
Abstract
Hematopoiesis during mammalian embryonic development has been perceived as a migratory phenomenon, from the yolk sac blood island to the aorta-gonad-mesonephros (AGM) region, fetal liver (FL), and subsequently, the fetal bone marrow. In this study, we investigated the effects of primary stromal cells from fetal hematopoietic niches and their conditioned media (CM), applied singly or in sequential orders, on induction of human embryonic stem cells, H1, H9, and H14 lines, to hematopoietic cells. Our results demonstrated that stromal support of FL, AGM + FL, and AGM + FL + fetal bone marrow significantly increased the proliferation of embryoid bodies (EB) at day 18 of hematopoietic induction in the presence of thrombopoietin, stem cell factor, and Flt-3 ligand. AGM + FL also increased hematopoietic colony-forming unit (CFU) formation. CM did not enhance EB proliferation but CM of FL and AGM + FL significantly increased the density of total CFU and early erythroid (burst-forming unit) progenitors. Increased commitment to the hematopoietic lineage was demonstrated by enhanced expressions of CD45, alpha-, beta-, and gamma-globins in CFU at day 32, compared with EB at day 18. CM of FL significantly increased these globin expressions, indicating enhanced switches from embryonic to fetal and adult erythropoiesis. Over 50% and 10% of cells derived from CFU expressed CD45 and beta-globin proteins, respectively. Expressions of hematopoietic regulatory genes (Bmi-1, β-Catenin, Hox B4, GATA-1) were increased in EB or CFU cultures supported by FL or sequential CM. Our study has provided a strategy for derivation of hematopoietic cells from embryonic stem cells under the influence of primary hematopoietic niches and CM, particularly the FL.
Introduction
D
To date, it has proven difficult to obtain sustainable engraftment of ESC-derived hematopoietic stem/progenitor cells in the mouse transplantation model. The failure of persistent hematopoiesis is believed to reflect the developmental immaturity of these cells [2 –4]. It is critical that ESC-derived hematopoietic stem/progenitor cells are capable of sustaining their normal functioning of self-renewal and differentiation, if applied for transplantation purpose. The migration of hematopoietic development has been a tightly regulated program conserved in mammalian development. It has been increasingly emphasized that the microenvironment, which provides cell-to-cell contact, the extracellular matrix, as well as secretory growth and differentiation factors, plays a key role in determining hematopoietic stem cell functions [5,6]. It is thus rational to speculate that such microenvironments and possibly the chronological shift of developmental sites are essential for normal derivation of definitive hematopoietic stem cells from ESCs and the subsequent maturity of these cells to functional hematopoietic progenitors.
In this study, we proposed to evaluate the effects of fetal cellular microenvironments and their conditioned media (CM), applied singly or sequentially, on the derivation of hematopoietic cells from ESCs. The read-out parameters were embryoid body (EB) proliferation, colony-forming capacity, globin gene switch, and expression of candidate genes that are strongly implicated in the self-renewal of hematopoietic stem/progenitor cells during various developmental stages [7 –9]. Our results demonstrated promoting effects of FL on EB proliferation and globin switch from embryonic to fetal and adult phenotypes.
Materials and Methods
Work on animal and human tissues was approved by Institutional Committees on Animal Ethics and Human Ethics, respectively.
Maintenance of ESCs
The human ESC lines H1 (P58-70), H9 (P48-53), and H14 (P44-68) (Wicell) were maintained as previously described [10,11]. In brief, they were cocultured with irradiated mouse embryonic fibroblasts (ICR-CF1 strain) in Dulbecco's modified Eagle's medium/F12 supplemented with 20% Knockout Serum Replacer (Invitrogen Corporation), 1% nonessential amino acids (Invitrogen Corporation), 1 mM
EB formation and induction of hematopoietic differentiation on fetal stromal cell supports
Dissociated ESCs (5 × 104/mL) were cultured in low-adherent 24-well plates (Corning Costar Corporation). Induction of EB toward the hematopoietic lineage was performed in Stemline II medium (Sigma) containing defined cytokines: human recombinant bone morphogenic protein-4 (50 ng/mL; Peprotech), vascular endothelial cell growth factor (50 ng/mL; Peprotech), Flt-3 ligand (Flt-3L, 50 ng/mL; Peprotech), stem cell factor (SCF, 50 ng/mL; Peprotech), and thrombopoietin (TPO, 50 ng/mL; Peprotech), with or without stromal cell support.
Mouse AGM stromal cells were extracted from day 11.5 mouse embryos [5]. Human FL and FBM stromal cells were obtained from aborted fetuses (weeks 9–14) after legal therapeutic abortions, performed in compliance with the Legislation of the Hong Kong Government and Institutional Policy of the Prince of Wales Hospital, The Chinese University of Hong Kong, with written, informed consent. The collected cells were cultured in Dulbecco's modified Eagle's medium supplemented with 15% (v/v) fetal calf serum (FCS; Invitrogen Corporation). After 4–5 passages, the established stromal cells were stored as frozen stocks.
For EB cocultured on stromal cells, ESC lines H1, H9, and H14 were investigated. ESCs (5 × 104 per mL) were transferred onto γ-irradiated (80-Gy) stromal monolayer in Stemline II hematopoietic cell expansion medium and cytokines. The treatment groups were (1) Stemline II medium with cytokines TPO, SCF, and FL (TSF only); (2) TSF + AGM; (3) TSF + FL; (4) TSF + FBM; (5) TSF + AGM + FL; (6) TSF + AGM + FBM; and (7) TSF + AGM + FL + FBM. For single stromal support experiments [treatments (2), (3), and (4)], cells were cultured for a total of 18 days, with the EB transferred to a freshly irradiated stromal layer of the same cell type every 3 days. For sequential induction of EB by 2 stromal supports [ie, treatments (5) and (6)], the EBs were cocultured with the first stromal cell type for 9 days and then with the second stromal cell type for another 9 days. For EB induction with 3 stromal supports [ie, treatment (7)], the EBs were cocultured sequentially on each stromal cell type for 6 days. All induction experiments were performed for a total of 18 days, with the EB transferred to a freshly irradiated stromal layer (same or different cell type) every 3 days. EB transfer was also performed for the TSF-only control treatment (1), except that the cultures contained no stromal cells. Half medium was changed daily for all treatment groups.
