
Abstract
Human bone marrow mesenchymal stromal/stem cells (BMSCs) have been reported to possess immunomodulatory functions with the capacity to suppress immune reactions partly mediated by immunosuppressive factors, indoleamine 2,3-dioxygenase and nitric oxide, and suggested to be potentially applicable for therapeutic use. More recently, other fibroblast-like cells have been reported to possess similar properties. Here we demonstrate that human foreskin fibroblasts (HFFs) express an MSC-like immunophenotype and possess immunosuppressive properties similar to BMSCs but lack the capacity for osteogenic and adipogenic differentiation. HFFs suppressed human peripheral blood mononuclear cells (PBMC) proliferation stimulated with mitogen or in an allogeneic mixed lymphocyte reaction comparable to BMSCs. However, HFFs showed undetectable levels of indoleamine 2,3-dioxygenase and inducible nitric oxide synthase expression, in contrast to BMSCs when cocultured with activated PBMCs. To identify HFF specific immunosuppressive factors, we performed array profiling of common cytokines expressed by HFFs and BMSCs alone or when cocultured with activated PBMCs. Real-time polymerase chain reaction analysis confirmed that multiple factors were upregulated in HFFs cocultured with activated PBMCs compared with HFFs alone or BMSCs cultured under the same conditions. Functional assays identified interferon-α as the major immunosuppressive mediator expressed by HFFs. These results suggest that the HFFs possess immunosuppressive properties, which are mediated by alternate mechanisms to that reported for BMSCs.
Introduction
B
The various mechanisms by which different MSC populations mediate immune modulatory effects have been reported. Some studies have shown that soluble factors including transforming growth factor β1 (TGF-β1), hepatocyte growth factor (HGF) [5,13], and prostaglandin E2 [14] play important roles in mediating the immunosuppressive effects of BMSCs, and other reports have suggested that cell–cell contact is also important for the immunomodulatory effects of BMSCs [15,16]. In addition, the intracellular enzyme, indoleamine 2,3-dioxygenase (IDO), has been shown to be one of the critical immunosuppressive factors produced by BMSC-like populations [13,17,18]. This enzyme has previously been identified in antigen-presenting cells as being involved in a major immunosuppressive effector pathway that inhibits T-cell responses to autoantigens and fetal alloantigens in vivo [19 –22]. IDO induced by interferon γ (IFN-γ) and other proinflammatory cytokines catalyzes and degrades tryptophan, an essential amino acid for cell growth [23]. Nitric oxide (NO) has been also reported to be one of the major mediators of T-cell suppression by BMSCs [24]. NO produced by inducible NO synthase (iNOS) in BMSCs suppresses phosphorylation of Stat5, which plays an important role in T-cell proliferation signaling pathway [25].
As MSCs derived from various tissues such as bone marrow [4 –7], placenta [18,26], adipose tissue [9], dental pulp [27,28], periodontal ligament [27], and gingival tissue [29] show antiproliferative, immunomodulatory, and anti-inflammatory effects in vitro or in vivo, it has been suggested that immunosuppressive properties are unique and specialized to MSCs [30]. However, some reports have demonstrated that fibroblasts from gingival tissue [27,31], cornel tissue [32], foreskin [33], synovial membrane [34], and dermis [34,35] also display immunosuppressive functions, which may be mediated by IDO similar to MSCs [27,33,35]. These studies suggest that the capacity of such cells to mediate immunosuppression may be a common property attributed to cultured fibroblast-like cell populations derived from tissues of different developmental origins.
Dermal substitutes containing allogeneic skin fibroblasts have been utilized for various clinical applications including the treatment of burns [36], chronic venous ulcers [37,38], and several other dermatological conditions and plastic/reconstructive surgery and have achieved the promising results [39]. One study demonstrated that treatment for venous ulcers using cultured allogeneic human skin equivalent containing allogeneic human dermal fibroblasts showed no symptoms or signs of rejection in recipients and no immune responses against fibroblasts in vitro [37]. Therefore, skin fibroblasts may offer an alternate allogeneic cell source for modulating immune responses. Accordingly, the aims of this study were to examine the immunomodulatory properties of human foreskin fibroblasts (HFFs) compared with cultured human BMSCs and determine the immunosuppressive mechanism(s) of HFFs using coculture systems with activated lymphocytes.
Materials and Methods
Cell culture
HFFs were isolated by collagenase I (3 mg/mL; Worthington Biochemical, Lakewood, NJ) and dispase II (4 mg/mL; Roche Diagnostics, Indianapolis, IN) digestion for 2 h at 37°C of foreskin specimens from 3 neonatal male donors with informed parental consent, at the Flinders Medical Centre, Adelaide, SA, Australia. Primary BMSC cultures were established by plating bone marrow mononuclear cells that were prepared from 3 normal human bone marrow aspirates obtained from healthy consenting adult donors (approved by the Royal Adelaide Human Ethics Committee) by density gradient centrifugation on Ficoll-Isopaque (Lymphoprep; Fresenius Kabi Norge AS, Olso, Norway) as previously described [2,40]. Ex vivo expanded HFFs and BMSCs were maintained in α-MEM (Sigma-Aldrich, St. Louis, MO) with 10% FCS (Thermo Electron, Melbourne, Australia), 2 mM
Differentiation assays
For osteogenic cultures, BMSCs and HFFs at passage 6 were seeded at 8 × 103 per cm2 in 24-well plates and cells were cultured in α-MEM supplemented with 5% fetal calf serum (FCS), 100 μM
Antibody reagents
Recombinant human TGF-β1, TGF-β3, IFN-α (IFN-αB2), IFN-β, and HGF were purchased from PeproTech (Rocky Hill, NJ) and NODAL was from R&D Systems (Minneapolis, MN). Neutralizing monoclonal antibody (Ab) against human TGF-β1 was from R&D Systems, and neutralizing polyclonal Ab against human IFN-α and monoclonal Ab against human IFN-β were from PBL Biomedical Laboratories (New Brunswick, NJ). Rabbit control IgG (Thermo Scientific, Rockford, IL) and mouse control IgG1 (1B5; kind gift from Prof. L.K. Ashman, University of New Castle, NSW, Australia).
