
Abstract
Homing and engraftment of hematopoietic stem/progenitor cells (HSPCs) in bone marrow is the major determining factor in success of hematopoietic stem cell transplantation. This is a complex, multistep process orchestrated by the coordinated interplay between adhesion molecules, cytokines, growth factors, and regulatory cofactors, many of which remain to be defined. Recent studies have highlighted the pivotal role of unique stromal-derived factor-1 (SDF-1)/CXCR4 signaling in the regulation of HSPC homing and subsequent engraftment. In addition, studies suggest that SDF-1/CXCR4 signaling acts as an essential survival-promoting factor of transplanted HSPCs as well as maintenance of quiescent HSCs in bone marrow niche. These pleiotropic effects exerted by SDF-1/CXCR4 axis make this unique signaling initiator very promising, not only for optimal hematopoietic reconstitution but also for the development of innovative approaches to achieve restoration, regeneration, or repair of other damaged tissues potentially amendable to reversal by stem cell transplantation. This goal can only be achieved when the role of SDF-1/CXCR4 axis in hematopoietic transplantation is clearly defined. Hence, this review presents current knowledge of the mechanisms through which SDF-1/CXCR4 signaling promotes restoration of hematopoiesis by regulating the homing and engraftment of HSPCs.
Introduction
H
Under the current scenario, the enhanced efficacy of HSPCs for homing and engraftment could improve the clinical applications of HSCT and success of gene therapy. This might be achieved by modulating the ability of stem cells to home and repopulate the recipient BM. One possibility is the modification of expression and affinity of adhesion molecules on HSPCs, which are critical for homing and engraftment. Although BM homing was described as a functional concept decades ago, clear understanding of its molecular regulation remains elusive till date. Hence, a resurgence of studies delving into its mechanistic aspects is essential to define precisely its molecular mechanism and identify the critical molecular targets of HSPCs for increasing their efficiency in homing and engraftment.
Chemokines are small chemotactic cytokines (8–13 kDa) that bind to G-protein–coupled receptors (GPCR) super family and regulate crucially the directed cell trafficking during embryonic development, steady-state homeostasis, and stress conditions [11,12]. The stromal-derived factor-1 (SDF-1), also known as CXCL12, is a CXC chemokine and is considered as one of the most potent chemoattractant of HSPCs to date, eliciting its effect through binding to CXCR4 receptor on HSPCs [13,14]. The significant role of SDF-1/CXCR4 axis in the regulation of hematopoiesis has been well established by studies in knockout murine models of SDF-1 and CXCR4, displaying severely reduced B-lymphopoiesis and myelopoiesis [15 –17]. Over the past few years, a key role of SDF-1/CXCR4 signaling has been explored in the homing and engraftment of transfused HSPCs into recipient BM [18 –20]. Further, recent studies have revealed its role in HSPC survival as well as in the quiescence of HSCs for long-term maintenance [21 –23]. Considering the multiple roles in the regulation of hematopoiesis, the SDF-1/CXCR4 signaling is considered as a promising molecular target for improving HSCT success. However, although its significance is well established, information about molecular mechanism through which SDF-1/CXCR4 interaction elicits downstream effects is not precisely illustrated. Hence, an improved knowledge of the CXCR4 signaling cascade would be definitely helpful in the development of innovative strategies for enhancing the homing and engraftment efficiency to improve the success of HSCT. Therefore, we have reviewed the current knowledge of molecular regulation of HSPCs homing and engraftment with respect to specific roles of SDF-1/CXCR4 axis in the regulation of hematopoietic reconstitution.
Homing and Engraftment
Homing is the initial rate-limiting step of HSCT, wherein intravenously infused HSPCs actively cross the intact and rigid BM endothelial barrier to reach the BM extravascular space to lodge and repopulate (engraftment) within its specific niche to exert their effects in hematopoietic reconstitution [24]. Similar in principle to leucocytes recruitment from the circulation to inflammation sites [25], homing is a multistep process in which circulating HSPCs loosely adhere/roll upon BM sinusoidal endothelial cells (BMECs) followed by their firm adhesion, preventing movement back to circulation. These firmly adhered HSPCs subsequently extravasate into marrow stroma to anchor in their BM niche, a niche that possesses a unique microenvironment capable of providing not only a matrix that promotes anchorage of the HSPCs but also elicits signals that enable their proliferation and differentiation to reconstitute hematopoiesis [26,27]. Homing and engraftment are sequential steps following in the process of HSCT. Homing is a rapid process that occurs within hours and involves HSPCs adhesion, transendothelial migration, and precise placement, whereas engraftment is dominated by repopulation, proliferation, and differentiation events, which ultimately result in reconstitution of hematopoiesis [24,28].
