
Abstract
Embryonic stem (ES) cells differentiate in vitro into all cell lineages. We previously found that the p38 mitogen activated kinase (p38MAPK) pathway controls the commitment of ES cells toward either cardiomyogenesis (p38 on) or neurogenesis (p38 off ). In this study, we show that p38α knock-out ES cells do not differentiate into cardiac, endothelial, smooth muscle, and skeletal muscle lineages. Reexpression of p38MAPK in these cells partially rescues their mesodermal differentiation defects and corrects the high level of spontaneous neurogenesis of knock-out cells. Wild-type ES cells were treated with a p38MAPK-specific inhibitor during the differentiation process. These experiments allowed us to identify 2 early independent successive p38MAPK functions in the formation of mesodermal lineages. Further, the first one correlates with the regulation of the expression of Brachyury, an essential mesodermal-specific transcription factor, by p38MAPK. In conclusion, by genetic and biochemical approaches, we demonstrate that p38MAPK activity is essential for the commitment of ES cell into cardiac, endothelial, smooth muscle, and skeletal muscle mesodermal lineages.
Introduction
E
Self-renewal of mouse ES cells is dependent on intracellular pathways initiated by the leukemia inhibitory factor (LIF), by either serum or bone morphogenetic protein 2 or 4, and by a complex interplay between specific epigenetic processes, miRNAs, and transcription factors involved in the development of the embryo, such as Oct4, Nanog, Sox2, or FoxD3 (for review, see [1]). Removing LIF and adding appropriate differentiation reagents result in the commitment of ES cells into a variety of mature differentiated cell types, including cardiac cells, skeletal muscle cells, neurons, or adipocytes (for review, see [2,3]). ES cell differentiation can be achieved by a wide variety of experimental protocols that lack or include fetal bovine serum (FBS) and the use of specific inducers such as the potent morphogen retinoic acid (RA). The 10−7 mol/L RA treatment between the second and fifth day is necessary for ES cell differentiation into neurons and adipocytes [4 –6]. In contrast, ES cell differentiation in the absence of RA yields efficient differentiation to cardiomyocytes with few neurons [4,7]. It is very likely that different differentiation protocols and inducers will selectively activate distinct signaling pathways that turn on cell lineage-specific genetic programs to bring about the observed enrichment in differentiated cell populations. Yet, the precise molecular process of these signaling pathways controlling cell differentiation of ES cells remains poorly understood.
p38 mitogen activated kinase (p38MAPK), extracellular signal-regulated kinase (ERK), and c-Jun NH2-terminal kinase (JNK) are conserved members of signal transduction pathways activated in response to growth factors or environmental stresses [8]. ERK appears to play a major role in cell proliferation and differentiation, whereas JNK is involved in apoptosis. The p38MAPK family includes 4 members (p38α, p38β, p38δ, and p38γ). These proteins are activated by phosphorylation in response to osmotic stress, UV, and various cytokines involved in inflammatory responses. p38MAPKs, mainly p38α, have been proposed to regulate several cellular processes, such as proliferation, cell survival, and differentiation [9]. Further, p38MAPK is directly involved in the differentiation and/or survival of several cell types, including neurons, endothelial cells, and myoblasts [10 –12]. However, these studies analyzed the role of p38MAPK in the terminal differentiation of established cell lines and primary cultures; therefore an early p38MAPK role in the formation of a given lineage cannot be evidence by these approaches. Deletion of the p38α gene leads to early embryonic lethality between 11.5 and 12.5 days due to erythropoiesis deficiency [13] and/or to abnormal placental development [14], demonstrating an essential role of p38MAPK in development. In this case, the early embryonic lethality impairs the unveiling of a potential role of this kinase pathway later on during development. Therefore, analysis of ES cell differentiation represents a unique experimental model to dissect p38MAPK functions from undifferentiated ES cells to well-differentiated cells.
In ES cells, p38α accounts for most of p38MAPK activity because p38α−/− ES cells present no remaining p38MAPK activity [15]. Interestingly, we previously found that, at the opposite of wild-type (wt) ES cells, this knock-out (KO) cell line spontaneously form neurons and are impaired in cardiomyocyte formation. We found similar results by studying the effects of the highly specific p38MAPK chemical inhibitor PD169316; demonstrating that p38MAPK activity constitutes an early switch in ES cell commitment into neurogenesis (p38 off ) versus cardiomyogenesis (p38 on) [16].
As this regulation takes place at the onset of the differentiation process, in this study, we investigated the role of p38MAPK in the ES cell commitment into other mesodermal lineages potentially important for cellular therapy, such as endothelial lineage, smooth muscle, and skeletal muscle. We found that p38MAPK is necessary early for these differentiations and that this requirement can be dissected into 2 independent successive steps.
Materials and Methods
Differentiation of ES cells
Mouse ES cells CGR8 [17] were grown on gelatin-coated plates, as well as the 15−/− ES cells [18] after adaptation on gelatin. 1−/− cells [18] were grown on MEF feeder cells treated with 10 μg/mL mitomycin C for 3 h. For embryoid body (EB) formation at day 0, 1 × 103 ES cells were aggregated in hanging drops (20 μL) without LIF, as described previously [17]. For cardiomyocyte, neuron, and myotube formation, EBs were cultivated in suspension until day 7 and then plated on gelatin-coated 6-well plate as already described [16, 19]. Cardiomyocytes (beating cells) and neurons (cells with neurites) were visible at day 12 and myotubes (spontaneous contractile polynucleated myotubes) at day 26. Quantification was given as the percentage of EBs with cardiomyocytes, neurons, or myotubes. As for the endothelial and smooth muscle differentiation, EBs were plated on gelatin-coated 6-well plates or on 5 mg/mL Matrigel-coated 24-well plates (BD Biosciences) at day 3 and the media were changed every 2 days until day 12. During EB differentiation, 10 μmol/L of the p38 inhibitor PD169316 (Sigma-Aldrich) was added to the culture media at different periods of time.
Western blot analysis
For p38MAPK protein analysis, EBs were lysed as previously described [17]. Samples (30 μg) were separated by SDS–polyacrylamide gel electrophoresis on a 10% gel (Invitrogen) and transferred onto polyvinylidene difluoride membranes (Millipore). Membranes were incubated with antibodies against either all p38MAPK isoforms, MAPKAP-K2, MAPKAPK2phospho (Cell Signalling Technology), ERK2, or HA epitope (Santa Cruz).
Immunofluorescence staining
Differentiated EBs were washed and fixed at day 7 in 3% paraformaldehyde for 15 min at room temperature. Cell membranes were permeabilized in phosphate-buffered saline (PBS) with 0.1% Triton X-100 for 15 min at room temperature and incubated with a monoclonal anti-CD31/PECAM (eBioscience), an anti-Troponin T (CT3), or an anti-MHC (MF2Ds) (DSHB) in PBS + 10% FBS for 1 h. After 3 washes in PBS, cells were incubated with anti-mouse ALEXA488 or ALEXA546 (Invitrogen) for 1 h at room temperature. Cell imaging was performed using a Nikon microscope Eclipse 90I. Z-stack pictures were obtained by using a Nikon Digital Sight DS-1QM camera controlled by NIS element AR software (Nikon).
Flow cytometry
EBs were dissociated into single-cell suspensions by trypsinization. Following a wash with 10% FBS/Dulbecco's modified Eagle's medium, cells were permeabilized with 0.2% Triton/PBS buffer for 10 min at room temperature. Cells were then stained with an α-cardiac Troponin T (CT3) antibody or an α-Myogenin (FD5) antibody at 1:10 dilution in 5% FBS/PBS (DSHB) for 1 h. Following washes with 5%FBS/PBS, cells were incubated with an anti-mouse IgG secondary antibody conjuguated to PE (1:100 dilution in 5% FBS/PBS; Beckman Coulter) for 30 min in the dark. After additional washes in PBS, cells were resuspended in 500 μL PBS and analyzed on an FC500 Flow cytometer using CXP software (Beckman Coulter). For each sample, at least 5,000 cells were analyzed. Cursor for positive fluorescent cells was placed, such as 95% of events were excluded (first decade) when an isotype IgG antibody was used. Results are given as percentage corresponding to positive fluorescent cells included in the region defined by the cursor.
RNA analysis
Total RNA was prepared using trizol reagent (Invitrogen). Real-time reverse transcription-polymerase chain reaction (RT-PCR) was performed with the ABI Prism7300 (Applied BioSystems) and Taqman or Mesa Green MasterMix (Eurogentec). The relative amounts of different mRNAs were quantified by using the comparative CT method (2−(ΔΔCT)). 36B4 was used as housekeeper transcripts and gene expressions were normalized using 36B4 RNA levels. Primer sequences are given in Supplementary Table S1 (Supplementary Data are available online at
Plasmids
Two sequences of chromatin insulator were added to the CAGop plasmid [20] using SacI restriction sites (pCAG-op-Ins2X). The pcDNA3-p38α vector containing the human p38α cDNA carrying an HA tag was a gift from Dr. Engelberg, Jerusalem, Israel. The pCAG-op-Ins2X-p38α was generated by inserting a KpnI-KpnI fragment from the pcDNA3-p38α into the pCAG-op-Ins2X.