Effects of neutralizing Abs against cytokines on allogeneic MLR with conditioned media of HFFs
To identify soluble factors responsible for the inhibitory effect of HFFs, neutralizing antibodies against selected cytokines were added to the allogeneic MLR-containing conditioned media (CM) of HFF cultures. Briefly, CM of HFFs (3 × 104) and BMSCs (3 × 104) at passage 3 cocultured with Concanavalin A (Con A)–stimulated PBMCs (1 × 105) in a 96-well round-bottomed plate for 5 days were collected and filtrated using a 0.45-μm pore-sized filter (Sartorius Stedim Biotech, Goettingen, Germany). Subsequently, CM (20% of whole media) treated with neutralizing Abs against TGF-β, IFN-α, and IFN-β (5 μg/mL) for 2 h at 37°C were applied to Con A-stimulated PBMC assay. All the experiments were performed using fresh CM.
Flow cytometric analysis
Passage 3 cells were harvested as single-cell suspensions by trypsin/ethylenediaminetetraacetic acid digestion and incubated in blocking buffer containing Hank's balanced salt solution (Sigma-Aldrich) supplemented with 5% FCS (Thermo Electron), 1% bovine serum albumin (ICN Biomedicals, Aurora, OH), and 5% normal human serum (Australian Red Cross, Adelaide, SA, Australia) for 30 min. A total of 1 × 105 cells were resuspended in Abs (10 μg/mL) specific for cell surface markers, which are CD14, CD34, CD45 (Beckman Coulter, Marseille, France), CD40, CD54, CD73, CD80, CD86, CD90, CD105, CD166, HLA-DR (BD Bioscience, San Jose, CA), CD44 (Clone H9H11; kindly provided by Dr. Andrew Zannettino, Division of Haematology, IMVS, Adelaide, SA, Australia), CD106 (Clone QE4G9; kindly provided by Dr. Ravi Krishnan, Transplantation Immunology Laboratory, The Queen Elizabeth Hospital, Adelaide, SA, Australia), CD146 (CC9) [41], STRO-1 [42], STRO-3 (CA12) [43], STRO-4 (heat shock protein-90) [44], HLA-ABC (7B6c) [45], or isotype-matched nonbinding control immunoglobulin, 1B5 (IgG1), 1D4.5 (IgG2), and 1A6.12 (IgM) (kindly provided by Prof. L.K. Ashman, The University of Newcastle, Newcastle, NSW, Australia) (10 μg/mL) for 1 h on ice. After washing, cells were incubated with secondary detection reagents, goat anti-mouse IgG-PE, IgG-FITC, or IgM-FITC Abs (Southern Biotechnology Associates, Birmingham, AL) for 45 min on ice. After washing and fixing, samples were analyzed using an Epics-XL/MCL flow cytometer (Beckman Coulter, Hialeah, FL).
PBMC proliferation assay with mitogenic stimulation Con A-stimulated PBMC assay and allogeneic MLR
PBMC proliferation assay following mitogenic stimulation with Con A-stimulated PBMC assay and allogeneic MLR assay were performed as previously described [27]. Human PBMCs were purified from heparinized blood collected from healthy consenting adult donors (approved by the Royal Adelaide Human Ethics Committee) by density gradient centrifugation on Ficoll-Isopaque [46]. PBMCs were cultivated in RPMI-1640 medium (Sigma-Aldrich) supplemented with 10% heat-inactivated FCS, 2 mM
To assess cell division and cell proliferation, PBMCs were labeled with the green fluorescence dye carboxyfluorescein succinimidyl ester (CFSE; Invitrogen, Eugene, OR) [47,48]. In brief, PBMCs were incubated with 2 μM CFSE for 15 min at 37°C in Hank's balanced salt solution with 0.1% bovine serum albumin (Sigma Chemical, St. Louis, MO) and washed 3 times with RPMI-1640 + 10% FCS before seeding the cells into tissue culture plates. After 5 days in culture, PBMCs were analyzed by flow cytometry (Epics-XL/MCL flow cytometer; Beckman Coulter) to detect green fluorescence. Analysis of cell division and proliferation rate was performed using ModFit software (Verity Software House, Topsham, ME). All culture assays were performed in 200 μL RPMI-1640 using a 96-well round-bottomed or flat-bottomed plate (Becton Dickinson). All experiments were run in triplicate.
Quantitative real time–polymerase chain reaction analysis
Total RNA was prepared using Trizol (Invitrogen Life Technologies, La Jolla, CA) according to the manufacturer's protocol. First-strand cDNA was synthesized using Oligo dT primer and Superscript II reverse transcriptase (Invitrogen) according to manufacturer's instructions. Real-time polymerase chain reaction (PCR) was performed with RT2 SYBR Green/ROX qPCR Master Mix (SuperArray Bioscience, Frederick, MD) using the Rotor-Gene 6000 Real-Time Thermal Cycler (Corbett Research, Sydney, Australia) (50°C/2 min; 95°C/15 min; 40 cycles of 95°C/15 s, 60°C/26 s, and 72°C/10 s; 72°C/3 min). Primer (GeneWorks, Hindmarsh, Australia) sequences used were as follows: IDO (F 5′-agagtcaaatccctcagtcc-3′/R 5′-aaatcagtgcctccagttcc-3′), iNOS (F 5′-cctgagctcttcgaaatccca-3′/R 5′-cccgaaaccactcgtatttgg), TGF-β1 (F 5′-cacgtggagctgtaccagaa-3′/R 5′-gaacccgttgatgtccactt-3′), TGF-β3 (F 5′-aaattcgacatgatccaggg-3′/R 5′-ctcgatcctctgctcattcc-3′), HGF (F 5′-caatagcatgtcaagtggag-3′/R 5′-ctgtgttcg tgtggtatcat-3′), NODAL (F 5′-agacatcatccgcagcctac-3′/R 5′-aggtgacctgggacaaagtg-3′), IFNA1 (F 5′-ggagtttgatggcaaccagt-3′/R 5′-gagtttctcccaccctctcc-3′), IFNA5 (F 5′-ggtgctcaactgcaagtcaa-3′/R 5′-tcccatttgtgccattatca-3′), IFNA8 (F 5′-cctgcctg aaggacagacat-3′/R 5′-tcctgcatcacacaggactc-3′), IFNB1 (F 5′-ttgttgagaacctcctggct-3′/R 5′-tccttggccttcaggtaatg-3′), and β-actin (F 5′-gatcattgctcctcctgagc-3′/R 5′-gtcatagtccgcctagaagcat-3′). β-Actin was used as an internal control.
