
Abstract
Snail genes are transcriptional repressors well known to play important roles in epithelial to mesenchymal transitions during both embryogenesis and cancer metastasis. Although they are generally regarded as markers of mesenchymal cells, Snail genes have also recently been implicated in regulating stem cell populations in both Drosophila and vertebrates. In this study we investigate Snai1, a member of the mouse Snail family, in the intestinal stem cell niche and examine the relationship between canonical Wnt signaling, a key regulatory pathway of intestinal stem cells, and expression and cellular localization of Snai1. Strong nuclear expression of Snai1 was detected in the crypt base columnar stem cells in the adult small intestine while Snai1 was mostly found in the cytoplasm of differentiated enterocytes and enteroendocrine cells. Expression and cellular localization of Snai1 in the intestinal epithelium appears to be regulated by the canonical Wnt signaling pathway as Snai1 expression was dramatically reduced after conditional deletion of β-catenin. Conversely, significant nuclear Snai1 was detected in polyps derived from Apcmin mice and in intestinal villi after conditional mutation of Apc in AhCre, Apcf/f mice, indicating that upregulation of the Wnt pathway in the intestinal epithelium induces both increased expression and nuclear localization of Snai1.
Introduction
W
Snail genes are transcriptional repressors well known for their role in triggering epithelial to mesenchymal transition (EMT) during embryonic development [17 –21] and metastatic phenotypes in the progression of cancer, including colorectal carcinomas [22,23]. Intriguingly, several recent reports have suggested that cells that undergo EMT acquire stem cell-like characteristics [24 –26]. This raises the question of whether genes implicated in EMT also play a role in stem cell populations within normal regenerating tissues. Members of the Snail protein family have been reported to be expressed in several multipotent cell populations, including Drosophila testicular somatic [27], neural [28] and intestinal stem cells [29], and vertebrate mammary stem cells [24]. This suggests that Snail transcriptional repressors may also have a role in regulating stem or progenitor cell fate. The Snail family of zinc-finger DNA-binding proteins bind to E-boxes in the promoter of target genes, thereby repressing gene expression [30,31]. Three equally related family members are present in mice and humans (Snai1, Snai2, and Snai3) [18]. Mouse and Drosophila Snail proteins have been implicated in the regulation of cellular differentiation, proliferation, and survival and act to downregulate E-cadherin expression during gastrulation [19,32] and metastasis. An increase in expression of Snai1 and Snai2 has been reported in several studies of colorectal carcinomas, but little is known about their role in normal intestinal tissue. Here, we examine cellular expression of Snai1 in the intestinal stem cell niche and show that both transcript levels and protein subcellular localization are regulated by Wnt-β-catenin signaling.
Materials and Methods
Ethics statement
All animals were handled in strict accordance with good animal practice as defined by the relevant national and/or local animal welfare bodies, and all animal work was approved by the appropriate committee.
Mice
Intestinal tissue from wild-type C57BL6 mice was harvested for expression analysis. Lgr5-EGFP-Ires-CreERT2 mice [5] were utilized to detect Lgr5 expressing crypt base columnar (CBC) stem cells (obtained from N. Barker and H. Clevers, Utrecht, The Netherlands). Apcmin mice [33] were used to analyze Snai1 expression in polyp tissue. Conditional deletion of β-catenin and Apc in the intestinal epithelium was achieved by crossing Ctnnb1fl/fl [34] (obtained from J. Huelsken ISREC, Switzerland) and Apcfl/fl to AhCre (obtained from A. Clarke, Cardiff, United Kingdom) mice as previously described [35,36]. Expression of Cre recombinase was induced by giving control and test mice 4 daily intraperitoneal injections of β-naphthoflavone (80 mg per kg). On day 5 the mice were killed and intestinal tissue was harvested for analysis.
Epithelial cell preparation for whole-mount immunohistochemistry and RNA isolation
Small intestinal tracts were cut open longitudinally and washed in phosphate-buffered saline (PBS) before incubation in PBS containing 3 mM ethylenediaminetetraacetic acid and 0.5 mM dithiothreitol (DTT) for up to 90 min at 4°C. After gently rinsing twice in PBS, vigorous shaking for a couple of seconds yielded crypts and villi. For whole-mount immunohistochemistry, crypts from 2 Lgr5-EGFP-Ires-CreERT2 mice were fixed for 5 min in 4% paraformaldehyde (PFA), washed in PBS, and stored at 4°C until further use. For RNA isolation, cells were spun down at 800 rpm for 5 min at 4°C and the cell pellet subsequently homogenized in Trizol® Reagent (Invitrogen).