Generation of ES cell lines
The 1−/− and 15−/− ES cell lines were cotransfected with the pGK-HygroR and pCAG-op-Ins2X-p38α, at a 10:1 ratio, using the FuGene HD transfection system (Roche Molecular Biochemicals) according to the manufacturer's protocol, and selected in 100 μg/mL hygromycin (Invitrogen) to generate p38−/− ES cell clones expressing exogenous p38α.
In the case of 1−/− cell line, transfection was realized onto mitomycin C-treated MEF feeder cells. Stable, hygromycin-resistant MEF cells were obtained by transduction with the CHC vector, an MLV retrovirus vector coding the hph cDNA [21] using an MOI of 6 UI/cell, followed by hygromycin selection (100 μg/mL). Moloney leukemia virus (MLV)-based vector was generated by transient transfection of 293T cells as previously described [22].
Statistical analyses
All experiments were performed at least 3 times. All data are expressed as the mean ± standard error of the mean. Treatments were compared with their respective controls and significant differences among the groups were determined using unpaired Student's t-test. A P-value of <0.05 was taken as an indication of statistical significance.
Results
p38α−/− ES cells do not differentiate into endothelial, smooth muscular, and skeletal muscular mesodermal lineages
In addition to cardiomyogenesis and neurogenesis, endothelial and smooth muscle differentiations are among the earliest lineages to arise, both in vivo, during embryo formation, and in vitro, during ES cell differentiation [23]. In the present study, we compared 2 p38α-homozygote KO ES cell lines [18], clones 1−/− and 15−/−, to the standard CGR8 ES cell line. Undifferentiated cells were induced to differentiate by the technique of EB formation [16] and RNAs were extracted by day 12 or 26 and gene expression for specific cell lineage markers was evaluated by real-time RT-PCR. Similar to what we previously found, spontaneous cardiomyogenesis was drastically impaired in these KO cells when compared with wt CGR8 cells. At day 12 of differentiation, cardiac phenotype, quantified by both the percentage of EB outgrowths with contractile cells and expression of the cardiac specific markers Nkx2.5 and GATA-4, was significantly reduced in the 2 KO cell lines when compared with wt cells (Supplementary Fig. 1A–C). In wt CGR8 cells, neuron formation by day 12 was not spontaneous and was strictly dependent on an RA treatment between days 3 and 5 [16]. In contrast, 35%–50% of EBs of the cell line 1−/− form neurons by day 12 of the differentiation protocol, with or without RA treatment, and express high levels of the neuronal specific marker MAP2, measured by real-time RT-PCR (Supplementary Fig. 2A, B and Fig. 5). Surprisingly, the 15−/− cell line does not show any neuronal differentiation tendency, even after RA treatment (Supplementary Fig. 2A, B). One could hypothesize that, in this cell line, neurogenesis is inhibited by an unknown molecular event, independent of KO of the p38α gene.
To test endothelial differentiation we used a previously established protocol [24]. Endothelial phenotype was characterized by anti-PECAM (also known as CD31) immunofluorescence (Fig. 1A, left column) and formation of typical vessel-like structures on Matrigel (Fig. 1A, right column). Endothelial differentiation was also analyzed by measuring the expression levels of endothelial-specific markers. mRNA expression profiles of Flk-1 and Tie2 genes show a sharp and robust increase after day 3 or 5 of differentiation, respectively, and an increasing Flk-1 expression until day 12, whereas Tie-2 expression diminishes after day 7 (Fig. 1B, C). The expressions of the endothelial markers PECAM and vWF show biphasic expression profiles, decreasing to low levels from day 0 to day 3, and then a progressive increase from day 3 to day 7 (Supplementary Fig. S1D, E). The PECAM profile is very similar to what has been already described in other ES cell lines [25]—the expression level found in undifferentiated cells was not correlated to endothelial differentiation. When induced to differentiate on either gelatin or Matrigel from day 3, p38−/− cells show almost no vessel-like structures (Fig. 1A); this lack of differentiation was characterized by the blockade of gene expression of the Flk-1, Tie-2, vWF, and CD31 markers (Fig. 1B, C and Supplementary Fig. S1D, E). These results present a discrepancy with those published by Guo et al., who showed that p38α KO ES cells can differentiate toward endothelial cells [26]. However, in this study, quantification of expression of endothelial markers was performed in a different KO ES cell line and, more importantly, only 2 markers were tested, one (PECAM) presenting no variation between KO and wt cells, whereas the other (VEcad) was found significantly downregulated in KO cells, suggesting an endothelial defect of these cells.
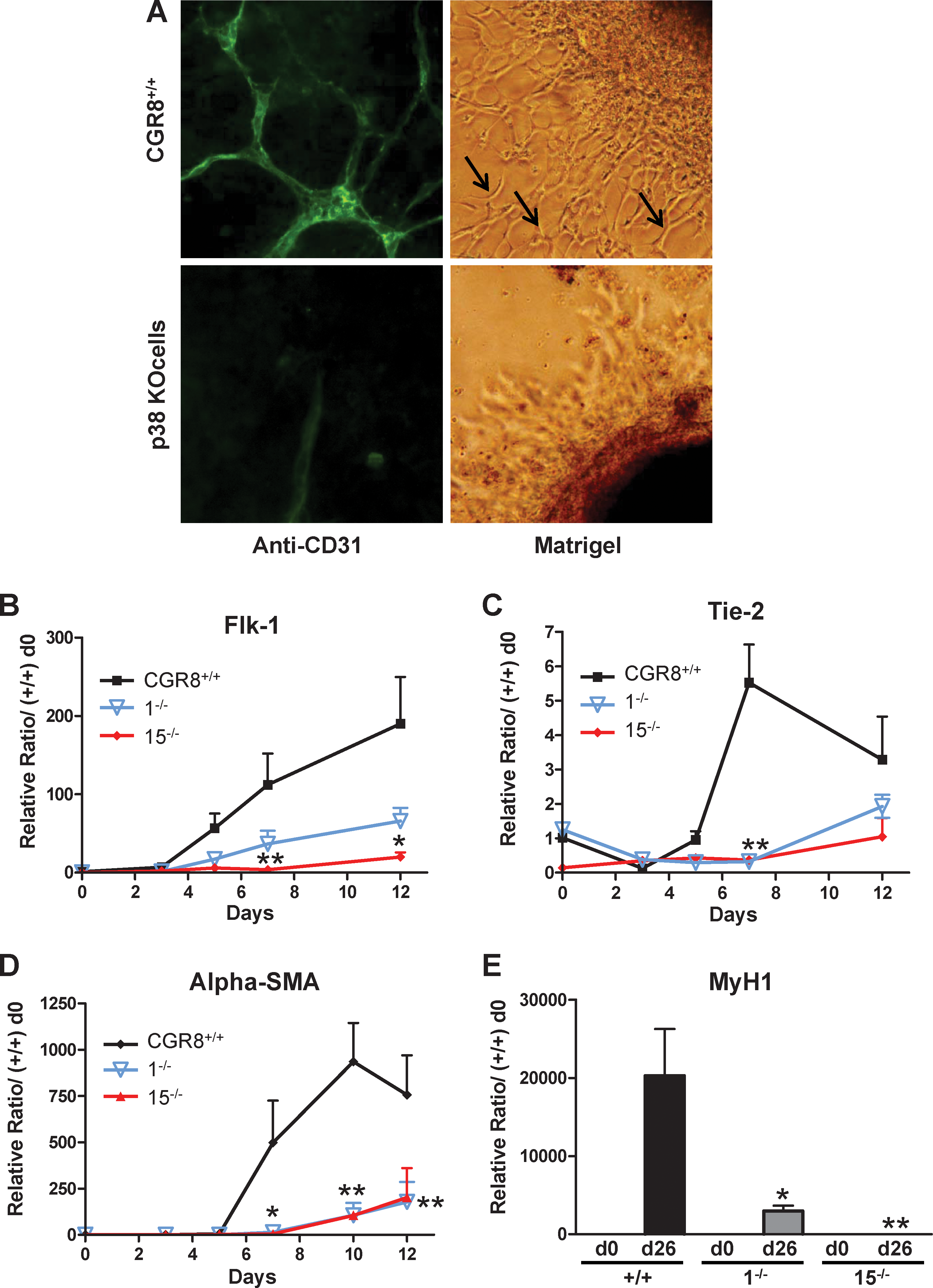
Wild-type but not p38α−/− ES cells differentiate toward endothelial cells, smooth muscle cells, or skeletal myotubes.