PCR array
Passage 4 HFFs and BMSCs (1 × 106) were seeded in the bottom chambers of 24-transwell plates with 0.4-μm pore-sized membranes (Nunc, Roskilde, Denmark), before the addition of either Con A-stimulated PBMCs or allogeneic MLR (3 × 105) in the top chambers in 0.4 mL RPMI-1640 for 3 days. Total RNA was prepared using Trizol (Invitrogen) and cleaned up with RNeasy Mini Kit (Qiagen, Hilden, Germany). Isolated total RNA was then subjected to reverse transcription using RT2 First-Strand Kit (SABiosciences, Frederick, MD) according to manufacturer's instructions. Real-time PCR was performed in the 96-well plate of RT2 Profiler PCR Array for human common cytokines (PAHS-021; SABiosciences) with RT2 SYBR Green/Fluorescein qPCR Master Mix (SABiosciences) using the iQ5 real-time PCR detection systems (Bio-Rad, Foster City, CA) (95°C/10 min; 40 cycles of 95°C/15 s and 60°C/60 s).
Quantitative analysis of IFN-α protein level
The levels of IFN-α in the CM of HFFs or BMSCs cocultured with or without Con A-stimulated PBMCs were detected using human IFN-α Multi-Subtype ELISA Kit (PBL InterferonSource, Picataway, NJ) according to the manufacturer's instructions. CM were prepared as described earlier.
Statistical analysis
All values are expressed as mean ± standard deviation from 3 independent experiments. To test statistically significant differences between paired observations, the Student's t-test for paired data was used. A P value of <0.01 was considered statistically significant.
Results
Cell surface marker characteristics of HFFs
The immunophenotype of HFFs was determined by flow cytometric analysis utilizing Abs against cell surface markers (Fig. 1A). HFFs were strongly positive for HLA-ABC (major histocompatibility complex (MHC) class I antigen), CD44, CD90 (markers associated with stromal cells), CD73, CD105, and CD166 (markers associated with stromal cells and endothelial cells), but were negative for HLA-DR (MHC class II antigen), CD14, CD34, CD40, CD45, CD80, and CD86 (markers of hematopoietic progenitor and immune cells). When cultured under either osteogenic or adipogenic inductive conditions, HFFs failed to form a mineralized matrix or lipid-containing adipocytes, respectively, in comparison to BMSCs (Fig. 1B).

Characterization of HFFs.
HFFs suppress activated PBMC proliferation