RNA isolation and quantitative reverse transcriptase polymerase chain reaction analysis
Total RNA was isolated from preparations of isolated epithelial crypts from adult mouse colon and Apcmin polyp tissue. Crypts from 4 wild type and polyps from 4 Apcmin mice were used to prepare RNA for analysis. Further RNA cleanup and DNase I digestion was performed using the RNeasy Kit (Qiagen) and cDNA prepared using Superscript III Reverse Transcriptase (Invitrogen) using the random hexamer method. Quantitative polymerase chain reaction (PCR) was performed using Platinum Taq Polymerase (Invitrogen) and Sybr Green (Fisher) using a RotorGene 3000 cycler (Corbett Research). The primers used were specific for mouse Snai1 (forward, 5′-aaacccactcggatgtgaag; reverse, 5′-gaaggagtcctggcagtgag), mouse Snai2 (forward, 5′-cctttctcttgccctcactg; reverse, 5′-acagcagccagactcctcat), and mouse Snai3 (forward, 5′-ttctcccgcatgtctctctt; reverse, 5′-gcaggacacaggtaacagtg). Amplification conditions were 95°C for 10 min followed by 45 cycles of 95°C for 20 s, 58°C for 30 s, and 72°C for 30 s. Individual samples were analyzed in triplicate. Expression was normalized relative to 18S rRNA expression. A Student's t-test was performed to verify statistical significance.
Histology and immunohistochemistry
Intestinal tissue for immunohistochemistry and Periodic Acid Schiff staining was collected, washed in PBS, fixed overnight in 4% PFA at 4°C, processed, embedded in paraffin, and sectioned at 5 μm. Immunohistochemistry was performed using standard protocols. Slides were dewaxed, dehydrated, and incubated in 0.5% H2O2 in methanol for 30 min. After rehydration, slides were heated for 10 min in a pressure cooker in 10 mM citrate buffer (pH 6.0) and blocked with 5% milk powder in PBS (for Snai1 immunohistochemistry) or CAS block (Invitrogen) for 1 h. Sections were incubated overnight at 4°C with the primary antibody in 5% milk powder in PBS (for Snai1 immunohistochemistry) or in PBS with 0.2% bovine serum albumin. The Snai1 antibody (1:400; Abcam, ab17723) was used together with Snai1 peptide, to control for nonspecific staining (1:75, ab19126) in peptide competition assays. Antibodies were used to detect Lysozyme (Dako;A0099 1:400) and β-catenin (BD Transduction Laboratories; 610154 1:500). For detection of the primary antibodies, anti-mouse or anti-rabbit horseradish peroxidase- or alkaline phosphatase-coupled antibodies (1:200, all from Invitrogen) in PBS with 0.2% bovine serum albumin were incubated for 1 h at room temperature. Peroxidase activity was detected with the 3, 3′-diaminobenzidine liquid kit from Dako. Phosphatase activity was detected with the 5-Bromo-4-Chloro-3-Indolyl phosphate, and Nitroblue tetrazolium salt staining kit from Invitrogen. Sections were counterstained with haematoxylin where appropriate, dehydrated, and mounted. For immunofluorescent detection anti-rabbit Alexa 488, anti-rabbit Alexa 594, anti-mouse Alexa 594, anti-GFP 488-conjugated antibodies, and 4′,6-diamidino-2-phenylindole (DAPI) nuclear stain (all Invitrogen) were used. Sections were mounted with Prolong Gold (Invitrogen).
Imaging was carried out using a Zeiss Axioimager microscope running Axio Vision Rel. 4.7 Software (Carl Zeiss), Nikon C1 (Upright) confocal microscope running NIS Elements Software (Nikon), and Leica SP5 confocal microscope (Leica).
Quantification of nuclear and cytoplasmic signal intensities
Tissue from AhCre control and AhCre, Apcfl/fl mice was harvested after induction with β-naphthoflavone, sectioning, and immunohistochemistry for Snai1 was as described above. Control and test samples were processed under identical conditions and 3 samples of each were analyzed. Bright-field images were exported as gray-scale TIFF files, and signal intensities in the nuclear and cytoplasmic regions were quantified using Image J. The ratio of the nucleo-cytoplasmic signal intensity was calculated from the mean signal intensities measured within a small circular region of interest placed within the nucleus and cytoplasm of 20 cells per sample. A Student's t-test was performed to verify statistical significance between the 2 groups.
Results
Snai1 is strongly expressed in the nuclei of CBC intestinal stem cells and their immediate progeny
Since Snail genes are primarily markers of mesenchymal tissue it was initially surprising to observe Snai1 expression in the intestinal epithelium. Snai1 expression was detected throughout the crypt villus axis but demonstrated a restricted pattern of expression within the crypts of the adult small intestine, where Snai1 was detected at higher levels and localized to the nuclei of many cells (Fig. 1A). Closer examination revealed that Snai1 was expressed in cells that had morphology corresponding to the CBC epithelial stem cells that have elongated nuclei compared to the round nuclei of adjacent Paneth cells (Fig. 1B). Strong nuclear expression of Snai1 was also detected in cells at the base of colonic crypts (Fig. 1C). Snai1 was also detected in transient amplifying cells and differentiated cells in the intestinal epithelium. Analysis of Snai1 expression using immunofluorescence co-stained with DAPI (Fig. 1A, D–I) revealed that Snai1 has a more prominent nuclear localization in the crypt base (Fig. 1G) compared to a more uniformly distributed expression throughout the cell in enterocytes located in the villi of the small intestine (Fig. 1D).
