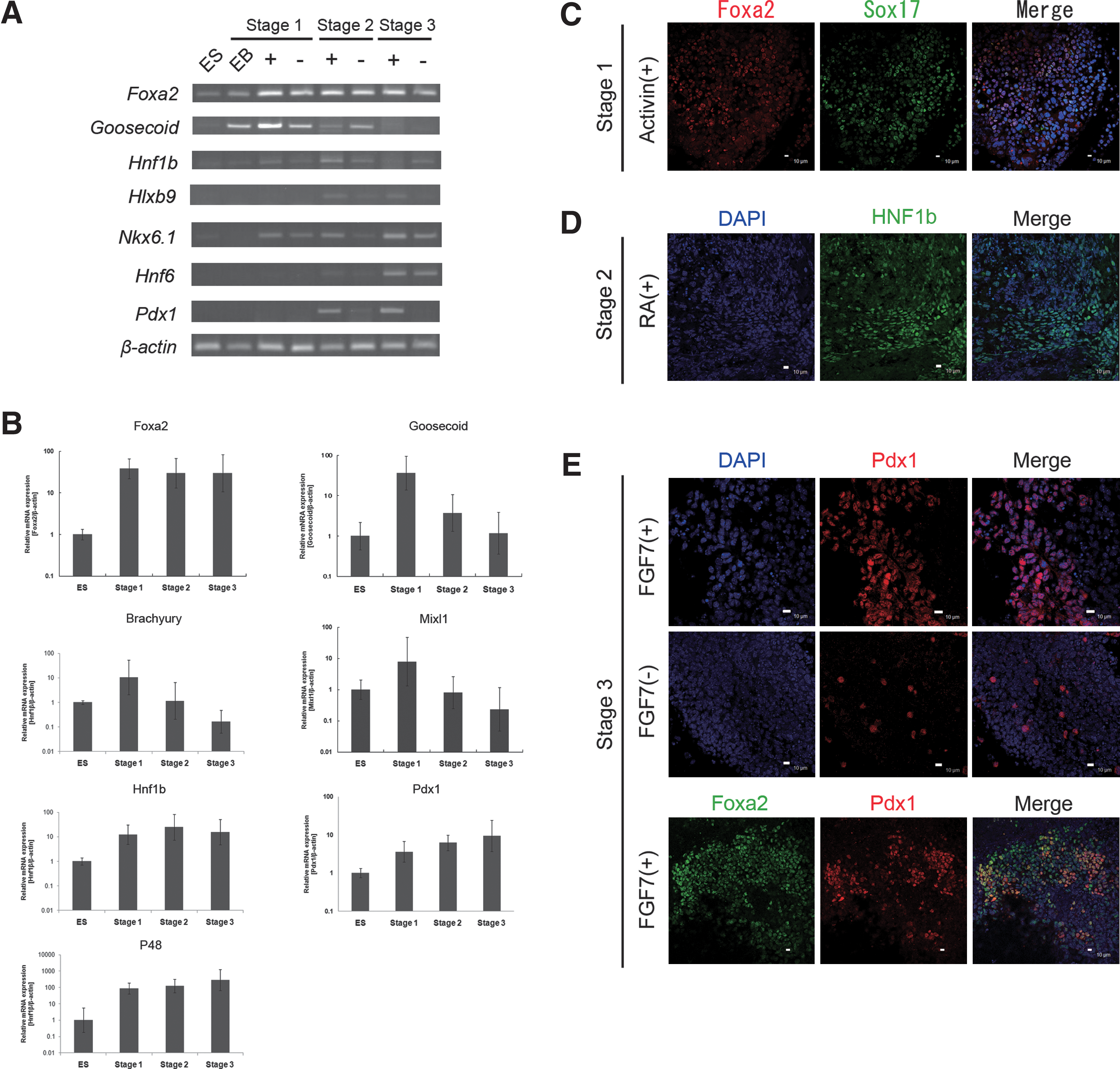
Derivation and characterization of mouse ESC-derived cells during early differentiation (Stages 1–3).