To examine whether HFFs affect the proliferation of 3H-thymidine–labeled PBMCs, HFFs were cocultured with PBMCs either treated with the mitogen stimulator, Con A (Con A-stimulated PBMC assay), or cocultured with allogeneic irradiated PBMCs (allogeneic MLR). Suppression of PBMC proliferation was observed when different numbers of HFFs (1 × 103 to 1 × 105) were added to Con A-stimulated PBMCs or allogeneic MLR cultures (Fig. 2A, B). HFFs (1 × 105) inhibited PBMC proliferation at similar levels to BMSCs (1 × 105) in both assays (Fig. 2C, D). We also examined the number of cell divisions exhibited by CFSE-labeled PBMCs cocultured with HFFs or BMSCs, or without cells in Con A-stimulated PBMC assay (Fig. 2E). Con A-stimulated PBMCs cocultured with 1 × 105 HFFs showed that the number of cell divisions peaked at generation 0 (G0) similar to BMSCs and colcemid-treated group, whereas the cell division peak of PBMCs cultured in the absence of other cells developed to G5 during the same time period. These results suggested that HFFs have a similar function to suppress PBMC proliferation compared with BMSCs. Further analysis of PBMCs cocultured with either HFFs or BMSCs demonstrated no increase in the number of apoptotic cells when compared with control cultures containing activated PBMCs alone (Supplementary Fig. S1, available online at
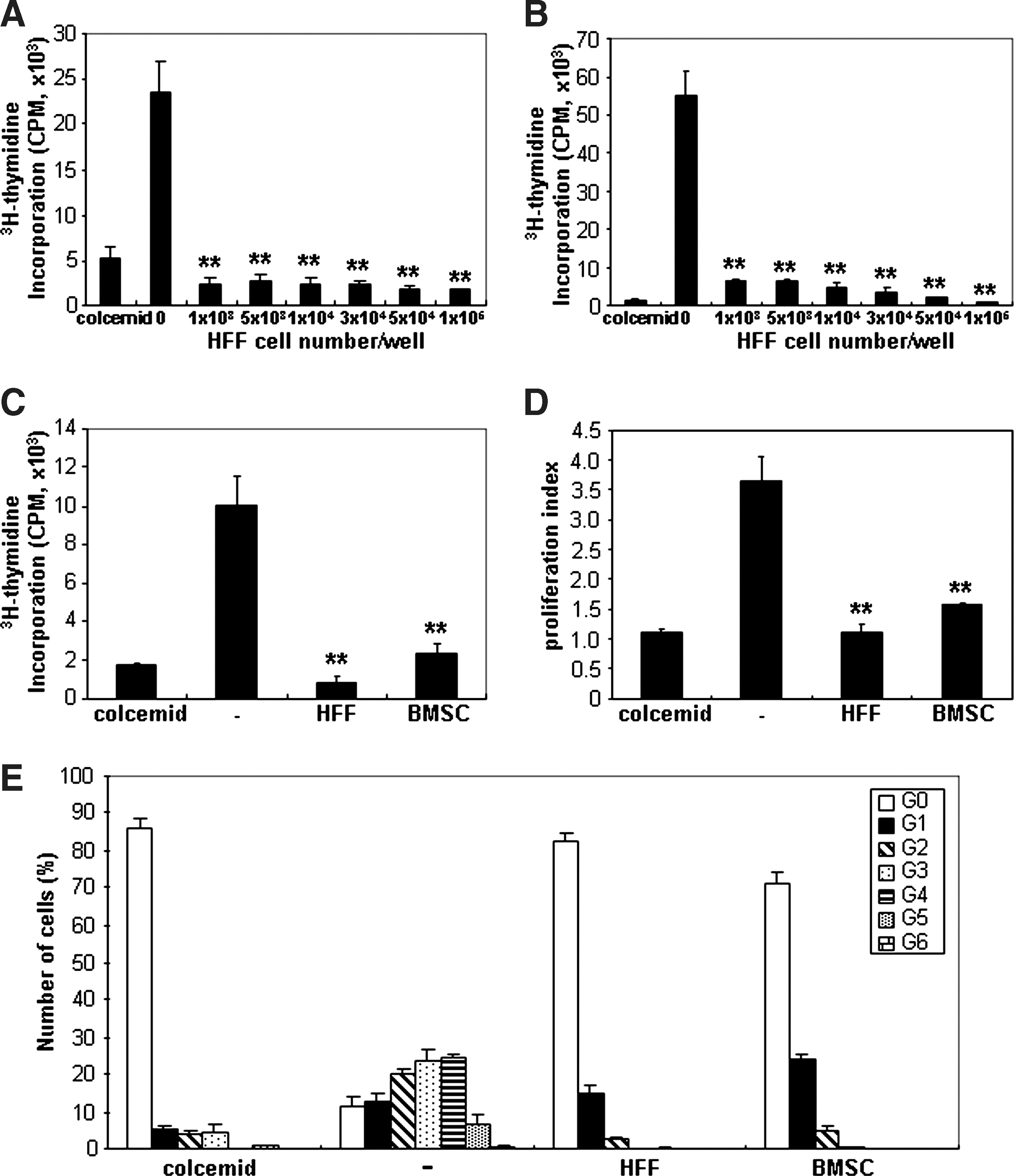
Inhibitory effects of allogeneic HFFs on peripheral blood mononuclear cells (PBMC) proliferation. 3H-thymidine–labeled PBMCs were cocultured with increasing numbers of HFFs (1 × 103 to 1 × 105) stimulated with either
Activated PBMCs do not induce IDO or iNOS expression in HFFs
The expression of IDO in HFFs or BMSCs was investigated following coculture with Con A-stimulated PBMCs or allogeneic MLR using transwells. IDO expression in HFFs cocultured with Con A-stimulated PBMCs or allogeneic MLR was found to be significantly lower than BMSCs in both assays (Fig. 3A). Further, we investigated the expression of IDO by HFFs stimulated with IFN-γ for 24 and 48 h as previously described for BMSCs [27]. In this study, BMSCs stimulated with IFN-γ increased IDO expression intensely from basal levels. In contrast, IFN-γ showed negligible effects on IDO expression in HFFs at both time points (Fig. 3B). In parallel experiments, iNOS expression was also found to be largely unaffected in HFFs when cocultured with activated PBMCs (Fig. 3C) or treated with IFN-γ (data not shown), compared with BMSCs. From these results, it appears that the HFFs' immunosuppressive function may occur through mechanisms different from the IDO- and iNOS-mediated immunomodulatory properties of BMSCs.
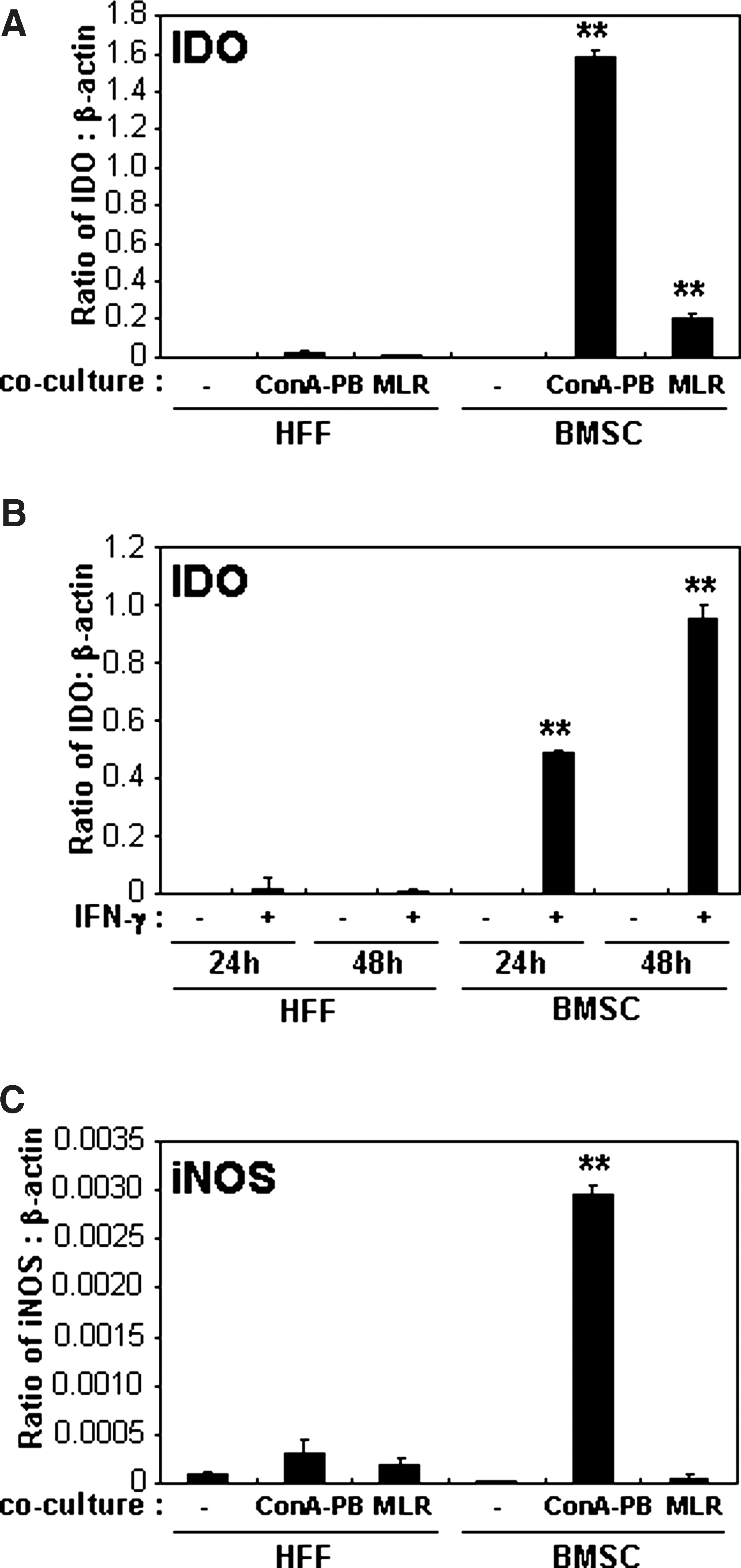
Comparison of IDO and iNOS expression in HFFs and BMSCs.