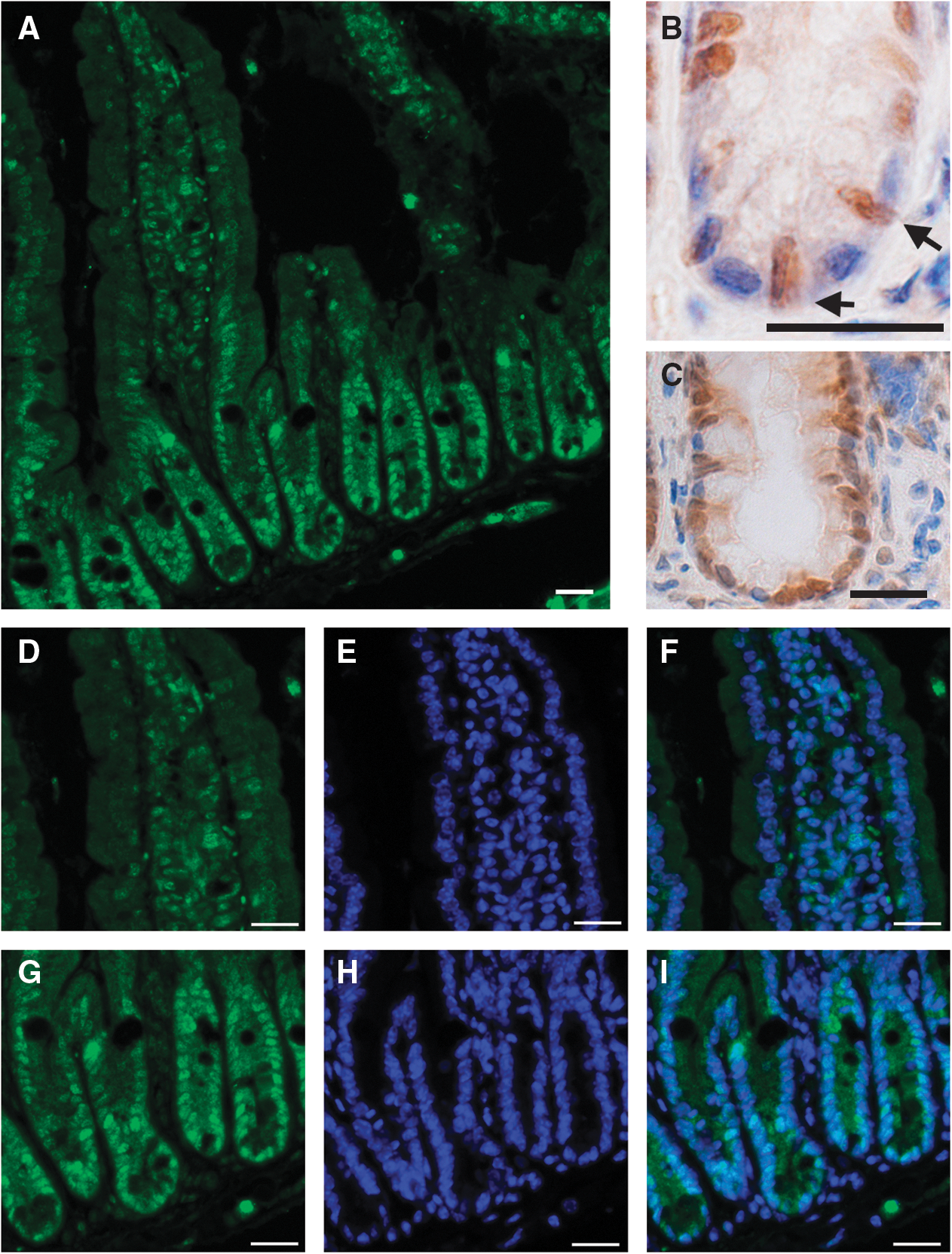
Snai1 is highly expressed and localized to the nucleus in the adult intestinal stem cells and their progeny. Immunostaining for Snai1 shows expression in the adult small intestine
CBC cells are known to express the stem cell marker Lgr5 (Leucine-rich-repeat-containing-G protein coupled receptor 5) [5] and Snai1 expression appeared to be present in these cells and notably absent in the adjacent differentiated Paneth cells (Fig. 1B). To confirm that Snai1 expression was localized to the CBC intestinal stem cells in the small intestine, we performed co-localization studies using crypts prepared from Lgr5-EGFP-Ires-CreERT2 mice where the Lgr5-positive CBC stem cells are marked with green fluorescent protein [5]. Whole-mount immunofluorescent staining for Snai1 and enhanced green fluorescent protein revealed that Snai1 and Lgr5 co-localize in cells at the base of crypts, confirming that Snai1 is expressed in CBC stem cells (Fig. 2).