In Stage 2, activin A was removed, and 6 h later the cells were treated with RA. Expression of Goosecoid and Mixl1 mRNAs was downregulated (Fig. 2A, B). In contrast, expression of Hnf1b mRNA, a GTE marker, was upregulated (Fig. 2A, B). Consistent with the presence of Hnf1b mRNA, HNF1b-positive GTE-like cells were detected by immunostaining (Fig. 2D). Further, the pancreatic endoderm and progenitor cell marker Pdx1 mRNA was also upregulated in this step (Fig. 2A, B) along with other pancreatic endodermal markers Hlxb9 and Hnf6 mRNA and the pancreatic progenitor marker Nkx6.1 mRNA.
In Stage 3, the cells were switched to a serum-free medium containing FGF7 for 3 days. At the end of this period, the expression levels of the pancreatic progenitor cell markers Pdx1, Nkx6.1, and P48 were increased (Fig. 2A, B), and there was a increase in the level of Pdx1 (Fig. 2A, B). Moreover, immunostaining showed that PDX1 proteins were localized in the nuclei of FGF7-treated cells (Fig. 2E). Additionally, the PDX1-positive cells existed only in the Foxa2-positive cell population, suggesting the possibility that the PDX1-positive cells were derived exclusively from DE (Fig. 2E). Conversely, in the absence of FGF7, few cells expressed PDX1 (Fig. 2E), which in our cultures was essential to the transition of DE cells into pancreatic progenitor cells. This is consistent with studies revealing that FGF7 is important for pancreatic development [12,13].
Induction of pancreatic enzyme-expressing cells from progenitor cells (Stage 4)
During Stage 4, FGF7, GLP-1, and NA were used to recruit PDX1-expressing progenitor cells to the pancreatic exocrine lineage. Expression of Amylase, Elastase 1, Carboxypeptidase A, and Chymotrypsinogen B mRNAs were continued through Stage 4, and Elastase 2 mRNA was expressed later in Stage 4 (Fig. 3A). Amylase mRNA expression, used as a marker for pancreatic exocrine cells, was confirmed by real-time PCR on day 4, and it increased until day 25 (Fig. 3B). The pancreatic exocrine progenitor cell markers P48 and Mist-1 were also present on day 4 after induction (Fig. 3A), but the expression levels either held steady or decreased. These data suggested that the pancreatic progenitor cells differentiated into the mature pancreatic cells during the culture period. Expression of Amylase or Elastase 2 was not detected in control cells (Fig. 3A).