Molecular Players in Homing: Cell Adhesion Molecules
Homing and engraftment of HSPCs in BM is regulated through a coordinated interplay of several molecular pathways. These pathways are mediated through mutual recognition of cellular adherence receptors by their cognate ligands present in the target hematopoietic microenvironment. Primitive CD34+ HSPCs express a wide range of cell adhesion molecules (CAMs), which include members of integrins (α and β heterodimers), selectins (L, P, and E), immunoglobulins (VCAMs and ICAMs), sialomucins (CD34, CD164, and others), cadherins, and specific antigens such as CD44 [29,30]. Several key molecules mediating the specific homing of human HSPCs into BM have been identified. Among these, the endothelial P and E selectins play a crucial role in the initial adherence and rolling of circulating HSPCs over BMECs under the shear flow of blood [31 –33]. Integrins, particularly VLA-4(α4β1)/VCAM-1, and LFA-1(αLβ2)/ICAM-1 pathways have revealed to be crucial for firm adhesion of rolling HSPCs over BMECs, followed by their transendothelial migration toward BM extravascular space. Recent studies have revealed the significant role of SDF-1/CXCR4 interaction in the conversion of rolling HSPCs to integrin-mediated firm adhesion over BMECs [27,31,34]. Further, HSPC transendothelial migration to the BM stroma via specific chemoattractants, especially SDF-1, has been shown in selectively directing their homing toward BM niche [35 –37]. The final anchorage of HSPCs in BM depends on the interactions of integrins, mainly VLA-4, LFA-1, and VLA-5, to stromal cell ligands VCAM-1, ICAM-1, and fibronectin, respectively [38 –40]. The marrow hematopoietic microenvironment consists of multiple types of stromal cells and an organized rich milieu of extracellular matrix. Together, these provide structural scaffolding for HSPCs to home and engraft, wherein they reconstitute hematopoiesis while maintaining the pool of quiescent stem cells through self-renewal to provide the mature blood cells throughout life [41,42].
Several studies depict a cooperative mechanism of homing pathways wherein some dominant players significantly regulate the entire process of homing and engraftment. Study on murine model indicated the VLA-4/VCAM-1 pathway as a major player of homing, wherein the administration of VLA-4 or VCAM-1 monoclonal antibodies have resulted in significant inhibition of HSPCs homing to BM, thus leading to significantly increased circulating HSPCs [43,44]. The significant contribution of SDF-1/CXCR4 signaling to BM homing became apparent when blockade of SDF-1/CXCR4 interaction was combined with deletion of VLA-4, resulting in significantly reduced HSPCs homing. In fact, while fresh steady state BM-derived hematopoietic progenitor cells (HPCs) have been shown to efficiently compensate for the loss of SDF-1/CXCR4 interaction through VLA-4/VCAM-1 pathway, the cytokine incubation of HPCs shifts the dominance among the homing molecules toward SDF-1/CXCR4 signals, wherein loss of CXCR4 has significantly reduced HPCs homing with partial compensation through VLA-4/VCAM-1 interaction and endothelial selectins. This indeed suggests the flexible dependency of homing pathways for effective homing of HSPCs in BM, wherein dominant players are repositioned with changing cytokine milieu and possibly the source of HSPCs [45]. Recently, the significant role of CD44 has been explored in homing. It has been shown that SDF-1 induces CD44 distribution to the leading edge of migrating HPCs, which in turn facilitate their transendothelial migration to BM [46,47]. Further studies are needed to define precisely the molecular links between SDF-1- and CD44-activated pathways. Taken together, these results indicate that homing and engraftment do not follow any single pathway, indeed they are a successful outcome of cooperative mechanism between several molecular pathways involving adhesion molecules, cytokines, chemokines, proteolytic enzymes, and growth factors.
SDF-1/CXCR4 Axis: An Exquisite Couple
SDF-1 was originally isolated from the murine BM stromal cell line (MS-5) and found to support the proliferation of B-cell progenitors in the presence of interleukin-7 [48]. The alternative splicing of the human SDF-1 gene results in 2 isoforms, of which SDF-1α is the predominantly expressed form of protein, consisting of 89 amino acids, and SDF-1β contains an additional 4 amino acid at carboxyl terminus. SDF-1α and β bind to CXCR4 with comparable affinity (Kd of 7.5 and 13.7 nM, respectively) and display similar physiological functions [49,50]. SDF-1 expression has been reported on stromal cells and endothelial cells of many organs including BM, skeletal muscle, liver, brain, and kidney [51]. In BM, it is mostly produced by osteoblasts in the endosteum region and highly expressed by BMECs, both of which define the niche for HSPCs [21,37].