Induction of hematopoietic differentiation by fetal stromal CM
Serum-free CM was prepared using primary mouse AGM, human FL, or human FBM stromal cells. Briefly, the cells were cultured at 2 × 105 per mL IMDM supplemented with 10% FCS, 1% nonessential amino acids, 1 mM
To induce hematopoietic EBs in CM, ESC lines H1 and H14 were investigated. ESCs were cultured in 50% CM harvested from AGM, FL, or FBM, supplemented with TSF. The treatment groups were (1) TSF only; (2) TSF + FL; (3) TSF + AGM + FL; and (4) TSF + AGM + FL + FBM. The culture time schedules of single or sequential CM support were same as those described for cultures with stromal cells. Half medium was changed daily. At day 18, the differentiated EB cells were dissociated into single cells using collagenase IV for 30 min, followed by cell dissociation buffer for 20 min (Sigma).
Hematopoietic differentiation of day 18 EBs was assessed by further culture of multilineage hematopoietic progenitor cells by colony-forming unit (CFU) assays on semisolid methylcellulose for 14 days (ie, a total of 32 days from seeding of ES cells). Quantitative analyses of expressions of alpha-, beta-, epsilon-, gamma-, and zeta-globin genes, as well as hematopoietic regulatory genes were performed on day 18 and 32 cells by quantitative polymerase chain reaction (qPCR). The proportion of cells bearing specific hematopoietic protein markers (CD34, CD45, beta-globin) were determined by flow cytometry.
CFU assay
Dissociated EB cells (2 × 105) at day 18 were cultured in semisolid Methocult GF medium (StemCell Technologies) consisting of 1% methylcellulose, 30% FCS, 1% bovine serum albumin, 50 ng/mL SCF, 20 ng/mL granulocyte–macrophage colony-stimulating factor, 20 ng/mL interleukin-3, 20 ng/mL interleukin-6, 20 ng/mL granulocyte colony-stimulating factor, and 3 units/mL erythropoietin. After 14 days, the plates were scored for mixed (CFU-GEMM), granulocyte–macrophage (CFU-GM), monocyte (CFU-M), and erythrocyte [burst-forming unit-E (BFU-E) and CFU-E] lineages according to standard criteria [12].
Reverse transcription and qPCR
Total RNA was extracted from EB (day 18) and CFU (day 32) cells using Trizol reagent (Invitrogen Corporation). Reverse transcription was carried out using High-Capacity cDNA Reverse Transcription Kit with RNase Inhibitor and TaqMan® Universal PCR Master Mix (Applied Biosystems) according to the manufacturer's instructions. qPCR analysis was carried out using human-specific TaqMan Gene Expression Assays (Applied Biosystems) as shown in Supplementary Table S1 (available online at
Flow cytometry analysis
For detection of CD34 and CD45 expressions at days 18 and 32, washed cells were incubated with antibodies against human CD34 and CD45 antigens: CD34-PE and CD45-FITC (Becton-Dickinson Biosciences), and their corresponding IgG1 isotype controls. Live cells were identified by 7AAD (Becton-Dickinson Via-probe™) exclusion and were analyzed for cell surface antigen expression with the Cell-Quest Pro Software (Becton-Dickinson). For detection of beta-globin protein expression, the cells were fixed and permeabilized using the Cytofix/Cytoperm™ Kit (Becton-Dickinson). Permeabilized cells were stained with mouse anti-human hemoglobin-beta-FITC antibody (Santa Cruz Biotechnology), 4'-6-diamidino-2-phenylindole (Abbott Molecular), and corresponding IgG1 isotype controls. As a positive control, cord blood mononucleated cells were stained with hemoglobin-beta-FITC, CD71-PE, and CD45-PC5, and the expression level of beta-globin in CD45-negative CD71-positive cells was determined (Supplementary Fig. S1, available online at
Statistical analysis
In pilot experiments, we have optimized the duration of EB culture and cytokine combination using TPO, SCF, and Flt-3 ligand (TSF) for induction of hematopoietic progenitor cells from human ESC lines H1, H9, and H14 (Supplementary Figs. 2 and 3, available online at
Results
Fetal stromal cells promoted EB proliferation and induction of CFU
In TSF-containing medium, ESC cultures with single FL stromal support and AGM + FL or AGM + FL + FBM sequential support significantly promoted cell proliferation at 18 days of EB formation, compared with those containing TSF only (*P = 0.012, **P < 0.01; Fig. 1). AGM had no effect on cell proliferation, whereas AGM + FBM significantly reduced cell number (# P = 0.028). Compared with TSF-only cultures, FBM showed a trend of reduction in cell proliferation [fold expansion: FBM (N = 3) vs. TSF only (N = 3); 63.2 ± 15.0 vs. 91.1 ± 3.46, P = 0.155].