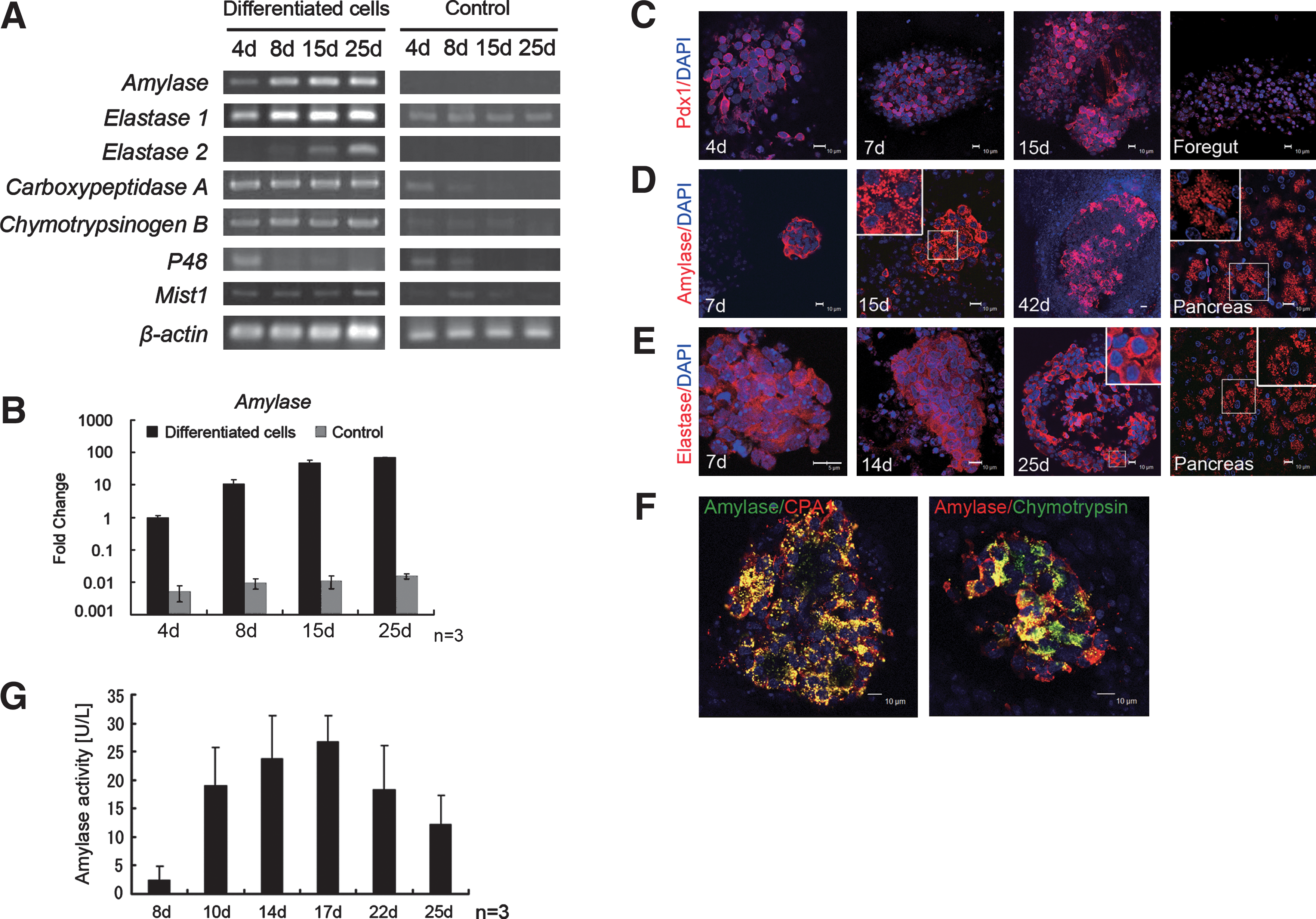
Characterization of mouse ESC-derived pancreatic exocrine cells by stepwise differentiation in Stage 4.
The presence of mRNAs associated with exocrine progenitor and differentiated cells does not necessarily mean that the proteins themselves were expressed. Therefore, we used immunocytochemistry to determine if they were. PDX1 continued to be expressed in the peripheral nucleoplasm of the induced cells at days 4, 7, and 15 (Fig. 3C). In the mouse fetal foregut at E11, PDX1 was localized to the nuclei (Fig. 3C). Amylase-positive cells were present at days 7, 15, and 42 (Fig. 3D), and elastase was detected at days 7, 14, and 25 (Fig. 3E). Expression of both enzymes increased during the culture period. The granular staining pattern, especially for amylase and elastase, was consistent with storage of digestive enzymes in the zymogen granules typically present in pancreatic acinar cells (Fig. 3D, E). The amylase and elastase-positive cells frequently formed clusters that were similar to structures found in the mature pancreas (Fig. 3D, E). Taken together, these findings suggested the differentiation of mature pancreatic exocrine cells from mouse ESCs.
To determine if the amylase-positive cells were actually pancreatic exocrine cells, we used double immunostaining for amylase and with CPA1 and with chymotrypsin. At the day 22 in Stage 4, most of the amylase-expressing cells were also positive for CPA1 or chymotrypsin (Fig. 3F). We then assessed physiological properties of the mouse ESC-derived exocrine cells by monitoring the activity of secreted amylase in the culture medium. Activity levels in the conditioned medium increased until day 17, but afterward decreased (Fig. 3G). These data showed that functional pancreatic cells were generated with our protocol.