Expression of cytokines upregulated in HFFs cocultured with activated PBMCs
A comprehensive analysis of the cytokine expression of HFFs was performed using the RT2 Profiler PCR Array for human common cytokines (Table 1). Total RNA was isolated from cultured HFFs and BMSCs and following coculture with Con A-stimulated PBMCs or allogeneic MLR using transwells. Various cytokines were highly expressed by HFFs cocultured with Con A-stimulated PBMCs and allogeneic MLR in comparison to HFFs cultured alone or BMSCs cocultured with or without activated PBMCs (Table 1). From these results, TGF-β1, TGF-β3, nodal homolog (mouse) (NODAL), IFN-α1, IFN-α2, IFN-α5, IFN-α8, IFN-β1, interleukin 10 (IL-10), tumor necrosis factor superfamily 13b (BAFF), and tumor necrosis factor superfamily 13 (APRIL) were identified as potential HFF-mediated immunosuppressive factors (Table 1). Studies using real-time PCR confirmed the differential expression of TGF-β1, TGF-β3, NODAL, IFN-α1, IFN-α5, IFN-α8, and IFN-β1 to be higher in HFFs cocultured with either Con A-stimulated PBMCs or allogeneic MLR (Fig. 4), in contrast to IFN-α2, IL-10, BAFF, and APRIL (data not shown). As HGF has been reported to inhibit PBMC proliferation [5], we also studied the effect of HGF expression by HFFs cocultured with PBMCs and found its inhibitory effect on PBMC proliferation to be higher than HFFs alone and BMSCs cultured under different conditions (Fig. 4).
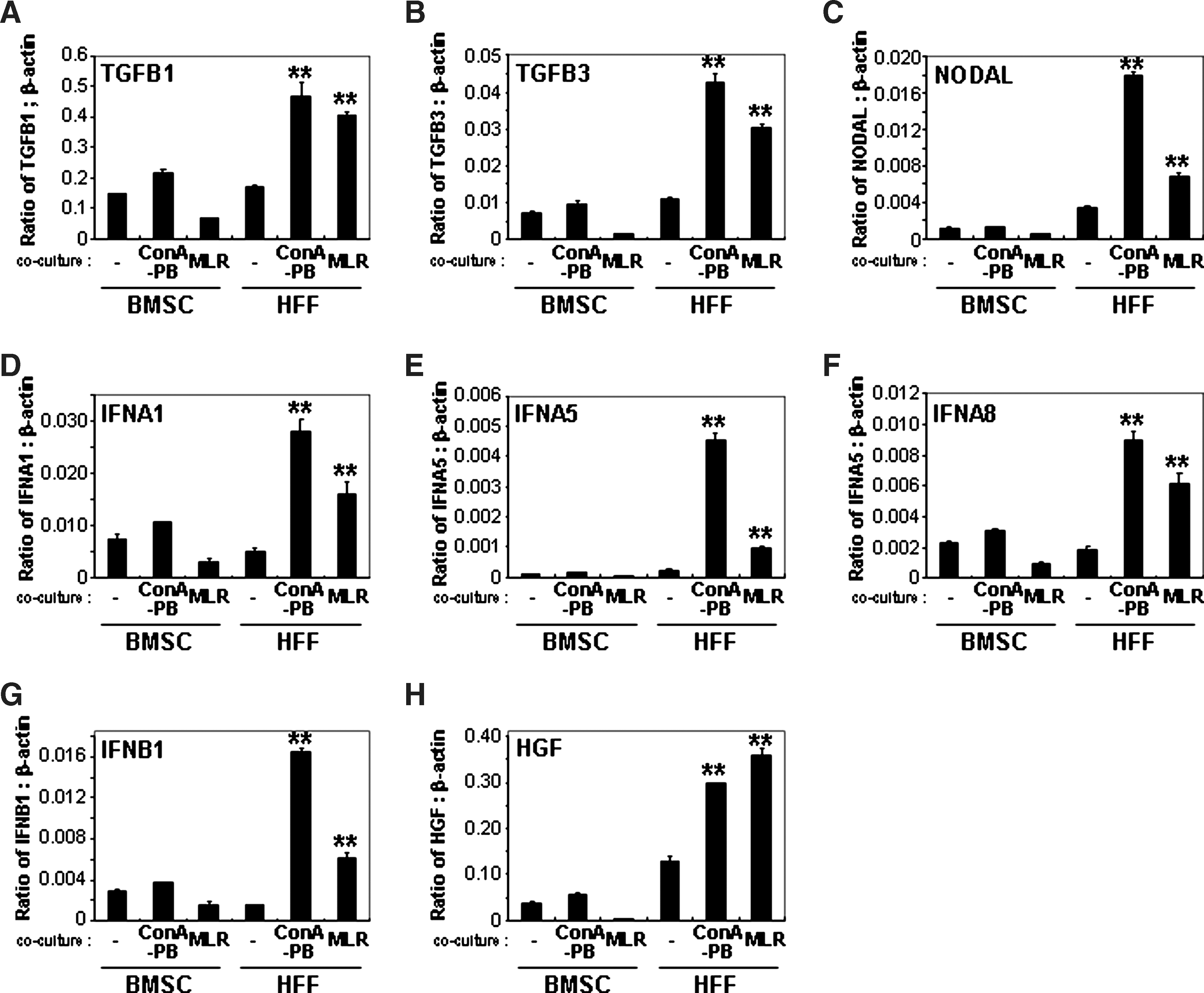
Gene expression in HFFs and BMSCs cocultured with Con A-stimulated PBMCs or allogeneic MLR. The expression of TGF-β1
BMSCs and HFFs were cocultured with Con A-stimulated peripheral blood mononuclear cells (PBMC) assay (Con A-PB) or allogeneic MLR using transwell plates. The expression of cytokines was analyzed by real-time polymerase chain reaction using a 96-well plate of RT2 Profiler Polymerase Chain Reaction Array for human common cytokines (PAHS-021; SABiosciences). The factors with underline in the Gene names column were picked up and the expression of these factors was confirmed in Fig. 4. A value of 2-fold change is represented in boldface.