SDF-1 acts via CXCR4, a GPCR previously known as LESTR-leukocyte–derived 7 transmembrane domain receptor [52,53]. The CXCR4 gene is expressed in 2 isoforms—“CXCR4,” the primary receptor for SDF-1, and a longer but less abundant variant “CXCR4-LO,” wherein the first 9 N (amino) terminal amino acids differ [54,55]. Similar in structure to rhodopsin, CXCR4 is a 39.7 kDa protein composed of 352 amino acids, in which 3 extracellular loop domains and the N-terminus define ligand binding communicated to 3 intracytoplasmic loops (ICLs) and cytoplasmic carboxy terminus, which activate the G-protein, mediating its influence to other downstream effectors. Its 7 transmembrane domains contain the functionally essential motifs that regulate the conformational transition/switching of CXCR4 between inactive to active stages [56 –58] (Fig. 1). CXCR4 is expressed on a variety of cells including mature blood cells (lymphocytes, monocytes, and megakaryocytes) as well as primitive CD34+ HSPCs. However, its expression on CD34+ HSPCs is differential based on its source of origin and developmental stage [59]. Significantly low level of CXCR4 surface expression has been observed on mobilized peripheral blood (MPB)-derived CD34+ cells in comparison to cord blood and BM-derived CD34+ cells, suggesting its downregulation during mobilization due to proteolytic cleavage of CXCR4 by a number of neutrophil proteases [60]. A recent study reported CXCR4 expression on endothelial and stromal cells of BM, exploring its role in SDF-1 regulation [61].
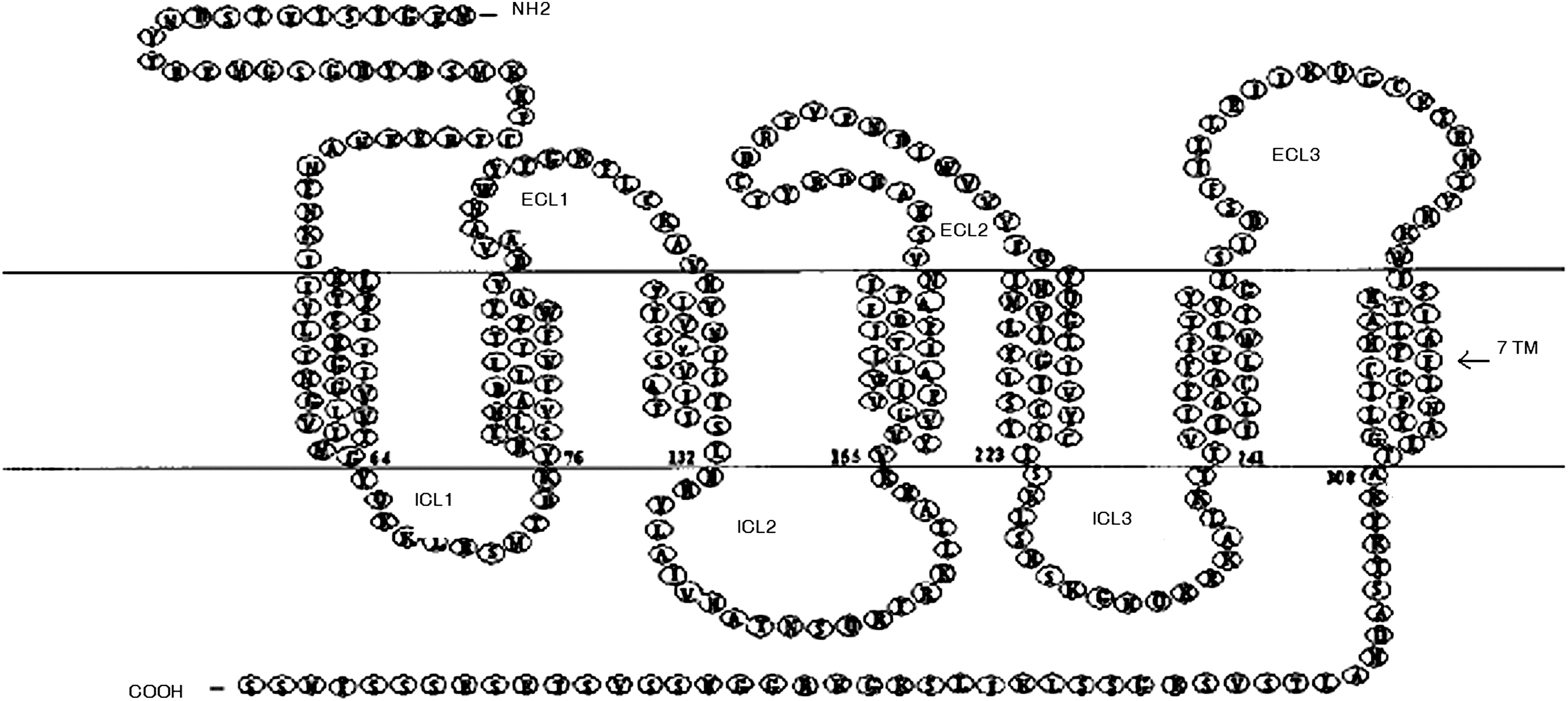
Schematic representation of CXCR4 protein depicting 3 extracellular loops (ECL), 7 transmembrane domains, and 3 intracytoplasmic loops (ICLs) along with extracellular N and intracellular C terminals.