Efficiency of stepwise differentiation and effect of FGF7 on the differentiation of pancreatic exocrine and endocrine cells
We estimated the efficiency of differentiated cell production at different periods by counting immunofluorescent-positive cells. Typically, for Foxa2-positive DE cells, an efficiency of 21.9% ± 4.5% was obtained by activin A treatment compared with nontreated cells, 4.3% ± 2.1% (P < 0.05, Fig. 4A). For pancreatic progenitor cells, PDX1-positive cells were present at a frequency of 12.0% ± 4.7% in FGF7-treated cells, which was a significantly greater than for nontreated cells, 1.7% ± 1.7% (P < 0.05, Fig. 4B). At day 15 in Stage 4, 10.7% ± 7.4% of the cells stimulated with FGF7, GLP-1, and NA expressed amylase, whereas only 1.3% ± 1.8% did in the absence of FGF7 but with GLP-1 and NA (P < 0.05, Fig. 4C). Thus, these growth factors play important roles for pancreatic differentiation and, as a result, improved the differentiation efficiency at various periods.
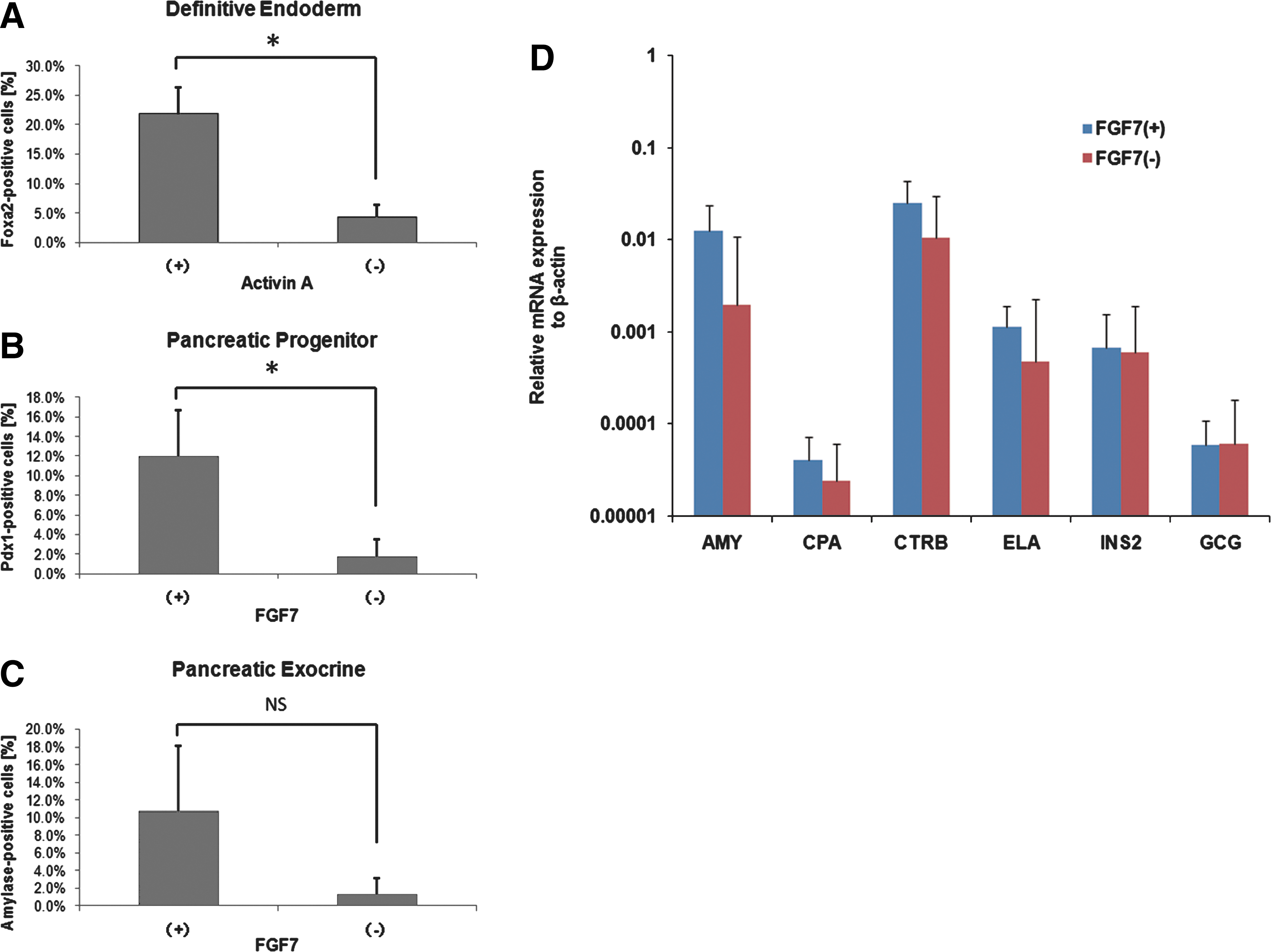
Efficiency of stepwise differentiation and effect of FGF7 on the differentiation of pancreatic exocrine and endocrine cells.
We next tested the differentiation of ESC-derived pancreatic progenitor cells with and without FGF7 to confirm the effect of FGF7 on pancreatic exocrine differentiation in the final stage. Expression of Amylase, Carboxypeptidase A, Chymotrypsinogen, and Elastase mRNAs tended to be greater in cells treated with FGF7 than those without it, but the differences were not statistically significant (Fig. 4D). On the other hand, the pancreatic endocrine markers Ins2 and Glucagon remained unchanged under both conditions (Fig. 4D).
Discussion