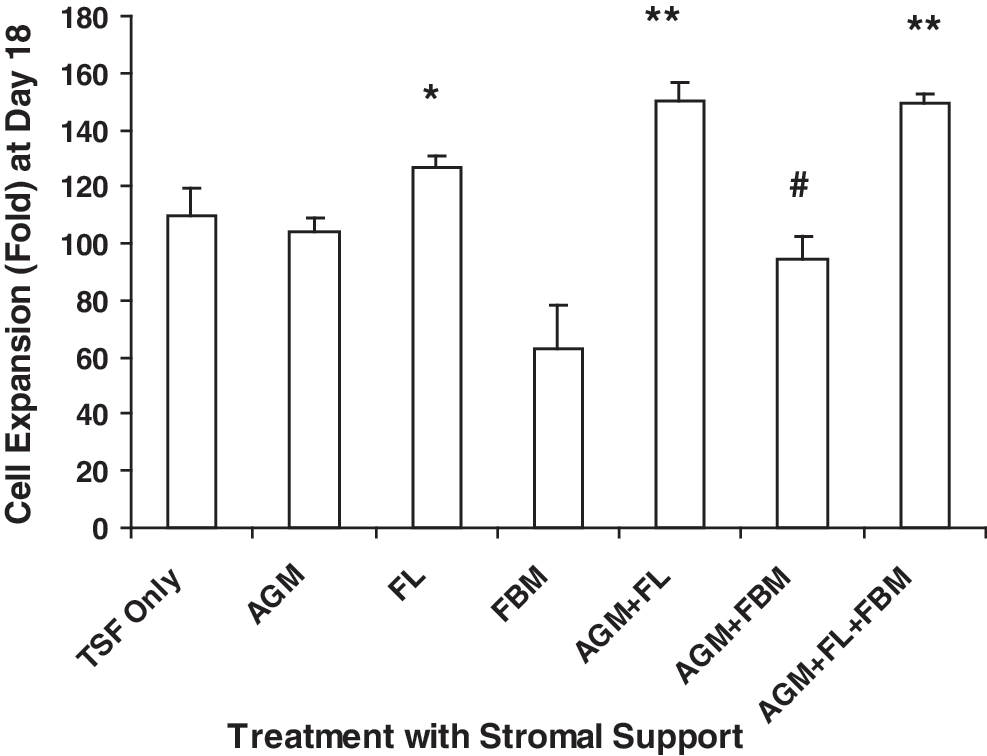
Effects of stromal support on cell proliferation at day 18 of EB culture. ESCs (5 × 104) were cultured for EB formation in the presence of TPO, SCF, and Flt-3 ligand (TSF, each 50 ng/mL) only, or with single stromal support of mouse AGM, human FL, or human FBM, or sequential stromal support with AGM + FL, AGM + FBM, or AGM + FL + FBM. Results are expressed as fold increase of cell number (mean ± SEM). Paired t-test analyses were performed on data from cultures supported by TSF + fetal stromal cells (N = 3–7), comparing with respective TSF-only cultures. FL, AGM + FL, or AGM + FL + FBM significantly increased cell expansion, whereas AGM + FBM decreased cell expansion. *P = 0.012; # P = 0.028; **P < 0.01. AGM, aorta-gonad-mesonephros; EB, embryoid body; ESC, embryonic stem cell; FL, fetal liver; FBM, fetal bone marrow; SCF, stem cell factor; SEM, standard error of the mean; TSF, thrombopoietin, SCF, and Flt-3 ligand; TPO, thrombopoietin.
Hematopoietic induction, in terms of multilineage CFU recovery from 5 × 104 ESCs seeded at day 0, was significantly affected by AGM + FL and AGM + FL + FBM sequential cocultures (Fig. 2). In AGM + FL cocultures, total CFU, CFU-GEMM, BFU-E, and CFU-E were significantly increased (*P < 0.05, **P < 0.01), compared with cultures containing TSF only without any stromal support. In AGM + FL + FBM cocultures, total CFU, CFU-GEMM, CFU-M, BFU-E, and CFU-E were significantly increased, compared with TSF-only cultures.
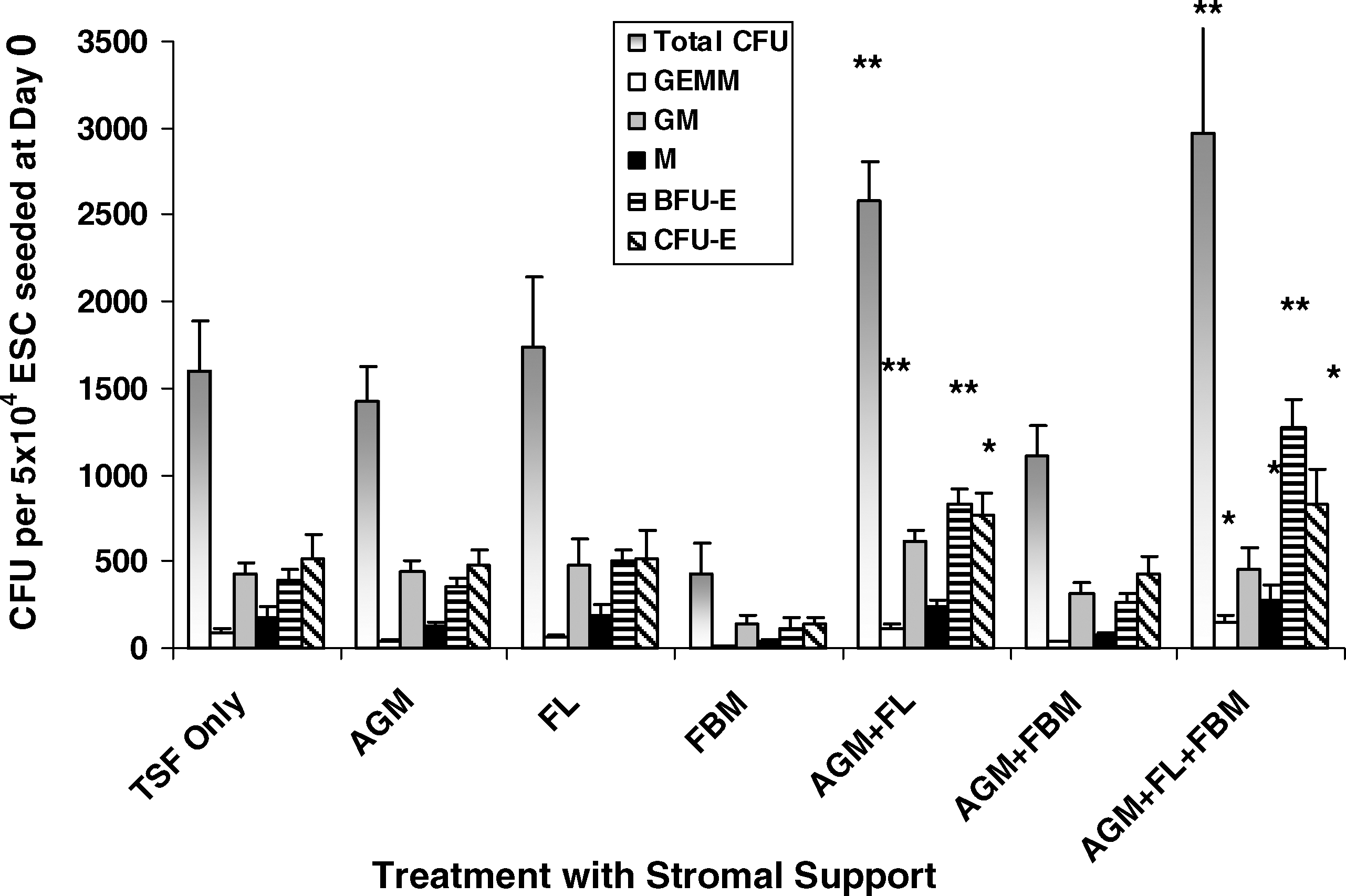
Effects of stromal support on hematopoietic progenitor CFU formation. ESCs (5 × 104) were cultured for EB formation in the presence of TSF (each 50 ng/mL) only, or with single stromal support of mouse AGM, human FL, or human FBM, or sequential stromal support with AGM + FL, AGM + FBM, or AGM + FL + FBM for 18 days. Differential CFU assay of the mixed (GEMM), myeloid (GM and M), and erythroid (BFU-E and CFU-E) lineages was then performed on Methocult semisolid culture for 14 days. Results are expressed as the number of CFU per 5 × 104 originally seeded cells (mean ± SEM). Statistical analyses were performed on data from cultures supported by TSF + fetal stromal cells, compared with those with TSF only. AGM + FL or AGM + FL + FBM significantly increased CFU formation. *P < 0.05; **P < 0.01; N = 3–7. BFU-E, burst-forming unit-erythrocyte; CFU, colony-forming unit.
Fetal CM promoted hematopoiesis
At day 18 of EB formation, exposure to fetal CM, single or sequential, did not promote cell proliferation (Fig. 3). Presence of 50% CM of AGM + FL slightly but significantly reduced cell proliferation from 64.9 ± 4.07-fold to 55.8 ± 3.71-fold (## P = 0.005, N = 9). The proportion of CD34+ cells (mean value range of treatment groups: 1.39%–2.01%, N = 7) was relatively low as indicated by flow cytometric assay, and their numbers were not affected by the presence of CM. Embryonic and fetal globin genes (alpha-, epsilon-, gamma-, and zeta-) were expressed, but adult beta-globin mRNA was barely detectable (Fig. 4A). All globin gene expressions were not significantly altered by various stromal CM in these day 18 cells.
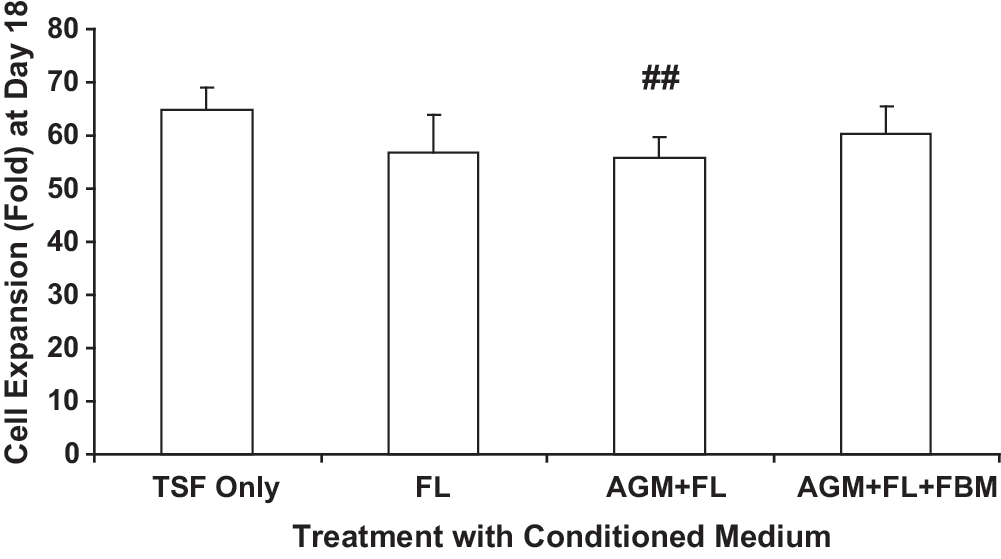
Effects of fetal stromal CM on cell proliferation at day 18 of EB culture. ESCs (5 × 104) were cultured for EB formation in Stemline II medium in the presence of TSF (each 50 ng/mL) only, or with additional 50% CM collected from FL, or sequentially with AGM + FL or AGM + FL + FBM. Results are expressed as fold increase (mean ± SEM) of cell number at day 18. Statistical analyses were performed on data from cultures supported by TSF + fetal stromal CM, compared with those with TSF only. AGM + FL significantly reduced cell expansion. ## P = 0.005; N = 9. CM, conditioned media.