In parallel to endothelial cells, these culture conditions lead also, in the same Petri dishes, to the differentiation of vascular smooth muscle cells, which participate in the formation of vessel-like structures [24,27,28]. Our culture conditions are also permissive for this differentiation, as indicated by the high level of gene expression in wt ES cells at days 7–12 of the specific marker α-smooth muscle actin (Fig. 1D). Similar to endothelial differentiation, formation of the smooth muscle cell lineage was also impaired in p38α KO cells (Fig. 1D).
Our results suggest that, in addition to cardiomyogenesis, the p38MAPK pathway is required for the formation of endothelial and smooth muscle cell lineages.
We then analyzed the potential role of p38MAPK in mesodermal lineages formed later on during development, such as skeletal myogenesis. After 26 days of induction, wt ES cells form spontaneous contractile myotubes (Supplementary Fig. S1F) and express specific skeletal muscular genes such as Myosin Heavy Chain 1 (MyH1; Fig. 1E) and Myogenin (Supplementary Fig. S1G). In contrast, both the 2 p38α KO cell lines were impaired in their skeletal myogenic capacities, reduced number of EB-forming myotubes and reduced expression of markers (Fig. 1E and Supplementary Fig. S1F, G). These results demonstrate that the p38MAPK pathway is also required for formation of skeletal muscle, a late-forming mesodermal lineage.
Construction of p38α KO cell derivatives expressing exogenous p38 proteins
To demonstrate that mesodermal differentiation defects of p38α−/− cell lines are due to the KO of p38, we reexpressed p38MAPK in these KO cells. We used an expression vector for an HA-tagged p38α cDNA under the control of the synthetic CAG promoter, known to be very potent in ES cells before and upon differentiation [20,29]. We isolated and analyzed p38-reexpressing ES derivatives from KO cell lines: clone R1-1 for the cell line 1−/− and R15-1 and R15-2 for the cell line 15−/−. In the presence of LIF, all these cell lines display an ES morphology (data not shown) and express levels of Oct4 and Nanog, major specific genes of pluripotency, similar to wt and p38α KO ES cells (Supplementary Fig. S3A, B). We first checked the persistence of transgene expression during the differentiation process by real-time RT-PCR. As shown in Supplementary Fig. S3C, all cell lines conserve their p38 cDNA transgene expression from day 0 to day 26. Protein expression levels were analyzed by western blots with either anti-p38 total (α-p38tot) or anti-HA antibodies and anti-ERK antibodies as control (Fig. 2). Reexpressing cell lines present only partial restoration of wt endogenous p38 protein levels, between 23% and 46% of levels present in wt cells, at day 4 of differentiation (Fig. 2); similar results were obtained at days 0, 12 (data not shown), and 20 (Supplementary Fig. S3D). As a functional test for investigating the restored activity of the p38MAPK pathway, we analyzed the phosphorylation status of a known direct p38MAPK target: the MAPK activated protein kinase-2 (MAPKAP-K2) [9]. Western blots using anti-MAPKAP-K2 total or phosphorylated proteins demonstrated that p38MAPK KO cells display the remaining proportion of phosphorylated MAPKAP-K2 proteins comprised between 16% and 21% of levels in wt cells (Fig. 2). These low but not null levels indicate that, although the phosphorylation of MAPKAP-K2 protein is a good indication of α-p38MAPK activity, this protein can be targeted by other kinases. We found that incubation of wt cells with the p38MAPK inhibitor leads to only 50% reduction in MAPKAP-K2 phosphorylation (Fig. 2); this result could also be due to alternative pathways phosphorylating this protein. Interestingly, reexpressing cell lines present intermediate levels (32%–61%) between KO and wt, compatible with their partial restoration of p38MAPK expression (Fig. 2).
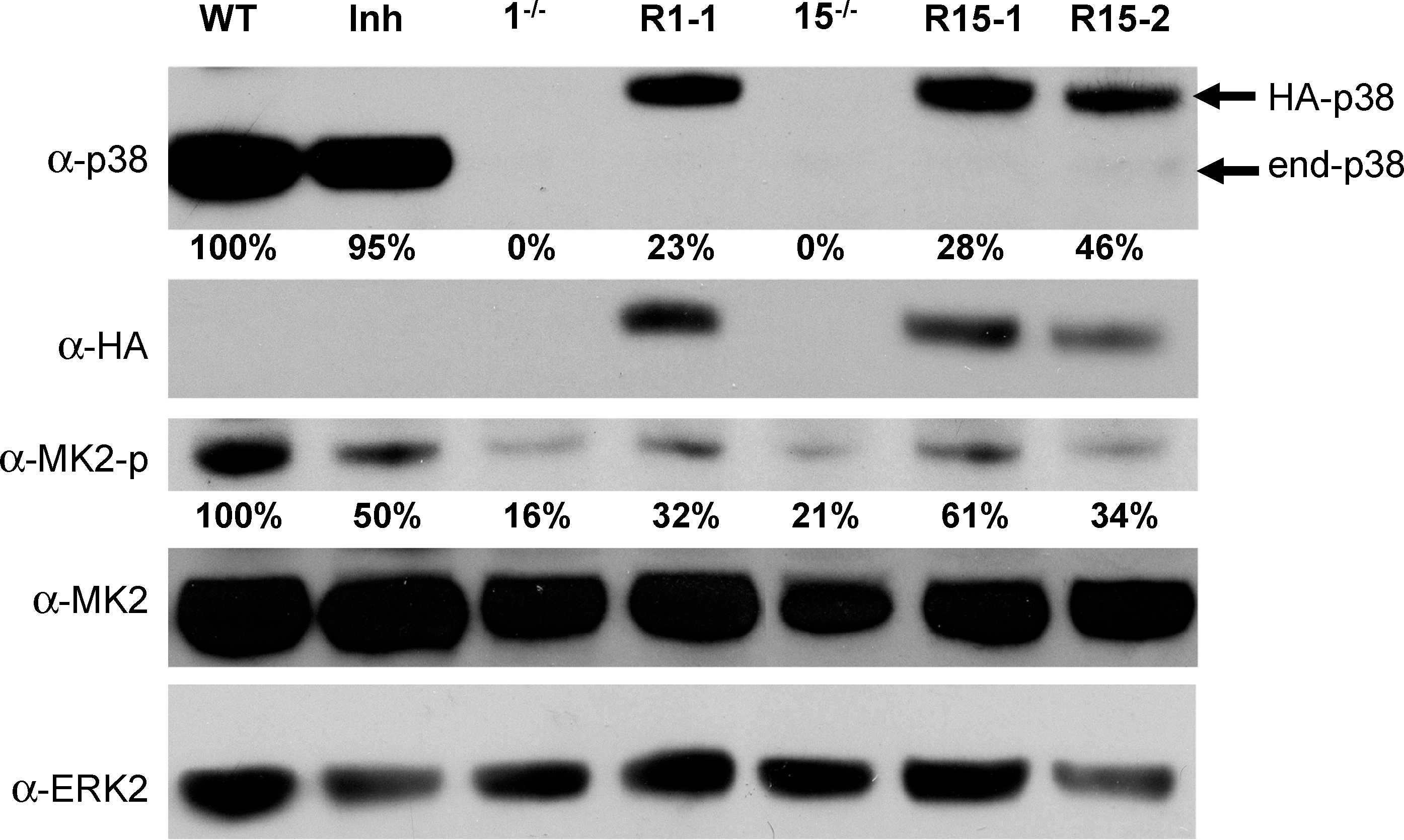
Molecular characterization of p38α KO cell derivatives partially reexpressing p38α protein. p38α protein expression at day 4 of differentiation of various ES cell lines was analyzed by western blotting with pan-p38 (α-p38), α-HA, α-phosphoMAPKAP-K2 (α-MK2-p), and α-MAPKAP-K2 total (α-MK2) antibodies. Membranes were reprobed with α-ERK2 antibodies as loading control. Migrations of endogenous p38 and HA-p38 proteins are indicated (right-hand side). Analysis of wild-type CGR8 cells (WT), treated (Inh) or not with the p38 specific inhibitor, the 1−/− and 15−/− p38 KO cell lines, and their derivatives R1-1 and R15-1 and −2 is shown. p38 protein expression signals obtained with pan-p38 antibodies were quantified, corrected for equivalent ERK expression, and expressed as percentage of the endogenous p38 expression found in CGR8 wt cells. PhosphoMAPKAP-K2-protein expression signals were quantified, corrected for equivalent total MAPKAP-K2 expression, and expressed as percentage of the expression found in CGR8 wt cells. α-HA, α-hemaglutinin antigen; MAPKAP-K2, MAPK, mitogen activated protein kinase-activated protein kinase-2; α-ERK2, α-extracellular signal-regulated kinase 2.