CXCR4 and SDF-1 genes are remarkably conserved across diverse species, suggesting their key biological significance in development [62,63]. The human and murine SDF-1 differs in one amino acid only and thus it is cross reactive [49], providing an opportunity to conduct in vivo studies of human SDF-1/CXCR4 signaling in immune-compromised recipient murine models. Recently, the CXCR7, a new receptor of SDF-1, has been identified on T cells, activated endothelial cells, and many tumor cell lines [64,65]. However, recent study has shown that it is scarcely expressed on the surface of human T lymphocytes and CD34+ progenitor and concentrates mainly intracellular. Although CXCR7 is unable to support on its own any SDF-1–induced integrin activation or cell migration, a study done using specific CXCR7 blockers has indicated a crosstalk between intracellular CXCR7 and CXCR4, which is involved in rapid SDF-1–induced integrin activation but not in chemotaxis of human T lymphocytes and CD34+ cells [66]. Further study in this direction will clearly identify other anticipatory pathways in homing and engraftment along with SDF-1/CXCR4 axis.
SDF-1/CXCR4 Signaling: A Key Regulator of Homing and Engraftment
Decades of research has established the current dogma of SDF-1/CXCR4 signaling, implicating its significant role in angiogenesis, neuronal development, immune cell trafficking, and hematopoiesis. Gene knockout studies of SDF-1 and CXCR4 in murine model have displayed identical phenotype with multiple early embryonic lethal defects including lack of BM seeding by HSPCs as well as impaired B-cell lymphopoiesis and myelopoiesis in BM [15 –17]. These results demonstrate an essential requirement of SDF-1/CXCR4 axis in homing of fetal liver-derived HSPCs to BM and hematopoietic retention and repopulation of this organ during embryonic development.
Further studies in this direction highlighted the pivotal role of SDF-1/CXCR4 signaling in adult BM homing and engraftment of transfused human HSPCs by mediating their chemotaxis to BM. Blocking of CXCR4 surface expression on human CD34+ cells by CXCR4 antibody or its downregulation through SDF-1 desensitization has resulted in their reduced chemotaxis toward SDF-1 gradient in vitro as well as significantly reduced engraftment in immune-compromised recipient mice. In contrast, upregulation of CXCR4 expression through short-term in vitro cytokines pretreatment with stem cell factor, interleukin-6, or hepatocyte growth factor has resulted in significantly increased SDF-1–induced chemotaxis in vitro as well as their in vivo homing in immune-compromised mice [18 –20]. Consistent with these results, CXCR4 overexpression on human cord blood and MPB-derived CD34+ cells has resulted in significantly enhanced chemotaxis toward SDF-1 in vitro as well as their enhanced BM engraftment in NOD/SCID recipient mouse [67 –69]. Similarly, retroviral-mediated CXCR4 overexpression on M07e cells (an established model of human HPCs) resulted in significantly increased SDF-1 chemotaxis and cell survival [70]. Taken together, these results delineate the significant role of SDF-1/CXCR4 axis in the homing and engraftment of HSPCs to BM.