Snai1 is expressed in CBC cells. Confocal imaging analysis of whole-mount immunostaining for Snai1 (red)
To investigate Snai1 expression in specific differentiated epithelial cell populations in more detail, we performed double-labeling experiments. Immunostaining for Snai1 followed by Periodic Acid Schiff staining to detect mucin secreting goblet cells revealed that goblet cells do not express Snai1 (Fig. 3A). Also Paneth cells detected by staining for Lysozyme confirmed the absence of Snai1 (Fig. 3B). In contrast, double immunofluorecent staining for Snai1 and Synaptophysin to detect enteroendocrine cells clearly demonstrated expression of Synaptophysin and Snai1 in these cells (Fig. 4). Interestingly, Snai1 expression was found predominantly in the cytoplasm of the enteroendocrine cells and not in the nucleus, suggesting that Snai1 is not functioning as an active transcriptional repressor in these cells. Analysis of Snai1 expression in mature enterocytes also showed a significant level of protein in the cytoplasm of cells (Figs. 1 and 3B).

Snai1 is expressed in absorptive cells of the intestinal epithelium but not in secretory Paneth and Goblet cells.
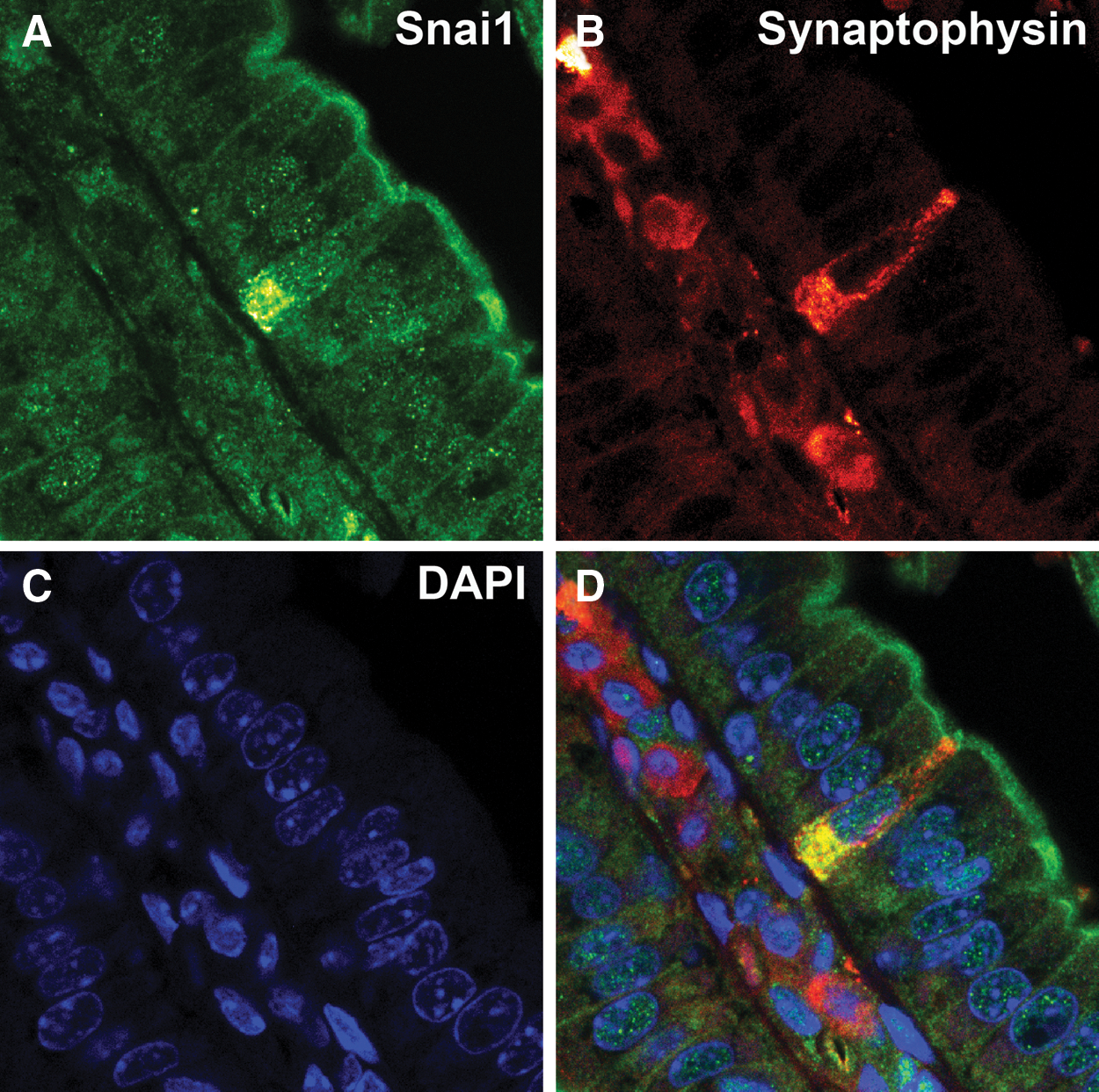
Snai1 is expressed in enteroendocrine cells but expression is largely cytoplasmic. Double-labeling for Snai1
The canonical Wnt signaling pathway regulates Snai1 expression
The Wnt signaling pathway has been shown to have a critical role in regulating maintenance of intestinal stem cells [16,37,38]. Further, there have been several reports linking canonical Wnt signaling and the regulation of Snai1 expression, protein localization, and stability [39 –41]. Our observation that Snai1 was highly expressed and specifically localized to the nuclei at the base of crypts where Wnt signaling is most active [1,42] prompted us to examine whether Snai1 is influenced by canonical Wnt signaling in the intestinal epithelium. We initially investigated the relationship between Wnt