Reexpression of p38 protein in p38α KO cell lines rescues their defects in mesodermal differentiation
We then analyzed the reexpressing cell lines for their differentiation capacities toward the cardiac, endothelial, smooth muscular, skeletal muscular, and neuronal lineages.
Cardiomyogenesis was evaluated by the percentage of EBs with beating cells (Fig. 3A), Gata-4 (Fig. 3B) and Nkx2.5 expressions (Supplementary Fig. S4A), and flow cytometry with anti-Troponin T antibodies (Fig. 3C and Supplementary Fig. S4C) at day 12. Remarkably, cardiomyocytes formation was significantly, but partially, restored in the 3 reexpressing clones. This restored cardiomyogenic phenotype was also illustrated by immunofluorescence analysis with anti-Troponin T antibodies (Fig. 3D and Supplementary Fig. S4D).
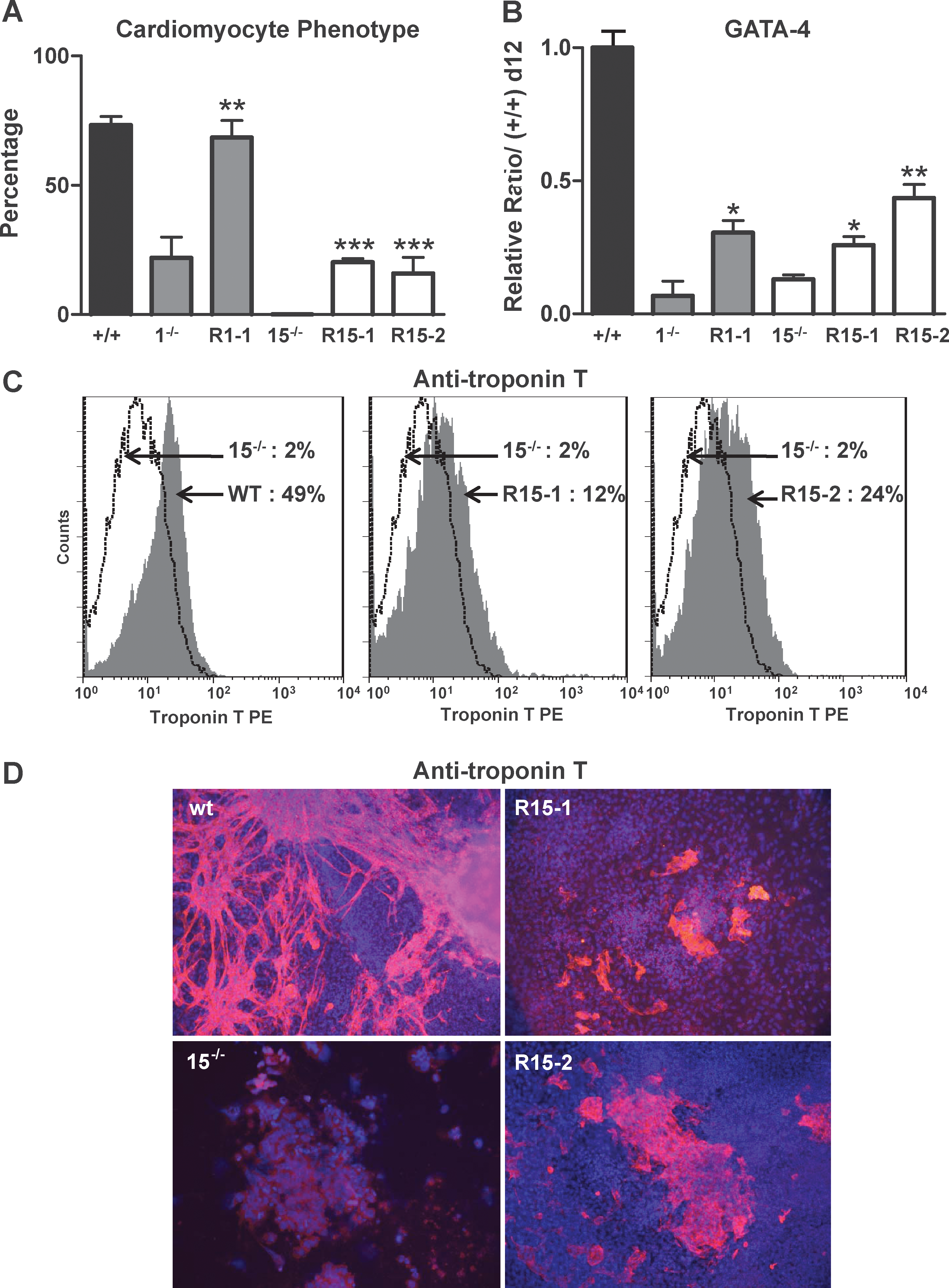
Reexpression of p38 protein in KO cell lines partially rescues their defects in cardiomyogenesis. EBs from wt CGR8, 1−/−, R1-1, 15−/−, R15-1, and R15-2 ES cells were plated on gelatin at day 7 of differentiation and analyzed at day 12.
Likewise, analysis of Tie-2, Flk-1, and alpha-smooth muscle actin expression levels show that endothelial and smooth muscle lineages are also partially, but significantly, restored in reexpressing clones (Fig. 4A, B and Supplementary Fig. S4B).
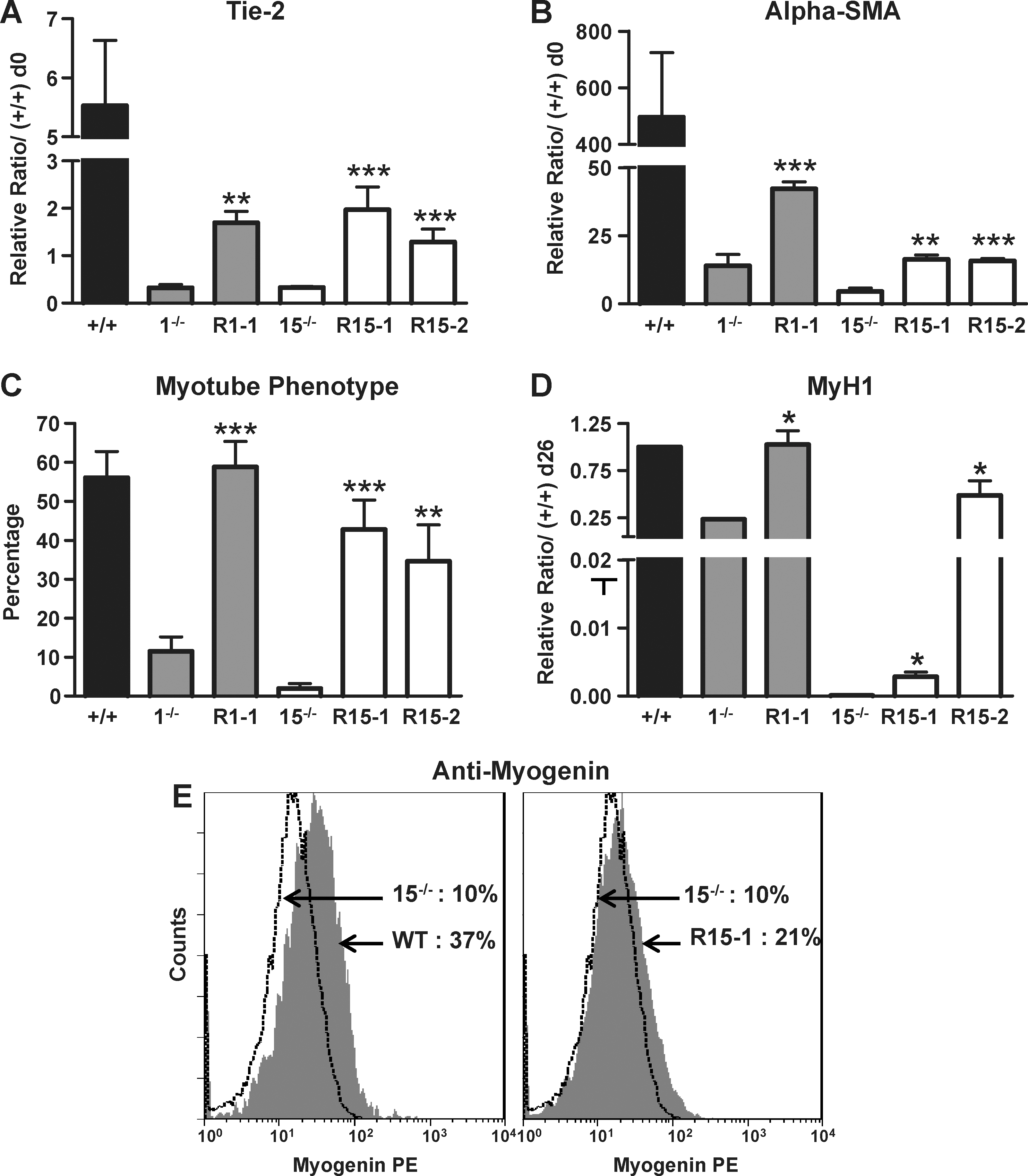
Reexpression of p38 protein in KO cell lines partially rescues their endothelial, smooth muscle, and skeletal muscle cell defects. EBs from wt CGR8, 1−/−, R1-1, 15−/−, R15-1, and R15-2 ES cells were plated on gelatin at day 3 for endothelial or smooth muscle cell differentiation or at day 7 for skeletal myogenesis and analyzed at day 7 for endothelial and smooth muscle cells or at day 26 for skeletal myogenesis. mRNAs at day 7 were analyzed by real-time RT-PCR for expression of the endothelial marker Tie-2