BMSCs, bone marrow mesenchymal stromal/stem cells; MLR, mixed lymphocyte reaction; Con A, concanavalin A; IL1F9, interleukin 1 family member 9; IL7, interleukin 7; CD70, CD70 molecule; IFNA8, interferon alpha 8; IL8, interleukin 8; IFNA2, interferon alpha 2; IFNB1, interferon beta 1; TNFSF13B, tumor necrosis factor (ligand) superfamily member 13b; IFNA5, interferon alpha 5; IL1A, interleukin 1 alpha; CSF1, colony stimulating factor 1 (macrophage); IL12A, interleukin 12A; IL24, interleukin 24; TNFRSF11B, osteoprotegerin (tumor necrosis factor receptor superfamily member 11b); IFNK, interferon kappa; IL1F8, interleukin 1 family member 8; IL10, interleukin 10; NODAL, nodal homolog (mouse); BMP2, bone morphogenetic protein 2; LEFTY2, left-right determination factor 2; TGFB3, transforming growth factor beta 3; IL18, interleukin 18; GDF9, growth differentiation factor 9; BMP6, bone morphogenetic protein 6; IL6, interleukin 6; IL1B, interleukin 1, beta; IFNA1, interferon alpha 1; TGFB1, transforming growth factor beta 1; LTA, lymphotoxin alpha (TNF superfamily member 1); TNF, tumor necrosis factor (TNF superfamily member 2); TNFSF13, tumor necrosis factor (ligand) superfamily member 13; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; ND, not done.
Effects of cytokines on PBMC proliferation
To investigate the effects of selected cytokines (TGF-β1, TGF-β3, NODAL, IFN-α, IFN-β, and HGF) on PBMC proliferation, recombinant proteins of these cytokines were added to the Con A-stimulated PBMC assay. TGF-β3 (10, 100 ng/mL, and 1 μg/mL), TGF-β1, IFN-α, and IFN-β (1, 10, 100 ng/mL and 1 μg/mL) significantly suppressed PBMC proliferation, whereas NODAL and HGF failed to show any significant inhibitory effects (Fig. 5). The addition of TGF-β1, IFN-α, and IFN-β caused a strong inhibition of PBMC proliferation. To confirm the inhibitory nature of these cytokines to suppress PBMC proliferation, neutralizing Abs against TGF-β1, IFN-α, and IFN-β were added to Con A-stimulated PBMC assays cultured with CM derived from either HFFs or BMSCs cocultured with Con A-stimulated PBMCs for 5 days. The CM derived from HFFs and BMSCs alone (20% of whole media) significantly inhibited PBMC proliferation (Fig. 6A). The addition of neutralizing Abs against