signaling and Snai1 by examining expression of Snai1 in Apcmin polyps. Apcmin mice harbor a germline truncating mutation at codon 850 in the Apc gene and are susceptible to polyp formation in both the small intestine and colon [33]. Apc is a tumor suppressor gene that has been well studied as a negative regulator of the Wnt signaling pathway where Apc is part of a protein complex that negatively regulates β-catenin activity [35,37] by targeting it for degradation via the ubiquitin proteasome pathway [43]. Polyps that form in the intestine of Apcmin mice exhibit high levels of β-catenin expression in the cytoplasm and nucleus of cells [1]. We therefore examined expression of Snai1 in Apcmin polyps using both quantitative reverse transcriptase-PCR (Fig. 5A) and immunohistochemistry (Fig. 5B). Quantitative reverse transcriptase-PCR analysis revealed a significantly higher level of expression of Snai1 (P ≤ 0.00015) as well as induction of Snai3 (P ≤ 0.0025) in Apcmin polyps compared to wild-type epithelial tissue (Fig. 5A). Snai1 immnuohistochemistry showed stronger nuclear Snai1 expression in the Apcmin polyp tissue (Fig. 5B) than wild-type colonic epithelium (Fig. 5C). This was investigated further in the small intestine by using genetic mouse models where Wnt signaling is constitutively activated (AhCre, Apcfl/fl [35]) and blocked (AhCre, Ctnnb1fl/fl [36]) throughout the intestinal epithelium. Immunohistochemical staining for Snai1 in small intestinal tissue from AhCre, Apcfl/fl mice showed stronger nuclear Snai1 expression in epithelial cells in the villi (Fig. 6D, E) than the Snai1 staining of control (AhCre, Apc+/+ ) tissue from littermates (Fig. 6B, E). This finding correlates with strong nuclear expression of β-catenin, a marker for active Wnt signaling, in epithelial cells lining the villi of the knockout tissue (Fig. 6C) in contrast to membrane staining of the control tissue (Fig. 6A; note nuclei in this panel only stain blue with the hemotoxylin counterstain and are not positive for β-catenin).