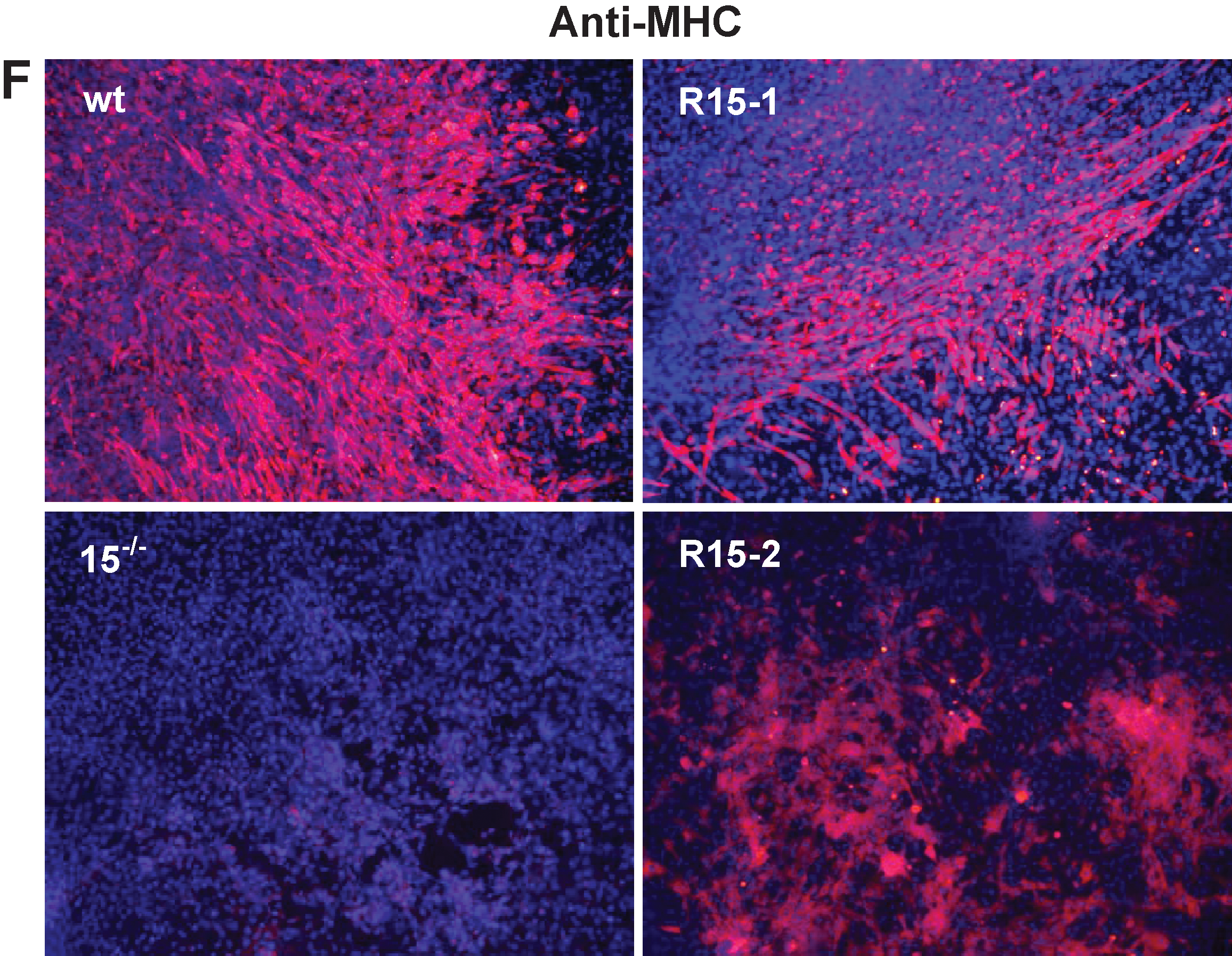
Skeletal myogenesis was evaluated by measuring the percentage of EBs with contractile myotubes, by Myogenin and MyH1 expressions, and by flow cytometry with anti-Myogenin antibodies at day 26. Strikingly, the clone R1-1 displays differentiation capacities completely restored to wt levels (Fig. 4C, D and Supplementary Fig. S5). The 2 other reexpressing clones, R15-1 and −2, present partial restored skeletal myogenic phenotypes (Fig. 4C–E and Supplementary Fig. S5A). This restored skeletal myogenic phenotype was also illustrated by immunofluorescence analysis with anti-MHC antibodies (Fig. 4F and Supplementary Fig. S5C).
Finally, neuronal differentiation, measured by the percentage of EBs with neuron outgrowths and MAP2 expression level, was analyzed in these cells. We found that the spontaneous neuron formation and the high level of MAP-2 expression characteristic of 1−/− cells are completely inhibited in R1-1 cells, back to wt levels, thereby restoring the p38 inhibitory effect toward this lineage (Fig. 5). Already being very low in 15−/− cells, neurogenesis was not modified in R15-1 and −2 derivatives (Supplementary Fig. S2).
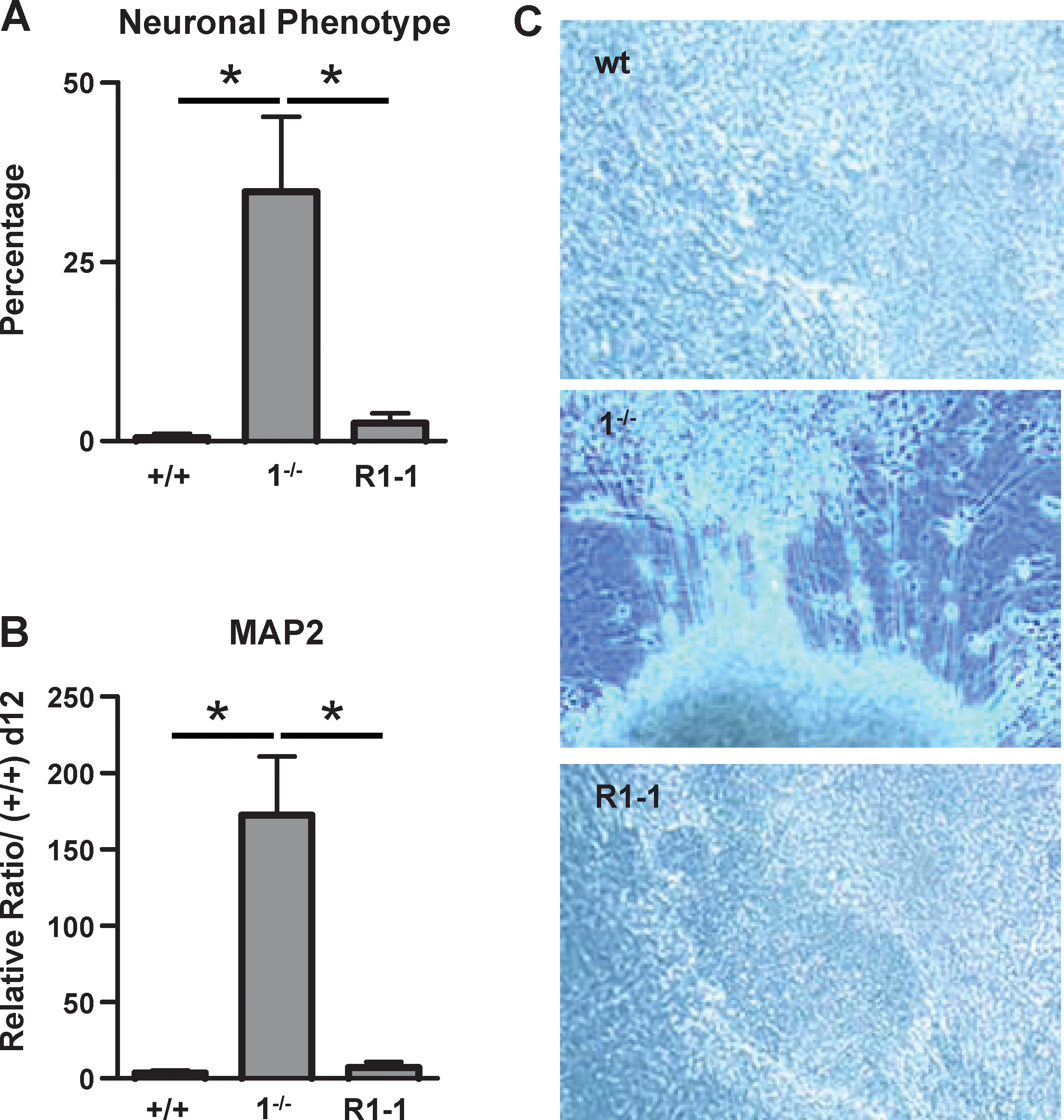
Reexpression of p38 protein in KO cell lines inhibits their spontaneous neurogenesis. EBs from wt CGR8, 1−/−, and R1-1 ES cells were plated on gelatin at day 7 and analyzed at day 12.