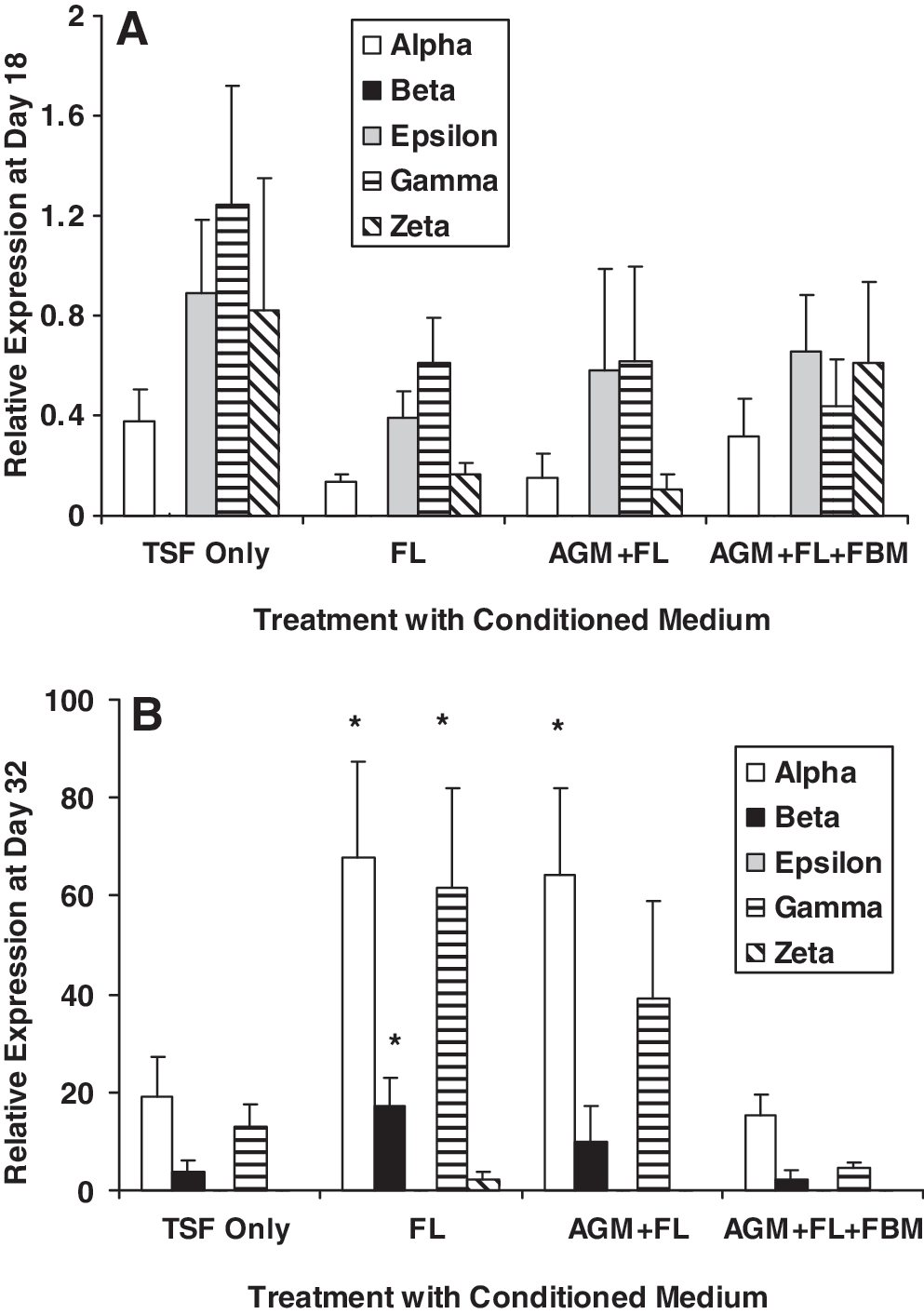
Effects of stromal CM on globin gene expressions. Globin gene expressions were analyzed by quantitative polymerase chain reaction using specific TaqMan probes at
After dissociation of EB at day 18, cells were subjected to a further 14 days of CFU culture (ie, a total of 32 days; Figs. 5 and 6E–6H). Treatment groups with FL or AGM + FL CM during EB culture had significantly higher density of total CFU, CFU-GEMM, and BFU-E in individual methycellulose plates (*P < 0.05; Fig. 5A). An increased proportion of BFU-E in the erythroid progenitor compartment (BFU-E + CFU-E) was also observed in FL-treated (P = 0.006) and AGM + FL-treated (P = 0.05) cultures, compared with TSF-only cultures. Wright-Giemsa staining of CFU cells showed multilineage hematopoietic progenitor cells of erythroid, myeloid, and megakaryocytic origins (Fig. 6I, J). We also observed a relative abundance of erythroid progenitor cells at different stages of maturation, displaying various extents of cytoplasm and coloration of hemoglobinization (Fig. 6K).
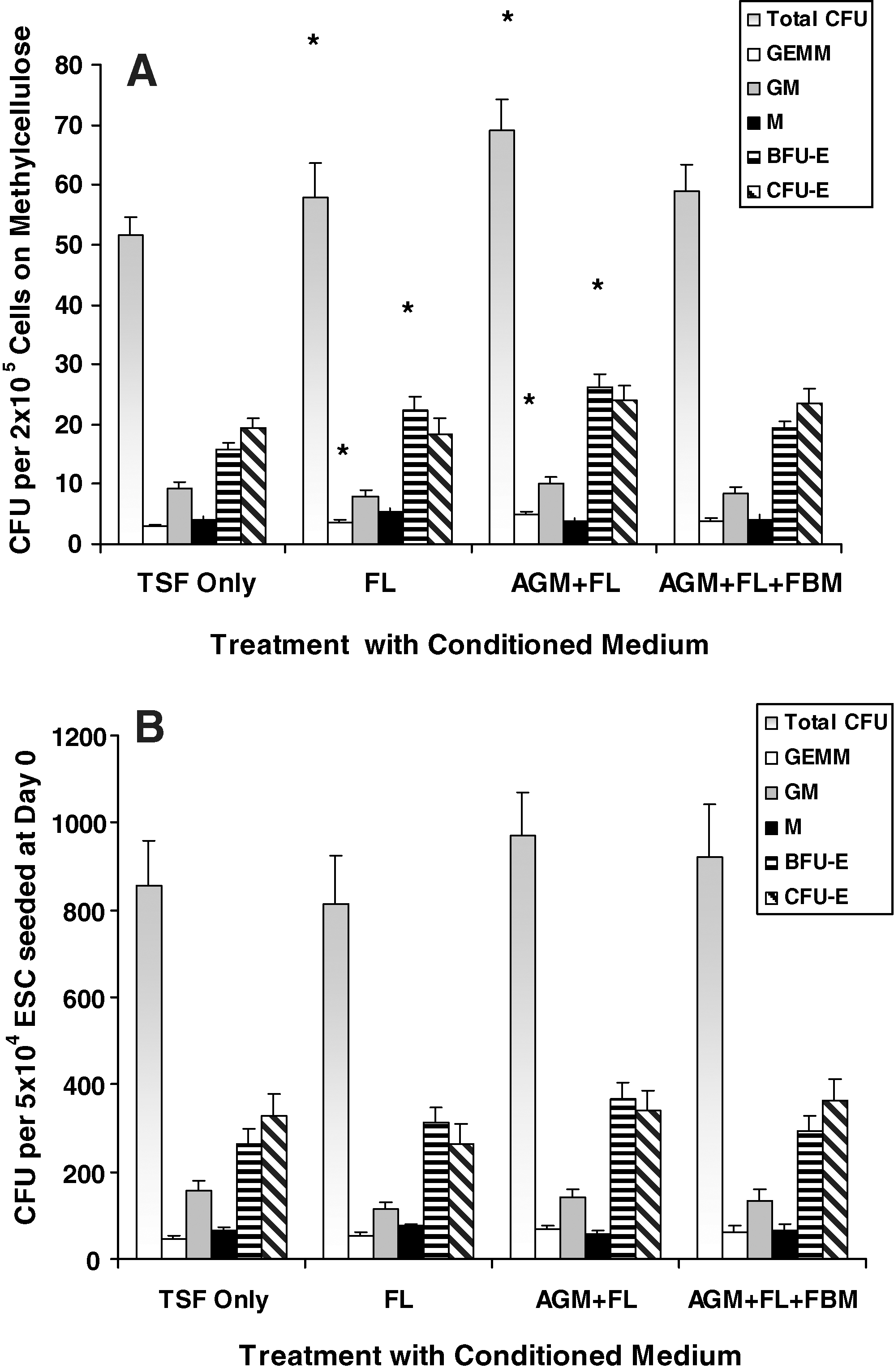