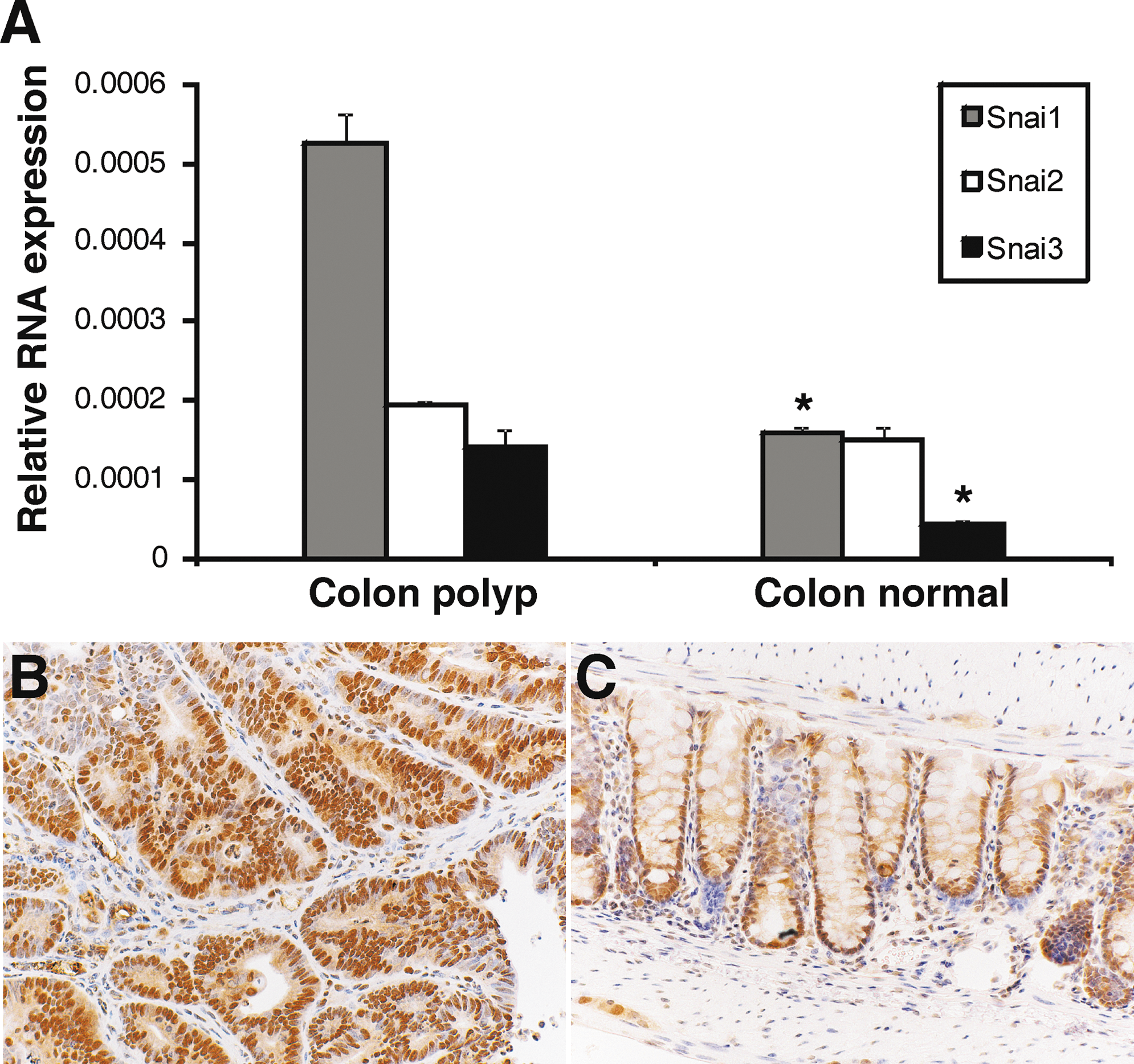
Intestinal polyps from Apcmin
mice exhibit increased nuclear Snai1 expression.
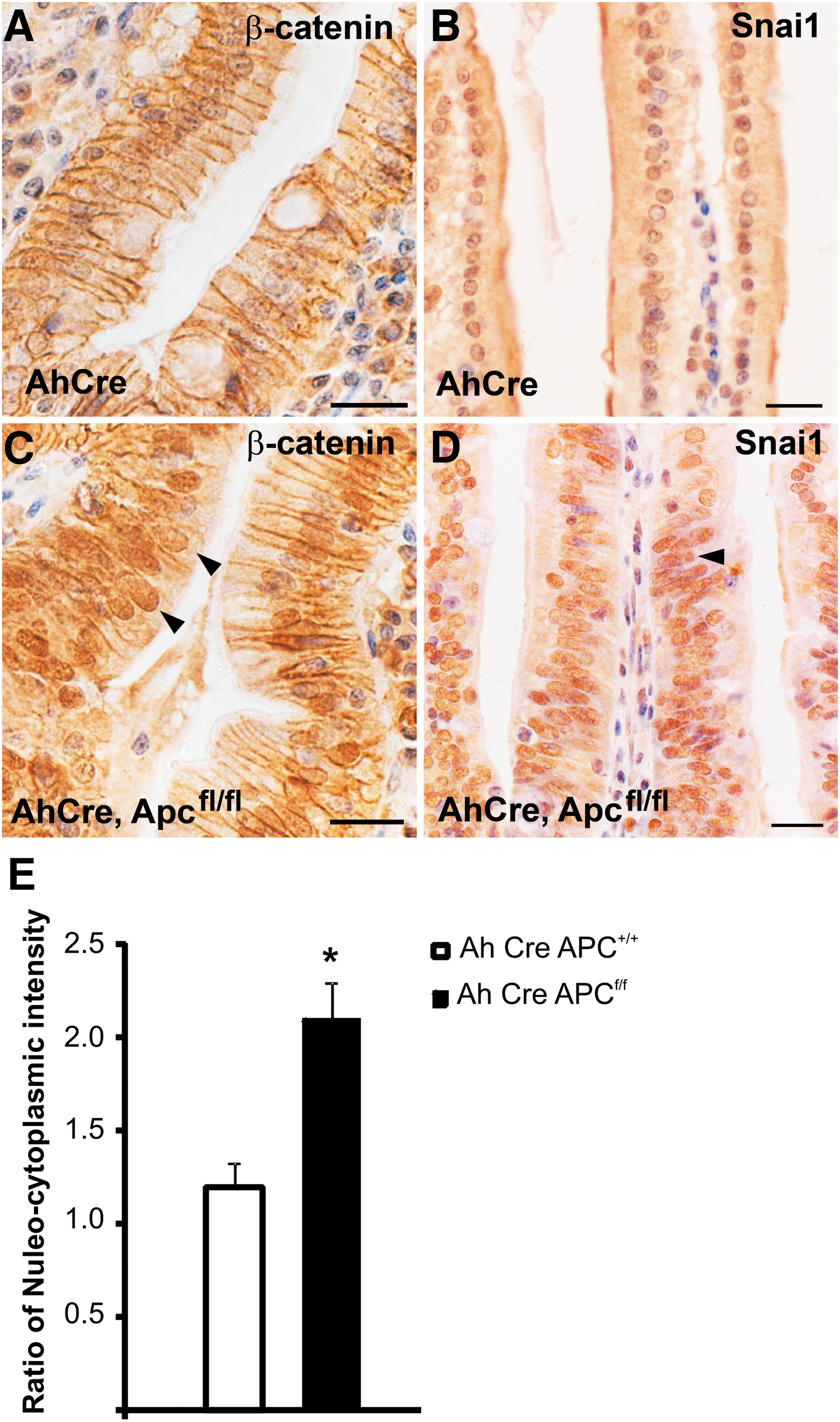
Activation of Wnt signaling produced by Apc mutation results in an increase in Snai1 expression and enhancement of nuclear localization. Immunohistochemical staining for Snai1 in
In the converse experiment, Snai1 expression was dramatically reduced in tissue from AhCre, Ctnnb1fl/fl mice where Wnt signaling is blocked by conditional deletion of β-catenin (Fig. 7C, D). Snai1 expression is considerably reduced even in cells at the base of crypts that normally display most prominent nuclear expression of β-catenin (compare panels A and B in Fig. 7 with those of panels C and D). A few cells expressing nuclear β-catenin can be seen in a crypt from the AhCre, Ctnnb1fl/fl mouse (Fig. 7C, asterisk). This is due to the induction of the Cre transgene in the AhCre mouse not being 100% efficient. An area where β-catenin is deleted as shown by immunohistochemistry (Fig. 7C, circled) clearly has reduced expression of Snai1 (Fig. 7D, circled compared to 7B). These results show that both expression and cellular localization of Snai1 is regulated by canonical Wnt signaling.
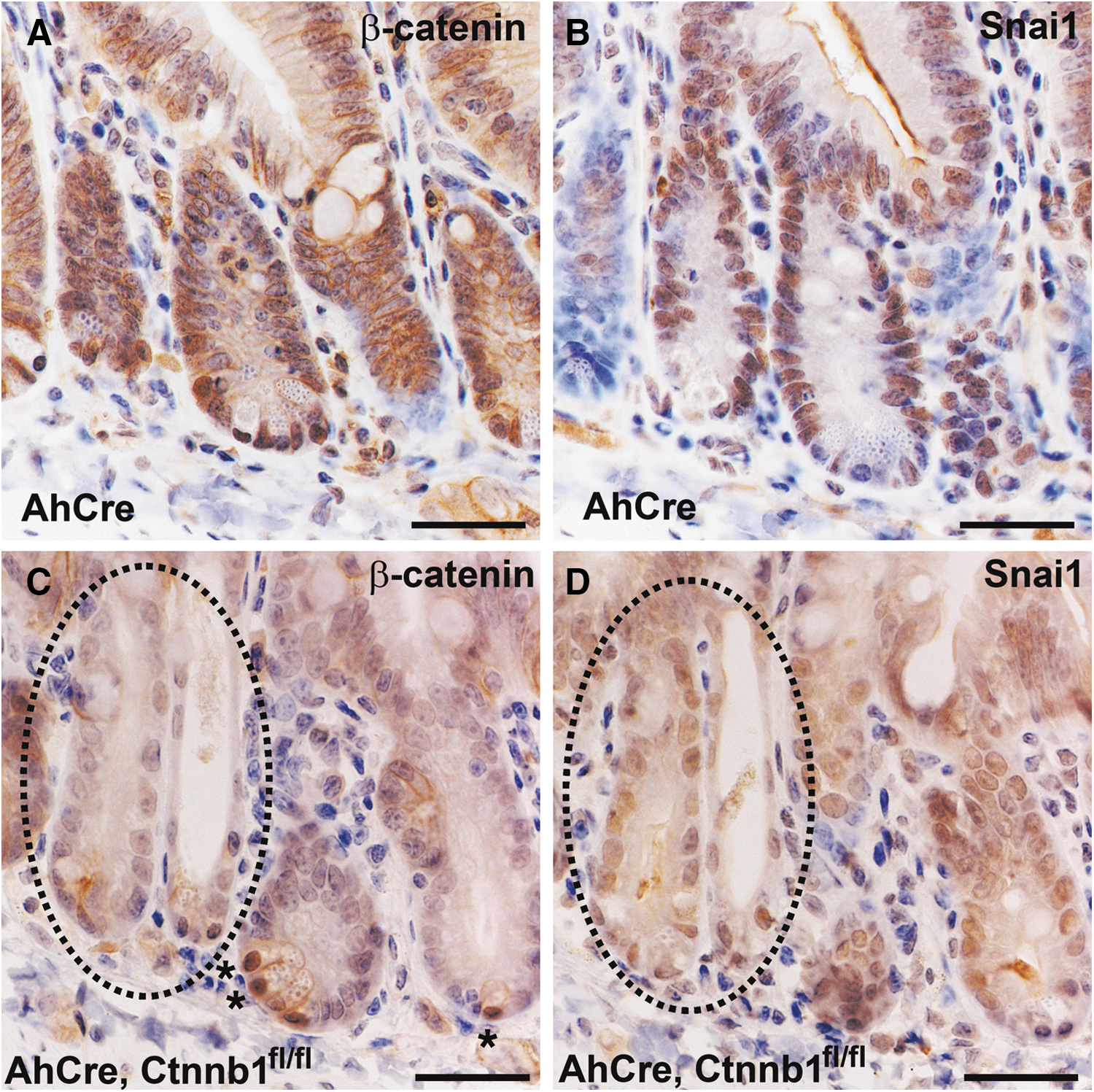
Inhibition of canonical Wnt signaling via mutation of β-catenin results in reduced Snai1 expression. Tissue was collected for immunohistochemistry from
Discussion
Expression of Snai1 is localized to cells within the epithelium. Intriguingly, most distinctive nuclear expression of Snai1 was detected in CBC intestinal stem cells and immediate progeny. Snail proteins have been associated with metastasis of intestinal tumors and indeed expression of Snai1 has been shown to have negative correlations with patient outcome [22,23]. Expression of Snai1 within the CBC cells of normal epithelial tissue suggests that Snai1 may have another function in addition to its role in the