Altogether our data show that, despite the only partial reexpression of wt p38MAPK protein level, the p38MAPK KO-derivative clones are profoundly modified in their differentiation capacities. p38MAPK reexpression in KO cells restores, to various degrees, all wt mesodermal and neuronal phenotypes.
Taking into account the analysis of neurogenesis, our results suggest that, in addition to the p38α KO, variable molecular characteristics could participate in the differentiation defects of KO cell lines. One can hypothesize that these differences could be due to genetic instability of KO cells leading to variability in their biological responses.
p38MAPK activity is required for 2 early successive steps in mesodermal commitment of ES cells
To bypass these potential genetic differences, we analyzed the role of p38MAPK by directly inhibiting its activity in differentiating wt ES cells with the specific chemical inhibitor PD169316. Further, inhibitor treatment of differentiating ES cells from day 3 to day 5 blocks the spontaneous peak of p38MAPK activity observed at that step [16]. The inhibitor was applied in 3 different conditions: (i) all the time of differentiation, from day 0 to day 7, 12, or 26 (d0–d7, d0–12, d0–d26, depending of the lineage analyzed); (ii) only from day 0 to day 3 (d0–d3); (iii) from day 3 to day 5 (d3–d5) (Fig. 6A). We focused our study on the d0–d5 period because inhibition of p38MAPK after day 5 has no effect (Supplementary Fig. S6).
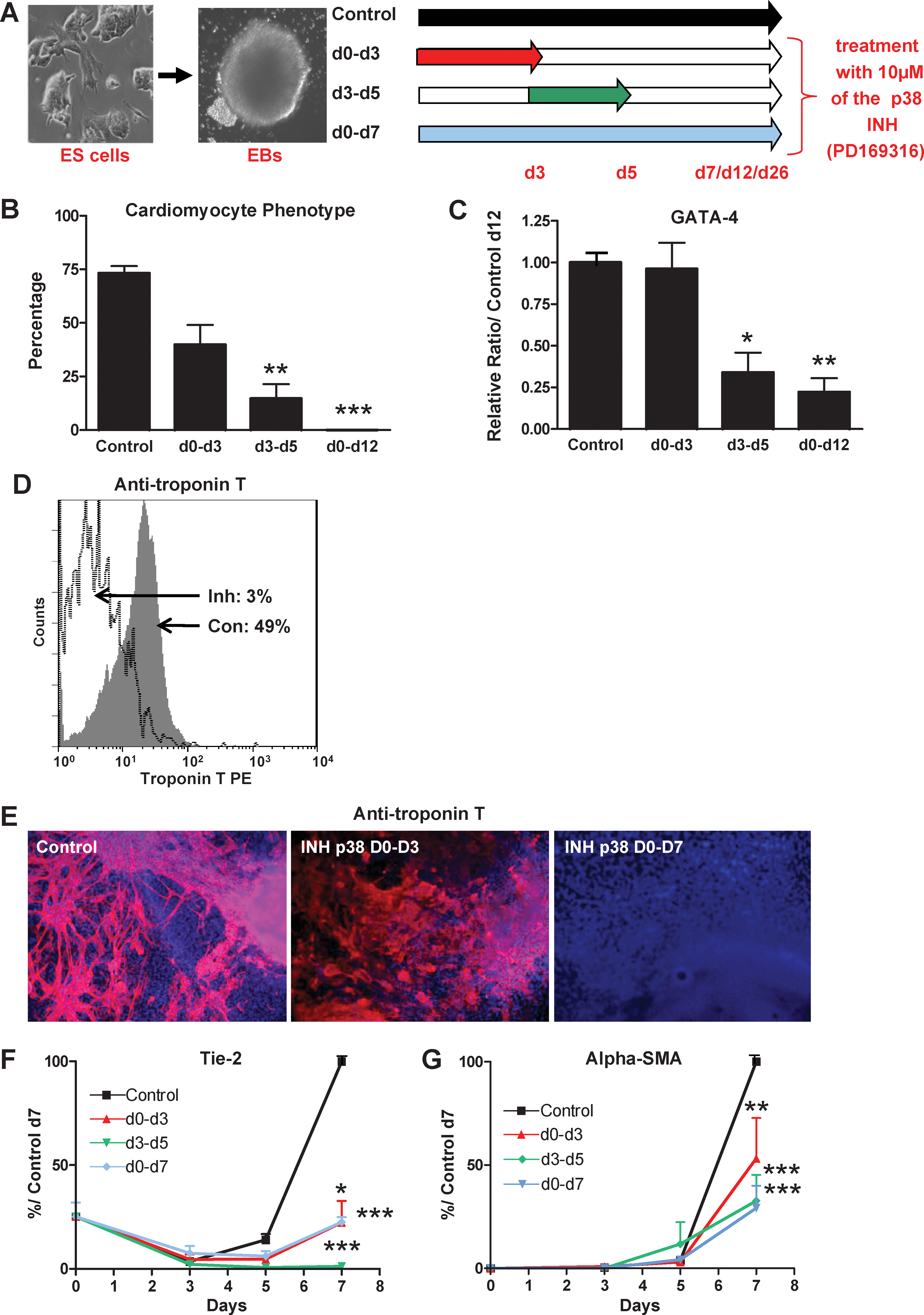
p38MAPK activity is required for 2 early successive steps in mesodermal commitment of ES cells. EBs from wt CGR8 ES cells were differentiated in the presence or absence of PD169316 for different periods as indicated schematically in
Cardiomyogenesis of ES cells is completely inhibited by either the d0–d12 or d3–d5 treatment (Fig. 6B–E and Supplementary Fig. S7A). This inhibition was similar to what was observed in the 1−/− and 15−/− KO cell lines. In contrast, the d0–d3 treatment has a very modest effect, reducing the percentage of EBs with beating cells at day 12 from 75% in control treated cells to 40% and without any effect on gene expression of cardiac markers. Therefore, the major effect on cardiomyogenesis of inhibition of p38MAPK is restricted to the d3–d5 period, coincidental to the spontaneous peak of p38 kinase activity.
We then analyzed the effect of the p38 inhibitor on ES endothelial and smooth muscle differentiation by day 7. In contrast to that observed for cardiomyogenesis, the 3 different treatments were equally efficient to completely block the expression of endothelial and smooth muscle markers (Fig. 6F, G and Supplementary Fig. S7B).
Similar to endothelial and smooth muscle cell formation, we found that wt ES cells require a continuous p38MAPK activity between days 0 and 5 to differentiate, latter on, into the skeletal myogenic lineage. The 3 different p38 inhibitor treatments completely inhibit the formation of contractile myotubes and expression of myogenic markers, evaluated at day 26 (Fig. 7 and Supplementary Fig. S7C).