Effects of fetal stromal CM on hematopoietic progenitor CFU formation. ESCs (5 × 104) were cultured for EB formation in Stemline II medium in the presence of TSF (each 50 ng/mL) only, or with additional 50% CM collected from FL, or sequentially with AGM + FL or AGM + FL + FBM. Results are expressed as
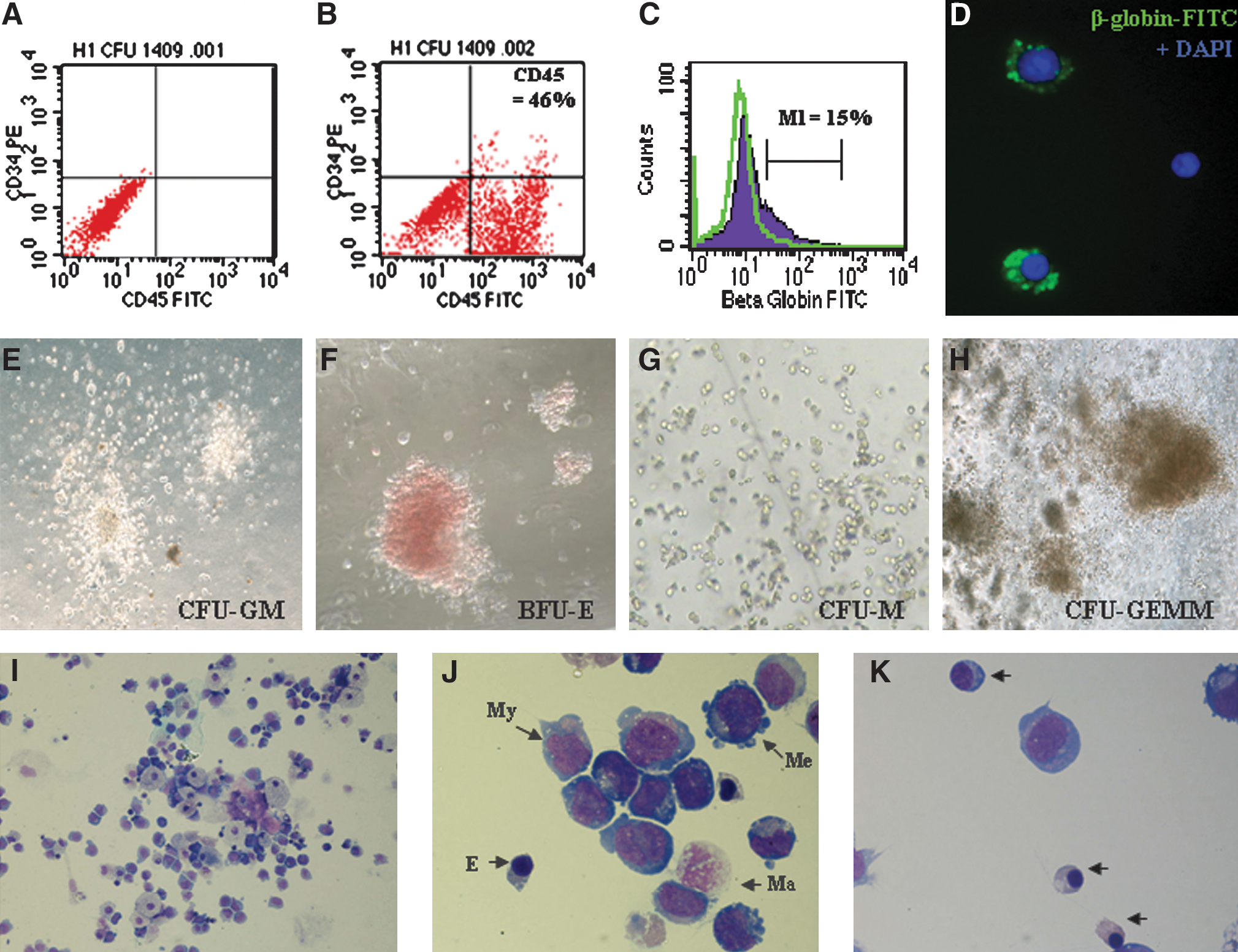
ESC-derived hematopoietic cells and expression of CD45 and beta-globin. ESC-derived HSCs at day 18 were subjected to CFU culture on Methocult semisolid medium. After 14 days, hematopoietic progenitors of the granulocyte–macrophage (CFU-GM), erythroid (BFU-E), monocytic (CFU-M), and mixed (CFU-GEMM) lineages were scored (
Taking into account the expansion efficiency (Fig. 3) of CM treatments, the CFU recovered from 5 × 104 ESCs initially seeded at day 0 were not different between groups (Fig. 5B). However, trends of increased CFU-GEMM (P = 0.07) and BFU-E (P = 0.09) in FL-treated cultures and increased CFU-GEMM (P = 0.10) and BFU-E (P = 0.07) in AGM + FL cultures were observed. When cells from CFU were subjected to flow cytometric analysis, we observed high percentages of CD45-expressing cells (range of mean values of treatment groups: 51.0%–53.8%, N = 7) and relatively low levels of CD34+ cells (<1%), indicating a high degree of hematopoietic maturation. Levels of CD45+ or CD34+ cells were not significantly affected by various CM treatments (Fig. 6A, B).