EMT that is a hallmark of epithelial tumor metastasis [26]. This relatively late function of Snai1 in tumor progression may be preceded by a role in promotion of stemness or generation of cancer stem cells. Intestinal epithelial tumors in mice have been shown to arise from the CBC cells, the stem cells that regenerate normal intestine [44]. Our results show that these same cells express high levels of nuclear Snai1 in a Wnt pathway-responsive manner. Wnt signaling upregulates the total cellular level of Snai1 and also promotes nuclear localization. As Snai1 is a transcriptional repressor that requires nuclear localization to function; this implies that Wnt signaling also regulates Snai1 function post-translationally to prevent export of Snai1 from the nucleus. Cell culture studies have demonstrated that the promotion of nuclear localization is not a β-catenin-dependent function but is due to the inhibition of glycogen synthase kinase-3β (GSK3β) activity that occurs upon Wnt activation. GSK3β directly phosphorylates Snai1, which promotes its nuclear export [41]. Axin 2 has also been shown to act as a nucleocytoplasmic chaperone for GSK3β [40].
Enteroendocrine cell differentiation has been shown to be independent of Wnt signaling [45], and our results clearly demonstrate that Snai1 was mainly cytoplasmic in this cell type. Enteroendocrine differentiation is dependent upon induction of 2 basic Helix-Loop-Helix proteins, Neurogenin-3 and NeuroD1, via Notch pathway activation [46,47]. Basic Helix-Loop-Helix proteins compete for the same target sites (E-boxes) [30] on target gene promoters as Snail proteins; hence, Snai1 may actively need to be kept out of the nucleus for differentiation of enteroendocrine cells.