Recent studies provide new insight into the molecular regulation of homing and engraftment mediated by SDF-1/CXCR4 interaction. SDF-1 expressed on BMECs binds to CXCR4 of CD34+ cells, inducing the G-protein–mediated downstream signaling cascade that is found to be crucial for increasing the adhesiveness of key integrins VLA-4, VLA-5, and LFA-1 of CD34+ cells to their endothelial ligands, VCAM-1 and ICAM-1. This in turn translates initial rolling of CD34+ cells into firm shear-resistant arrest on vascular endothelium under blood flow [31,34]. Further, SDF-1/CXCR4 signaling induces a transient increase in intracellular calcium level and rho GTPase-mediated cytoskeleton actin polymerization [71,72]. Combined activities of all these molecular events trigger integrin-mediated cell polarization of HSPCs and their subsequent transendothelial migration toward SDF-1 gradient in BM stroma (Fig. 2). In addition, HSPCs interaction with stromal protein, mainly fibronectin through VLA-4 and VLA-5 receptors, has also been shown to be enhanced in vitro by SDF-1, indicating the role in HSPCs anchorage to BM stroma [31,34]. Recent studies have revealed the role of small GTPase-Rac and Rho as critical downstream effectors of CXCR4 signaling. Rac1 colocalization with CXCR4 in lipid rafts is suggested to potentiate human BM CD34+ cell migration and repopulation, whereas the knocking out of Rac-1 significantly reduced the SDF-1–induced migration and in vivo homing of murine stem cells to BM [24,73,74]. A study done in murine model, wherein CD34+ cells highly express CXCR4 but were deleted of both Rac-1 and Rac-2 alleles, showed their inability to migrate toward SDF-1 gradient in BM, thus leading to their increased mobilization in peripheral circulation [75]. These results clearly delineate the significant role of Rac and Rho in SDF-1/CXCR4-mediated homing and engraftment. A recent study has shown that overexpression of sphingosine-1-phosphate (S1P) receptor-S1P(1) in MPB-derived CD34+ cells stimulated the S1P-dependent chemotaxis of these cells but significantly inhibited their SDF-1/CXCR4-dependent migration and in vivo BM homing in sublethally irradiated NOD/SCID mice. These results not only support the significant role of SDF-1/CXCR4 signaling in BM homing but also indicated the existence of its antagonistic molecular crosstalk with S1P(1) receptor [76].
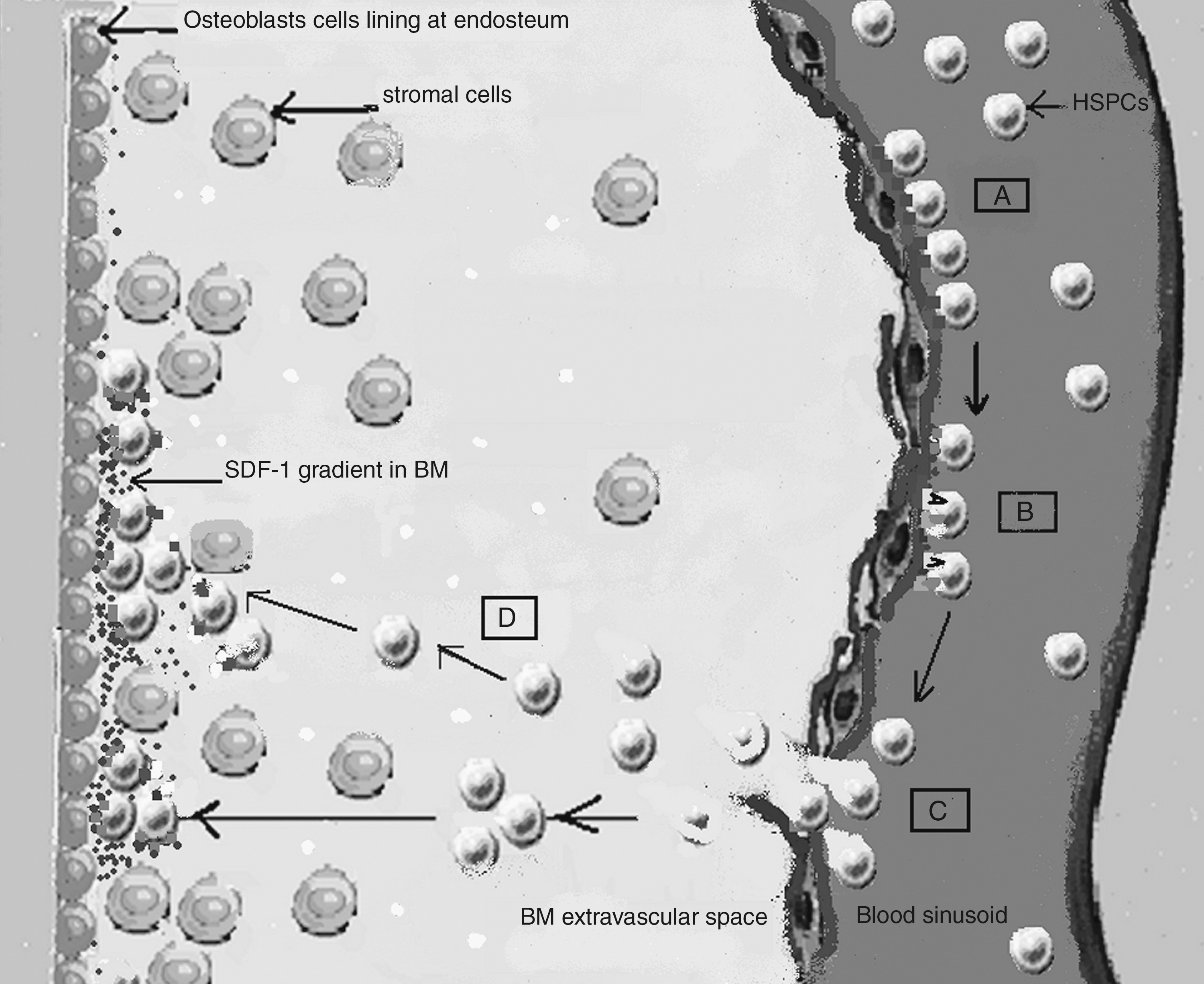