Although there are many reports of differentiation of pancreatic endocrine cells from ESCs, only a few reports demonstrate the presence of differentiated exocrine cells [2,14,15]. Both endocrine and exocrine pancreatic cells are derived from common progenitor cells that express PDX1 [16,17]. The molecular mechanism for the differentiation of exocrine cells is poorly understood, so the effective factor(s) that induce exocrine differentiation from cultured ESCs is not known. We assumed that the growth factors effective for endocrine cell differentiation also had the potential to produce exocrine cells. In the present study, we developed a novel stepwise differentiation protocol using these growth factors. Moreover, we demonstrated the necessity of FGF7 for producing PDX1-positive pancreatic progenitor cells. The protocol described here consistently generated pancreatic exocrine enzyme-expressing cells from mouse ESCs, and these exocrine cells secreted active α-amylase.
A previous report suggested that TGF-β is necessary for formation of endoderm in vertebrates [18]. Activin A, a member of the TGF-β superfamily, induces expression of many endodermal genes, including Mixl1 and Goosecoid, that are important for early endoderm development [19,20]. Indeed in Stage 1 of the present study, the cells expressed endodermal markers Foxa2, Goosecoid, and Mixl1, and both Foxa2 and Sox17 proteins were present. Further, the expression level of mRNA for Brachyury was very high in this stage, suggesting that the cells had transited through the mesendoderm stage.
In Stage 2, RA induced differentiation of GTE from DE as demonstrated by downregulation of Goosecoid and Mixl1 and upregulation of Hnf1b and Pdx1. In addition, expression of pancreatic endodermal markers Hlxb9 and Hnf6 was initially detected. RA plays a key role in specification of the foregut endoderm that becomes pancreas [10], and it regulates the commitment of ESCs to form PDX1-positive pancreatic endoderm in vitro [21]. However, in long-term cultures, RA suppresses the differentiation of exocrine cells from embryonic pancreas [22]. We improved this procedure by shortening the RA treatment to 24 h.