Wnt signaling has been found not only to be essential for the maintenance of intestinal stem cells but has a critical role in the differentiation of the Paneth cell lineage [13]. CBC cells are located between Paneth cells at the base of crypts in the small intestine and both stem cells and Paneth cells exhibit nuclear localization of β-catenin indicative of active Wnt signaling [38]. In our study Snai1 was strongly expressed in CBC cells but notably absent from adjacent Paneth cells. This suggests that additional signals distinctive from Wnt signaling are also required for expression of Snai1. This is supported by our expression studies where we show that Snai1 expression is significantly downregulated when Wnt signaling is blocked but is not completely absent. Studies in other systems have shown that Snai1 expression can be regulated by multiple signaling pathways in different contexts, including Notch, transforming growth factor β, and fibroblast growth factor signaling pathways, so it is possible that Snai1 may also be regulated by these factors in the intestinal epithelium [21].
It was initially surprising to find Snai1 in the intestinal epithelial stem cell population, but several studies have now indicated that Snail proteins may play a role in promoting a stem cell state in epithelial tissues. Snai1 is expressed in developing hair buds in mice, and Snai1 misexpression in the epidermis leads to an expansion of epidermal progenitors and a reduction in intercellular adhesion [48]. Further, normal mammary stem cells express Snai1, and ectopic expression of Snai1 in human mammary epithelial cells leads to the acquisition of many properties of mammary stem cells [24]. Since metastatic cells not only migrate but also invade and proliferate at different sites, at least a subset of them may have some stem cell-like properties. Snai1 is a novel target of the Wnt pathway within the intestinal epithelium and may represent a link between Wnt signaling, epithelial stem cells, and metastatic cancer cells.
Footnotes
Acknowledgments
This work was supported by National Health and Medical Research Council (NH&MRC) Australia project grants to H.E.A. and G.R.H.
Author Disclosure Statement
No competing financial interests exist.