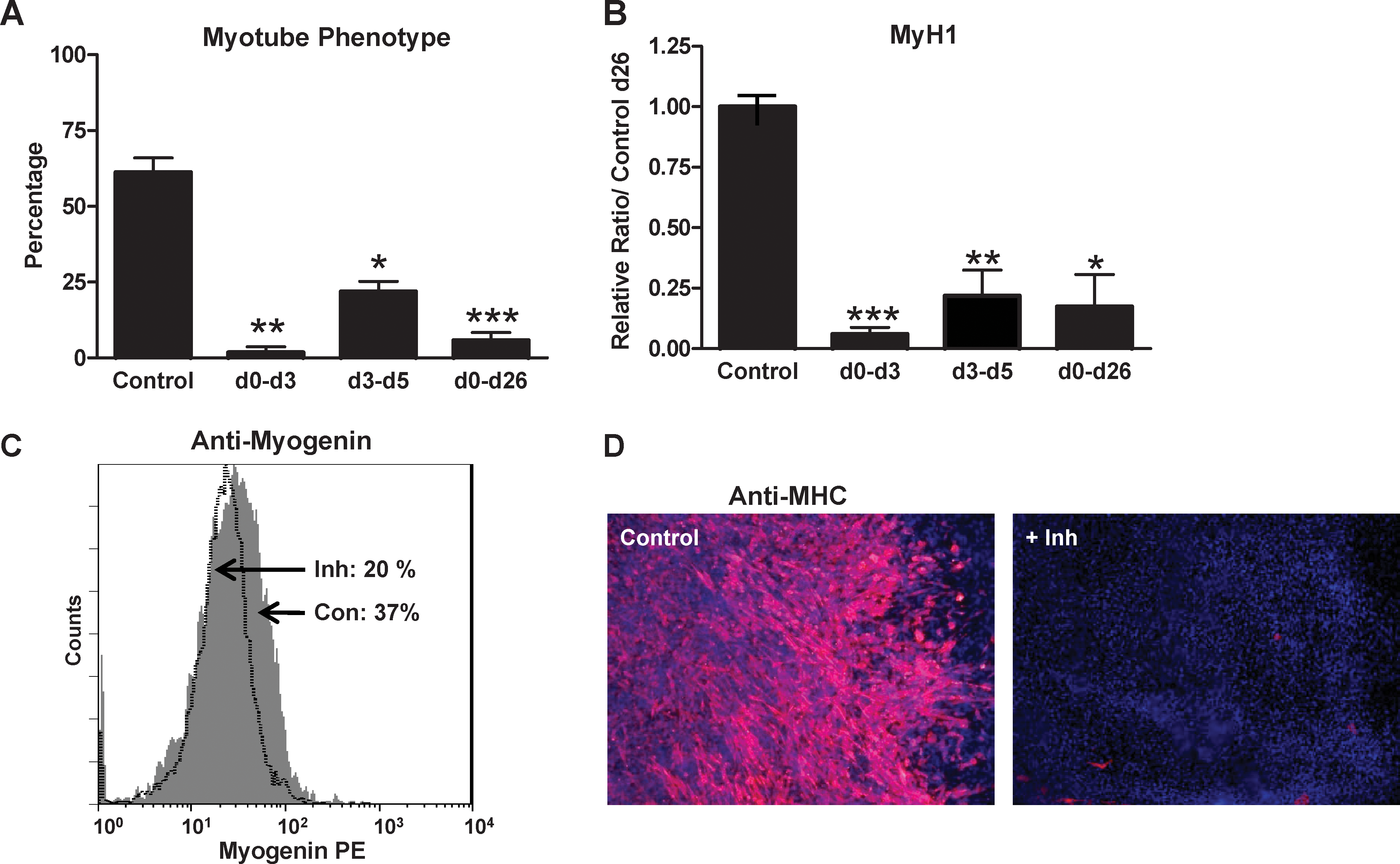
p38MAPK activity is required for 2 early successive steps in skeletal myogenesis of ES cells. EBs from wt CGR8 ES cells were differentiated in the presence or absence of PD169316 for different periods as indicated schematically in Fig. 6A. EBs were plated at day 7 and analyzed at day 26 of differentiation.
p38MAPK activity controls expression of the mesodermal master gene Brachyury
In the mouse embryo, mesoderm is generated from the epiblast through the process of gastrulation, which is initiated at day 6.5 of gestation. The transcription factor-encoding Brachyury gene is specifically expressed in nascent mesoderm and downregulated as these cells differentiate into skeletal muscle, cardiac muscle, blood, and endothelium. In vivo studies have demonstrated the importance of Brachyury in regulating cell fate decisions. Brachyury−/− mice die shortly after gastrulation and display several mesodermal abnormalities [30]. However, the role of this gene in ES cell differentiation in vitro is unknown. Because our results demonstrate the early requirement of p38MAPK in mesodermal commitment of ES cells, we hypothesized that this effect could be mediated by regulation of Brachyury.
Brachyury gene expression was investigated between days 0 and 7 of differentiation of wt, KO, and p38α reexpressing ES cells. As expected, expression of this gene is very low in wt undifferentiated ES cells. Expression profile during EB differentiation shows a peak of expression, centered at day 3, of 32-fold the level at day 0, going down to the basal level of undifferentiated cells by day 5 and staying low until day 7 (Fig. 8). This profile is identical to the one described in the work by Fehling et al. [31], using another wt ES cell line, and is compatible with an important function for this gene in ES cell commitment toward mesoderm lineages. In contrast to wt cells, p38α KO ES cells present no peak of Brachyury expression (Fig. 8A, B). This result is in line with the work of Duval et al. [32], who found that Brachyury gene expression is under the control of p38MAPK pathway. Our results indicate that there is a correlation between mesodermal defects of KO cells and low levels of Brachyury expression. However, both 1−/− and 15−/− reexpressing derivatives do not recapitulate the Brachyury expression profile of wild-type ES cells, expressing Brachyury levels slightly different to KO cells only after day 4 (Fig. 8A, B). Therefore, these results indicate that the restoration of mesodermal differentiation capacities displayed by reexpressing cells cannot be explained by a rescue of Brachyury expression.
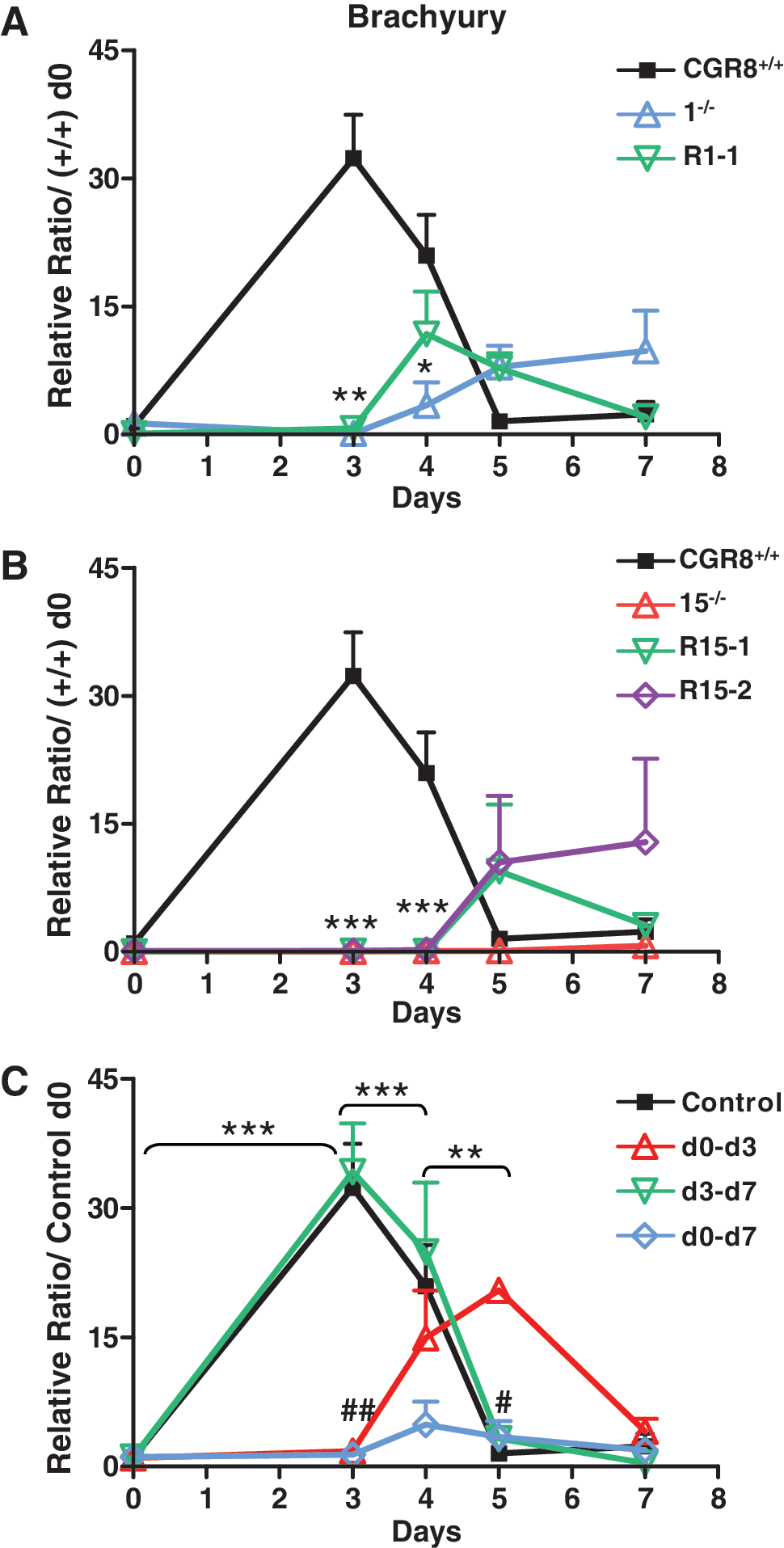
The Brachyury gene expression is regulated by p38MAPK during ES cell differentiation. In
To precisely understand the role of p38MAPK in the regulation of Brachyury gene expression, we then analyzed the effect of the p38 inhibitor treatment on wt ES cells during differentiation (Fig. 8C). The d0–d7 treatment completely blocks the expression of this gene, recapitulating the effect of the genetic KO and showing a correlation between Brachyury inhibition and blockade of mesodermal differentiation. In contrast, the d3–d5 treatment has no effect on Brachyury, leaving the peak of expression unchanged, centered at day 3. Importantly, because this treatment is as efficient as the d0–d7 in inhibiting mesodermal differentiation (Fig. 7), this result demonstrates that p38MAPK can regulate the formation of mesodermal lineages independently of Brachyury regulation. Surprisingly, the d0–d3 treatment blocks Brachyury expression until day 3, but then its expression increases and peaks at day 5 before going down by day 7. This treatment blocks the formation of endothelial and skeletal muscular lineages, leaving cardiomyogenesis almost unchanged (Fig. 7); this indicates that cardiac cells can be formed even if the peak of Brachyury expression at day 3 is absent.