There were relatively high levels of globin mRNA expression in CFU cells (day 32; Fig. 4B) compared with day 18 cells (Fig. 4A), especially the mature (beta-) and fetal (alpha- and gamma-) globins, whereas embryonic globins (epsilon- and zeta-) were low (Fig. 4B). The expression ratios of alpha/zeta, gamma/epsilon, and beta/gamma globins were distinctively higher in day 32 CFU, compared with those of day 18 EB, indicative of globin-switch from embryonic to fetal and fetal to adult phenotypes (Supplementary Table S2, available online at
Fetal CM supported hematopoietic regulatory gene expressions
Hematopoietic regulatory genes were generally enhanced in the presence of stromal CM, especially after CFU culture (Fig. 7B). In day 18 EB culture (Fig. 7A), FL CM significantly enhanced the expressions of Bmi-1 and β-catenin (P < 0.05), whereas AGM + FL + FBM CM increased Hox B4 expression (P = 0.028). In CFU cultures (Fig. 7B), FL CM significantly increased GATA 1 expression (P = 0.047) and showed a trend of increased Bmi-1 (P = 0.098). Wnt-1 expressions were low in most cultures.
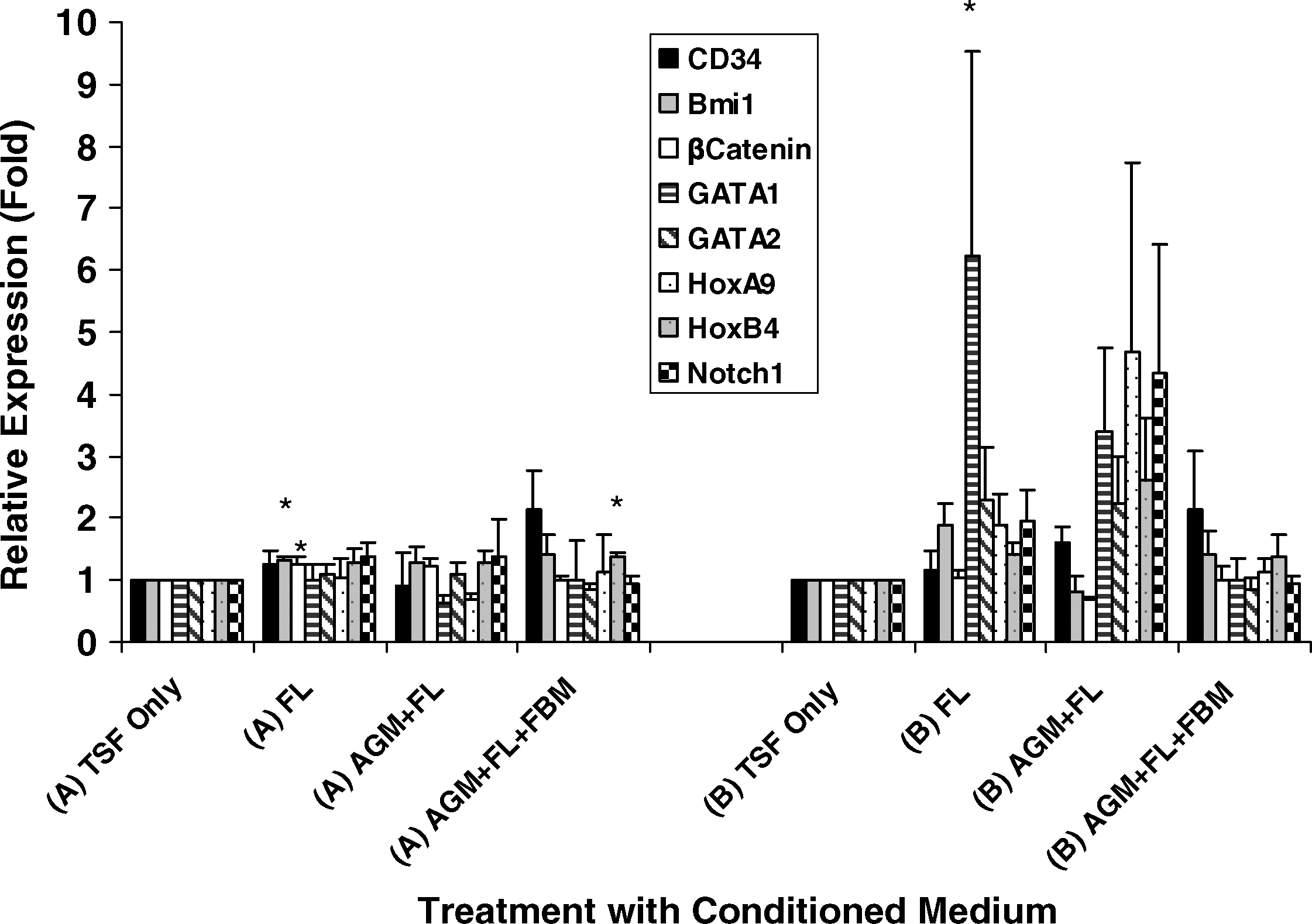
Effects of stromal CM on hematopoietic regulatory gene expressions. At
Discussion
Our study demonstrated that stromal cells and CM from specific fetal hematopoietic niches, particularly FL and AGM + FL, promoted induction of human ESCs to hematopoietic cells. We also reported increases of alpha-, beta-, and gamma-globin expressions in FL CM-treated cells, indicative of globin switches from embryonic to fetal and from fetal to adult erythropoiesis.