TGF-β1 and IFN-β (5 μg/mL) failed to inhibit the suppressive effects on PBMC proliferation when cultured with CM derived from both HFFs and BMSCs (Fig. 6A). However, the addition of a neutralizing Ab against IFN-α (5 μg/mL) partly restored PBMC proliferation suppression mediated by CM derived from treated HFFs, but had no effect on CM derived from treated BMSCs (Fig. 6A). The secretion of IFN-α protein was measured by ELISA using media from HFFs or BMSCs cocultured with or without Con A-stimulated PBMCs (Fig. 6B). Levels of IFN-α protein secreted by treated HFFs were significantly upregulated compared with nontreated HFFs. In addition, IFN-α levels by HFFs (118.25 ± 0.182 pg/mL) were significantly higher than BMSCs (36.8 ± 0.364 pg/mL) when cocultured with Con A-stimulated PBMCs. The addition of recombinant IFN-α to Con A-stimulated PBMCs demonstrated that the effects of IFN-α (0.01–10 ng/mL) on suppression of PBMC proliferation were dose dependent.
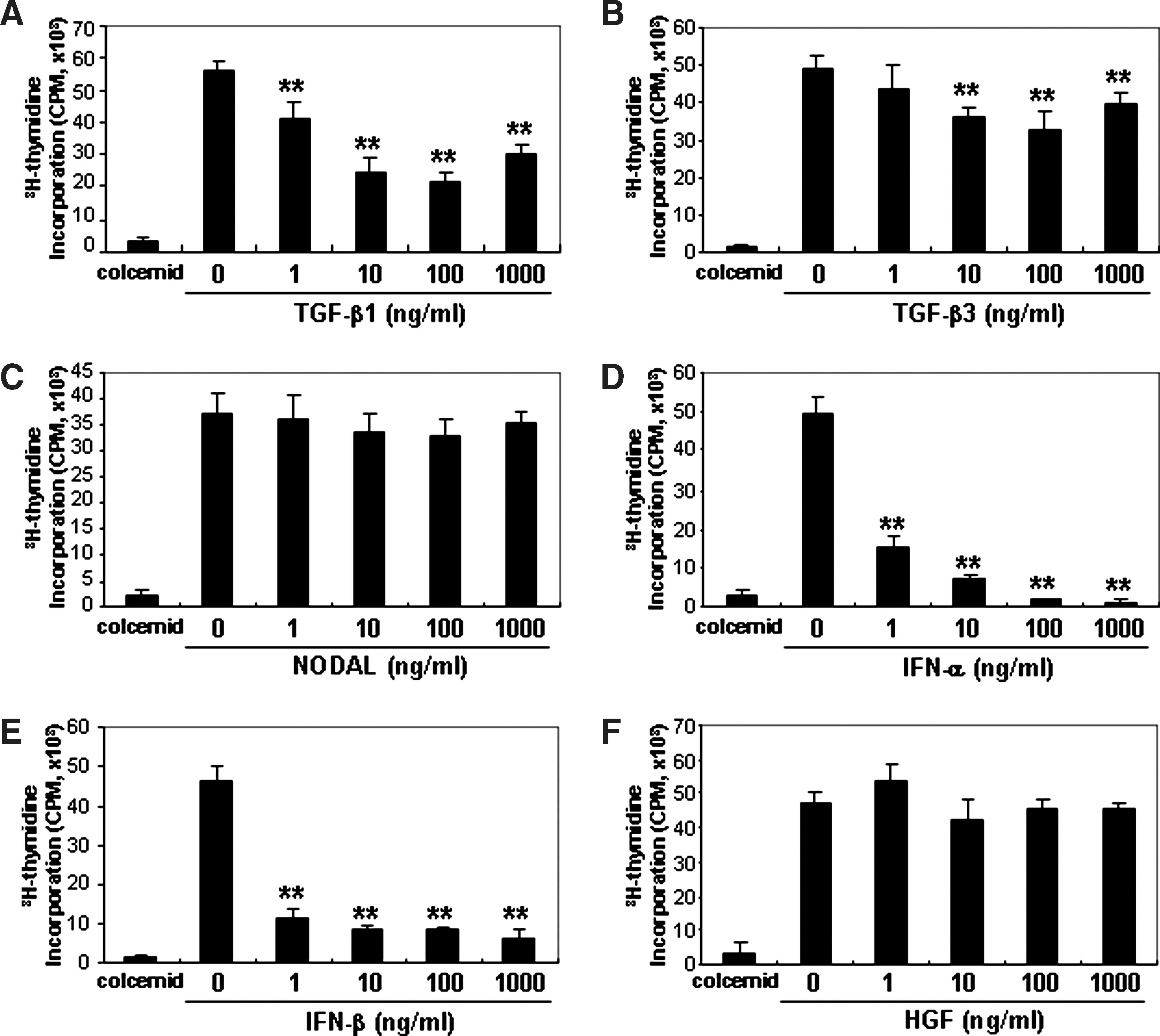
Effects of cytokines on PBMC proliferation stimulated with Con A. Recombinant proteins of TGF-β1
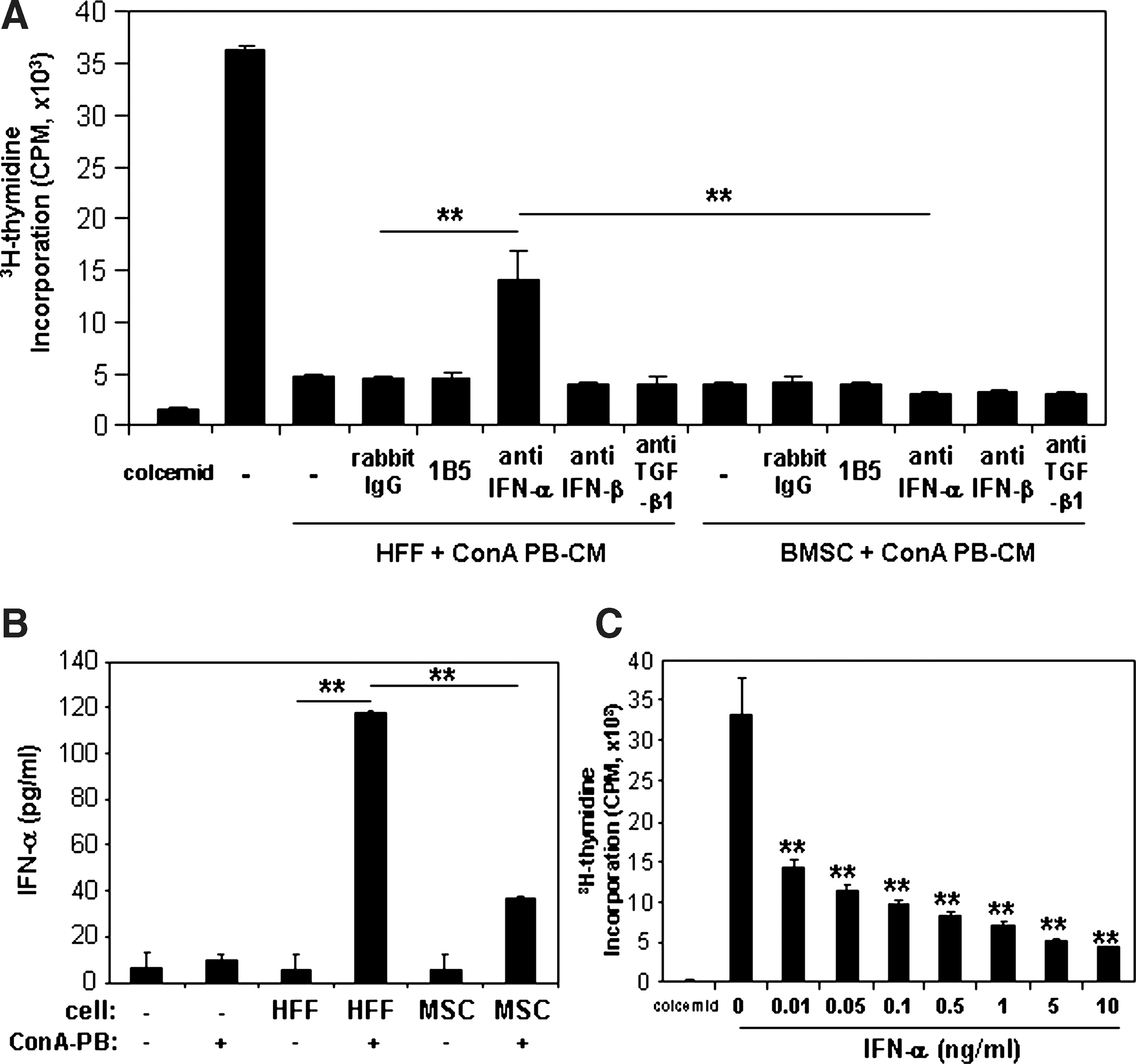
IFN-α secreted by HFFs cocultured with activated PBMCs suppressed Con A-stimulated PBMC proliferation.
Discussion
In this study, we showed that HFFs exhibited similar immunomodulatory properties to BMSCs but used an alternate mechanism to mediate the immunosuppression of activated PBMCs. Accordingly, a comparison of the cytokine gene expression profiles between HFFs and BMSCs cocultured with activated PBMCs was undertaken to identify potential novel factors that regulate these immune responses. Recent in vivo studies have demonstrated the efficacy of allogeneic BMSCs to help manage a wide range of conditions through suppression of immune responses and inflammatory reactions [4,7 –12]. The underlying feature of these findings is the capacity of BMSCs to modulate the proliferation and function of several types of immune cells such as CD4 + T lymphocytes, CD8 + T lymphocytes, and NK cells [49]. More recently, other studies have shown that this phenomenon is not a unique and specialized property of BMSCs, but may also be a feature of other fibroblast-like cells isolated from various tissues [27,31 –33,35].