Discussion
By genetic and biochemical approaches we previously described a role of p38MAPK as an early switch of ES cell commitment into cardiomyocytes or neurons [16]. Either gene deletion or specific inhibition of the peak of p38MAPK activity, observed between days 3 and 5 during ES cell differentiation, partially mimicked the cardiomyogenesis inhibition by 10−7 mol/L RA treatment. In the present study, we extended this observation to the endothelial, smooth muscle, and skeletal muscle lineages and found that the p38MAPK requirement can be dissected into 2 successive steps during the differentiation process, one between days 0 and 3 and the second between days 3 and 5. We found that the first step correlates with the p38MAPK regulation of Brachyury gene expression.
A role for p38α in various aspects of cardiomyogenesis, including the regulation of cardiomyocyte differentiation, apoptosis, and hypertrophy, has been described [33 –35]. Interestingly, p38α−/− embryos present a massive reduction of the myocardiac muscle attributed to a defect in placental development [14]. By analysis of both p38α KO ES cells and effects of biochemical inhibition of the pathway, we demonstrate that p38MAPK is required for the cardiomyogenesis of ES cells. Moreover, successive treatments of ES cells with the p38 inhibitor show that the major effect of inhibition of p38MAPK kinase activity on this lineage is evidenced during the d3–d5 period.
In contrast to our results, a recent work described an enhanced cardiomyogenesis of human ES cells by a small molecular inhibitor of p38 MAPK [36]. This apparent discrepancy could be due to different experimental parameters between the 2 studies: 2 different cellular models (human ES cells vs. mouse ES cells); different inhibitors (SB203580 vs. PD169316), which are known to differently affect ES cell differentiation [32]; and importantly, in human ES cells these effects are concentration dependent, with an inhibitory effect at high concentrations, similar to what we found in mouse ES cells.
Circulating endothelial precursor cells provide an endogenous repair mechanism to counteract ongoing risk factor-induced endothelial injury and to replace dysfunctional endothelium [37]. p38MAPK has been involved in both the control of endothelial precursor cell proliferation [38] and the migration of mature endothelial cells [11]. Smooth muscle cells are important contributors to blood vessel integrity. Our study demonstrates that p38MAPK is crucial early on during the differentiation process of ES cells toward the endothelial and smooth muscle lineages. In contrast to cardiomyogenesis, this signal transduction pathway is required continuously between days 0 and 5 for the ES commitment of these lineages. Our data are consistent with the p38MAPK pathway constituting an important potential player in vessel reconstruction from ES cells.
To explain the role of P38MAPK in ES cell differentiation toward cardiac, endothelial, and smooth muscle cells, one simple hypothesis would be that this pathway targets a precursor common to these lineages. For example, such precursor could be located in defined structures that could be represented in vivo by the hemangioblast, which has been defined as the early embryonic structure capable to generate both hematopoietic and vascular lineages [39,40]. This structure is molecularly characterized by the coexpression, during a limited period of time, of Brachyury and Flk-1 genes. Interestingly, we also found these 2 genes to be regulated by p38MAPK in our cellular model. Whether Brachyury and Flk-1 are the only p38MAPK targets required for the control of hemangioblast formation leading to endothelial and smooth muscle lineages necessitate further investigations. However, the fact that cardiomyogenesis presents sensitivity to the p38 inhibitor different from the other lineages suggests the existence of several distinct precursors. Because the d3–d5 treatment with the inhibitor affects differentiation without changing Brachyury gene expression, we hypothesize that, at this precise step, p38MAPK regulates critical unknown mesodermal regulators.
There is an abundant literature showing that p38MAPK activity increased during myoblast differentiation and that this pathway regulates the expression of muscle-specific genes via the control of MyoD and MEF2 muscle-specific transcription factors (for review, see [41]). In vivo, p38MAPK is involved in the developing somite myotome of mice [42]. The p38 MAPK pathway is required for the activation of the quiescent muscle precursor, the satellite cell [43], whereas neither the ERK nor the JNK pathway is involved in the skeletal myogenic process [44,45]. Importantly, myoblasts lacking p38α are unable to differentiate and to form multinucleated myotubes [46]. Our study extends the requirement of p38MAPK in skeletal myogenesis to the early embryonic events of this process. Particularly, analysis of muscle differentiation capacities of either p38α KO ES cells or wt ES cells treated by the p38 inhibitor demonstrated that this pathway is required continuously between days 0 and 5 for skeletal myogenesis.
Altogether our results demonstrate that the p38MAPK activity is required continuously between days 0 and 5 for cardiac, endothelial, smooth muscle, and skeletal muscle mesodermal commitment and, conversely, inhibits neurogenesis. We were able to dissect this requirement into 2 successive steps: the first one between days 0 and 3 corresponds to basal p38 activity necessary for the formation of endothelial, smooth muscle, and skeletal muscle lineages and correlates with regulation of Brachyury expression, whereas the second step corresponds to the spontaneous peak of p38MAPK activity between days 3 and 5 [16]; in addition to the former mesodermal lineages, cardiomyogenesis is controlled at this step.
Owing to the known biological role of Brachyury and the effect of p38MAPK inhibition on the expression of this gene, one can hypothesize that the d0–d3 p38MAPK biological function in ES commitment is mediated by Brachyury. However, the reexpressing cell lines, although rescuing the mesodermal defects displayed by p38MAPK KO ES cells, do not recapitulate wt Brachyury expression, suggesting that this gene is not involved in this p38MAPK function. Importantly, it is noteworthy that the mesodermal rescues and the reexpressed p38MAPK protein levels are partial. Therefore, it is possible that the peak of Brachyury expression at day 3 requires wt p38MAPK levels and, in turn, that this peak is necessary for the full mesodermal response of ES cells. Further works on Brachyury regulation are necessary to decipher the molecular links between p38MAPK, Brachyury, and mesodermal commitment of ES cells.
Regarding the control of mouse ES cell differentiation, it is noteworthy that the p38MAPK pathway displays unique biological properties when compared with ERK and JNK MAPK pathways. Indeed, we found that p38MAPK constitutes a major switch between mesodermal lineages (p38 on) and neurogenesis (p38 off), whereas the ERK pathway plays a different control earlier, because ERK2 KO cells are blocked in both mesodermal and neuronal differentiations [47], and JNK is involved only in neurogenesis [48].
Owing to the fact that p38MAPK pathway controls ES cell commitment toward mesodermal lineages, our results suggest that the precise modulation of this pathway early on during ES cell differentiation would help their use in regenerative medicine by improving the efficiency of differentiation into specific lineages.
Footnotes
Acknowledgments
The authors thank Drs. B.P. Sleckman and David Engelberg for providing the p38 KO ES cell lines and p38 cDNA clones, respectively. This work was supported by a grant from the Association Française contre les Myopathies. E.B. was supported by fellowships from the Groupe d'Eude sur l'Hémostase et la Thrombose and the Fondation pour la Recherche Médicale. O.H. was supported by a fellowship from the Association Française contre les Myopathies.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.