PDX1 is essential for determination and differentiation of the posterior foregut, in particular regarding the proliferation and differentiation of the pancreatic progenitor cells [23,24]. Thus, the induction of PDX1-positive cells must be a necessary step for differentiation of pancreatic exocrine cells. In Stage 3, the treatment of FGF7 resulted in a large increase in expression of PDX1. Some reports showed that several FGFs are expressed in pancreatic rudiments where they participate in epithelial–mesenchymal interactions [12,13]. In particular, FGF7 stimulates both the proliferation of embryonic pancreatic epithelial cells and the development into acinar exocrine cells expressing amylase [13]. Further, FGF7 upregulates expression of pancreatic markers, including Pdx1 in the developing EB [3]. Our results similarly showed FGF7 promotion of pancreatic progenitor cell differentiation. In addition to increased expression of Pdx1, pancreatic progenitor markers Hnf6, Nkx6.1, and P48 were also notably upregulated by FGF7. Additionally, these PDX1-positive cells were also positive for Foxa2, suggesting that they were derived from DE. It is indicated that FGF7 has been demonstrated to play a key role in the differentiation of GTE into pancreatic progenitor cells and then into pancreatic exocrine cells in vitro.
In Stage 4, ESC-derived PDX1-positive cells treated with FGF7, GLP-1, and NA became differentiated into functional pancreatic exocrine cells. The molecular mechanisms by which pancreatic exocrine cell differentiation occurs are largely unknown. In the absence of FGF7 in vitro, most expanded progenitor cells differentiate into endocrine cells [25]; therefore, FGF7 may play a crucial role for formation of exocrine cells by suppressing endocrine cell differentiation. Our results are consistent with the importance of FGF7 in the induction of exocrine differentiation. However, the effect of FGF7 was limited because ESC-derived nonpancreatic cells also continued to exist in this culture. GLP-1 and NA are usually used as growth factors for pancreatic endocrine differentiation [26, 27]. GLP-1 is associated with increased cell proliferation of both the endocrine and the exocrine component of the pancreas [28]. Additionally, a previous study showed that the combination with GLP-1 and NA induced both exocrine and endocrine gene expression [29]. Our data are consistent with the idea that the combination of FGF7, GLP-1, and NA induces pancreatic exocrine cell differentiation from mouse ESCs.
In the final stage of differentiation, the transcription factor PDX1 was localized in the peripheral nucleoplasm although that was localized in nucleoplasm in earlier stages. The translocation of PDX1 from the nuclear periphery into the nucleoplasm is associated with the activation of pancreatic cells [30]. As for the adult pancreas, PDX1 is expressed in endocrine but not in exocrine tissues [31]. Thus, the movement of PDX1 into the peripheral nucleoplasm might promote the differentiation of exocrine cells in this system.
The differentiated cells secreted increasing amounts of amylase into the medium through day 17 of culture. After that, the secretion diminished, which is consistent with the known difficulty of maintaining differentiated exocrine pancreas in long-term culture. At the same time, expression of P48 mRNA, a marker of pancreatic exocrine progenitor cells, decreased in Stage 4. The role of FGF7 in promoting differentiation was subtle. When present, it tended to increase the number of amylase-positive cells, but the results were not statistically significant. These findings suggest that further pancreatic exocrine differentiation requires other signals in addition to FGF7.
A major contribution of this works is that we generated pancreatic exocrine cells from mouse ESCs. The combination of growth factors that we used promoted the recapitulation of embryonic developmental processes by the ESCs. This system offers a new induction model for studying the mechanism of pancreatic exocrine cell formation and differentiation in vitro. In our future works, we will improve the differentiation efficiency via an inductive or a selective mechanism, and then these differentiated cells will be available for use in digestion assays and cytotoxicity evaluation. There is even the possibility that this methodology will have applications in regenerative medicine. In future works, we will clarify the individual roles of FGF7, GLP-1, NA, and other factors in pancreatic exocrine cell differentiation.
Footnotes
Acknowledgments
The authors thank Ms. Kayo Suzuki and Dr. Kiyokazu Kametani (Research Center for Instrumental Analysis, Shinshu University) for their outstanding technical assistance.
Author Disclosure Statement
No competing financial interests exist.