Single FL stromal support, or sequential AGM + FL or AGM + FL + FBM, significantly increased EB proliferation in hematopoietic cytokines (Fig. 1). Such effects were not observed upon culturing EB in CM of these fetal stromal cells (Fig. 3), indicating that the proliferation process might involve factors such as cell-to-cell contact or matrix support, in addition to secretary factors collected in the CM. Our results are in line with other studies that demonstrated efficient hematopoietic differentiation of murine or human ESCs on stromal cells derived from primary mouse AGM and FL cells or cell lines [13,14]. It was noted that Transwell cocultures or extracellular matrix of these stromal cells significantly reduced their hematopoietic promoting activities [13]. In their study, transforming growth factor-beta 1 (TGF-β1) and TGF-β3 enhanced hematopoietic induction from ESCs. Under our experimental conditions, however, addition of TGF-β1 did not affect the number of CFU in EB cultures with TSF (Supplementary Fig. S4, available online at
Hematopoiesis through early development is marked by switches of gene expression from embryonic to fetal (zeta- to alpha- and epsilon- to gamma-) and from fetal to adult (gamma- to beta-) globins [15,16]. In previous studies, the occurrence of adult erythropoiesis was rarely seen during in vitro induction of hematopoietic cells from ESCs. In the presence of growth factor support during hematopoietic cell induction, only embryonic and fetal globins, with little or no adult erythropoiesis, had been reported [3,17,18]. Using 35 days of coculture with a human FL hepatocyte cell line FH-B-hTERT, Qiu et al. demonstrated a maturation switch of embryonic to fetal globin in H1-derived hematopoietic cells, but beta-globin was expressed in a very low level [19]. They also observed enucleated erythrocytes at 35-day but not at 14-day cocultures. In the present study, the coculture duration for all treatments was 18 days, which probably was not sufficient for the appearance of enucleated erythrocytes. However, we could detect relatively high expressions of fetal and adult globins (alpha-, beta-, and gamma-; Fig. 4B) and low levels of embryonic globins (epsilon- and zeta-) in cells from CFU colonies, indicating the occurrence of significant globin switch. These expressions were increased in FL and AGM + FL cultures (Fig. 4B). The protein expression of beta-globin in a subpopulation (>10%) of CFU cells was further confirmed by flow cytometry (Fig. 6C, D). This observation suggested that primary FL cells or CM, rather than an immortalized cell line [19], might be required to provide signals and factors for bridging the maturation gap during in vitro induction of hematopoietic cells from ESCs. Our results also indicated that although the number of beta-globin protein–expressing cells was not increased by pretreatment with stromal CM for 18 days during EB proliferation, those cells in the FL CM cultures had a higher capacity of beta-globin mRNA expression (Fig. 4B). The increased cellular and expression outcomes at day 32 compared with those at day 18 are in agreement with the stepwise developmental model of hematopoietic cells from ESCs [4]. It is also of significance that the globin switch was enhanced in CM-supported cultures, indicating that the switch could be triggered by soluble factors.
We observed upregulation of Bmi 1, β-catenin, Hox B4, and GATA 1 gene expressions in day 18 EB or day 32 CFU cells under the influence of FL or sequential CM stimulation (Fig. 7). These genes and transcription factors have established activities for promoting self-renewal, proliferation, differentiation, and engraftment potential of hematopoietic stem/progenitor cells [4,8,20 –23]. It is of significance that GATA 1 has known functions for promoting maturation switch from gamma- to beta-globin [23,24]. Bacon et al. suggested that during fetal development, commitment to the adult versus the fetal program was determined at an early pre-BFU-E stage of differentiation and was facilitated by effector cells present in the respective erythropoietic organs (liver vs. bone marrow) [24]. Further, Hox B4 has been reported to promote the transition of primitive to definitive hematopoietic stem/progenitor cells with engraftment potentials [25].
Our data demonstrated that CM support from FL or AGM + FL resulted in similar outcomes that were superior to those from other culture conditions. The presence of FL appeared to be sufficient for producing the targeted read-outs, and priming with AGM CM might be redundant. Because of the difficulty of obtaining AGM cells from human fetuses, we opted for using the primary mouse AGM stromal support as the initial step of sequential induction. Thus, it remains to be resolved whether a sequential support from primary, all human fetal hematopoietic niches would produce a superior induction of ESCs to HSCs and adult erythropoiesis. In a pilot study, we observed that the inclusion of stromal cells from adult human BM in the sequential culture did not contribute to induction of hematopoietic cells from ESCs. However, it remains to be investigated whether FBM stromal cells from other developmental stages (eg, third trimester) would affect globin switch. Further, induction of functional and definitive hematopoietic stem/progenitor cells would need to be confirmed by transplantation assays in NOD/SCID animals. For clinical transplantation of ESC-derived hematopoietic cells, it is also crucial to eliminate residual undifferentiated ESCs so as to prevent the formation of teratomas [26,27]. Studies might also be performed to identify specific factors in the fetal niches that regulate induction of ESCs to hematopoietic stem/progenitor cells and maintain their self-renewal and differentiation capacity.
In conclusion, we have established a culture system for derivation of hematopoietic cells from ESCs under the support of primary hematopoietic niches and CM, particularly the FL. The efficient induction of globin switch from embryonic to fetal and adult erythropoiesis is a significant step forward for the provision of sustainable hematopoietic cells for clinical purposes.
Footnotes
Acknowledgments
This study was financially supported by the Li Ka Shing Institute of Health Sciences, The Chinese University of Hong Kong, and Earmarked Grant 4518/05M, Research Grant Council, The Government of the Hong Kong Special Administrative Region, Hong Kong.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.