Several studies have reported that intravenous administration of allogeneic MSCs from various tissues suppresses host immune reactions to autoimmune attack [8], allogeneic haploidentical hematopoietic grafts [9], progression of experimental inflammatory diseases such as arthritis [50], colitis [29,51] in murine models, and human acute graft-versus-host diseases [10 –12]. Although several studies have demonstrated that fibroblast-like cells possess immunomodulatory properties in vitro [27,31 –33,35], further studies are required to determine the efficacy of different fibroblast-like populations in vivo. Clinically, dermal substitutes containing allogeneic foreskin fibroblasts have been used for the treatment of burns [36] and chronic venous ulcers [37,38] with varying success. Previous reports have demonstrated that dermal fibroblasts derived from foreskin tissue contain MSC-like populations [52] or progenitors, which display various potential to differentiate into other cell lineages such as adipocytes, osteoblasts, chondrocytes, neuronal cells, and hepatocytes [53]. Moreover, it has been recently reported that adult human dermal fibroblasts express surface proteins normally associated with BMSCs and may also possess a multidifferentiation capacity [35]. Other studies have shown that MSC-like populations derived from gingival tissue [27,29] are capable of immunosuppressive functions partly mediated by IDO in vitro and in vivo. IDO has been considered to be one of the principal immunosuppressive factors secreted by human MSC-like populations stimulated by exposure to IFN-γ produced by activated lymphocytes. IDO catalyzes tryptophan, an essential amino acid for T-cell proliferation [17,18,54]. NO produced by MSCs is also one of the major mediators of T-cell suppression by MSCs [24]. In the present study, IDO and iNOS expression by BMSCs was upregulated when cocultured with activated PBMCs or stimulated with IFN-γ, but this did not appear to be the case for HFFs treated under the same conditions. Hence, the effect of HFFs on immunosuppression may operate through a different effector pathway.
HFFs exhibited positive expression for MHC class I antigen and cell surface markers associated with MSCs (CD73, CD90, CD105, and CD166) but lacked the expression of immune costimulating factors such as MHC class II antigen, CD14, CD34, CD40, CD45, CD80, and CD86 similar to BMSCs. However, the expression of the mesenchymal precursor cell markers [42,43,55], STRO-3 and CD106, by HFFs was negligible, whereas STRO-1 expression was slightly lower than BMSCs (data not shown). Moreover, HFFs in this study lacked the capacity of osteogenic and adipogenic differentiation, suggesting the absence of MSC-like cells in this cell population.
These findings suggested that HFFs did not appear to possess the stem cell properties attributed to different MSC-like populations derived from various tissues including skin, but still displayed immunosuppressive properties by alternate mechanism(s) rather than through the actions of IDO and NO. Further, apoptosis did not appear to be the mechanism contributing to the observed reduction in activated PBMC numbers when cocultured with either HFFs or BMSCs, in accordance with previous studies [56].
Soluble factors, TGF-β1, HGF, and IL-10, secreted by BMSCs have been also demonstrated to play important roles in immunosuppressive properties [5,13,57], whereas other reports suggested that these 3 factors in BMSCs or adipose-derived MSCs are not involved in mediating immunosuppressive effects [13,49,58]. In our RT2 Profiler PCR Array assay, TGF-β1, HGF, and IL-10 expression in HFFs cocultured with Con A-stimulated PBMCs and allogeneic MLR was higher than HFFs cultured alone and BMSCs cocultured with or without activated PBMCs. However, the result of IL-10 expression in HFFs and BMSCs by real-time PCR was not consistent with the RT2 Profiler PCR Array data (data not sown). Further, recombinant HGF (1, 10, 100 ng/mL, and 1 μg/mL) failed to suppress PBMC proliferation, and neutralization assays using a functional Ab against TGF-β1 failed to block the immunosuppression mediated by CM derived from HFFs and BMSCs, although recombinant TGF-β1 showed suppression of PBMC proliferation. These findings suggest that TGF-β1 and HGF are highly expressed by HFFs cocultured with Con A-stimulated PBMCs, and allogeneic MLR did not play a pivotal role in the immunosuppression mediated by HFFs.
In the present study, a neutralizing Ab against IFN-α, which was highly expressed in HFFs cocultured with activated PBMCs, partly restored PBMC proliferation suppressed by CM derived from HFFs but failed to block the immunosuppression mediated by CM derived from BMSCs. These findings suggest that IFN-α is one of the critical immunosuppressive factors produced by HFFs. IFN-α, which belongs to type I IFNs, is a product of a multigene family of 13 functional genes that have antiviral, antiproliferative, and immunomodulatory properties [59]. The antiproliferative effect of IFN-α in malignant cell lines has been shown to mediate cell-cycle arrest in the G1 phase via the upregulation of the cyclin-dependent kinase inhibitors, p15, p21, and p27 [60 –62]. Moreover, previous studies have demonstrated that IFN-α also inhibited phytohemagglutinin (PHA)– or IL-2–stimulated T-lymphocyte proliferation by causing arrest at the G0/G1 phase and inhibiting entry into S phase through the inhibition of key cell-cycle molecules without inducing apoptosis [63,64]. In this study, we examined the cell cycling status of Con A-stimulated PBMCs cocultured with HFFs and found that HFFs had arrested PBMC division, which had peaked at G0, similar to that observed when mitogen-stimulated PBMCs were cocultured in the presence of BMSCs. Collectively, these findings suggest that the IFN-α–induced suppression of PBMCs by HFFs is due to the inhibition of cell-cycle division rather than cell death. However, it remains unclear which factors produced by activated PBMCs are critical for the upregulation of IFN-α expression by HFFs. Although not the full story, these studies lay the ground work for the identification of other soluble factors secreted by different cultured fibroblastic lines that inhibit PBMC proliferation. Importantly, the present study demonstrates that cultured fibroblastic lines derived from different sources may utilize alternate mechanisms to inhibit activated PBMC proliferation, which may be influenced by the isolation protocols and culture conditions used by different research groups. Future clinical studies may need to select specific cell populations for the treatment of inflammatory and autoimmune diseases depending on the type of disease and specific mode of action used by different cultured stromal cells. The present study implies that non–BMSC-like populations such as HFFs may serve as alternative cell sources to modulate immune responses of inflammatory diseases following allogeneic transplantation.
Footnotes
Acknowledgments
The authors acknowledge Mr. Krzysztof Mrozik and Ms. Danijela Menicanin for their excellent technical support. This work was partly funded by an NHMRC project grant (No. 453599) and an ADRF project grant (No. 63/2008).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.