Role of SDF-1/CXCR4 interaction in homing of HSPCs. (A) Initial tethering of HSPCs on BM endothelial cells. (B) SDF-1/CXCR4 interaction results in integrin-mediated firm adhesion of rolling HSPCs. (C) Transendothelial migration of CXCR4 + HSPCs toward SDF-1 gradient in BM stroma. (D) Transmigration of HSPCs into BM stroma to reach their specific niche for lodgment. Molecule coding: black square, selectins; gray square, CXCR4; gray circle, SDF-1; black V, VLA-4; white square, VCAM-1; dark gray rectangle, LFA-1; light gray rectangle, ICAM-1; white circle, fibronectin; black small square, VLA-5.
Despite these results strongly support the implication of SDF-1/CXCR4 signaling in homing and engraftment, it was unexpectedly found that CXCR4 null HSCs isolated from the fetal liver of CXCR4 knockout embryos were able to home BM of immune-compromised wild-type recipient adult mice preconditioned with total body irradiation (TBI). However, the stem cell self-renewal, retention of HSPCs within BM, and migration patterns of both myeloid and lymphoid progenitor cells were found to be defective in these recipient mice [77,78]. Indeed, it is noteworthy that transplantation of CXCR4 null cells recovered from primary recipients into secondary recipient mice resulted in significantly reduced engraftment [77]. Although these results delineate the essential role of SDF-1/CXCR4 interactions in long-term stem cell repopulation and engraftment, the mechanistic basis of homing of these cells to normal murine BM is currently poorly understood. Therefore, further research is necessary to be undertaken on SDF-1/CXCR4 signaling, especially the functional significance of CXCR4 expression on BM endothelium and stromal cells in the directed trafficking of HSPCs. Interestingly, it has been found that CXCR4 expressed on blood platelets can be functionally transferred to CD34+ cells, which in turn augment their SDF-1–mediated homing and engraftment [79,80]. These results suggest the possible role of CXCR4 + blood platelets in wild-type recipient mice for homing of CXCR4 null cells. Also, this is suggestive of other alternative mechanisms of homing that may compensate the lack of CXCR4 in these null cells. However, if such mechanisms exist, they appear to be secondary in the given context where CXCR4 null cells did not qualify as definitive repopulating stem cells capable of repopulating and reestablishing hematopoiesis in recipient BM [77,78]. This notion is further strengthened by a recent study that demonstrated the inability of CXCR4-deficient HPCs to home and engraft into BM of lethally irradiated adult recipient mice; however CXCR4 deletion in these adult mice did not impair the HSC retention in BM [81]. Overall, this suggests further investigation into HSPCs homing and engraftment, not only for proper clarification of any ambiguity but also to explore the hidden role of SDF-1/CXCR4 signaling in the reconstitution of hematopoiesis following HSCT.
Beyond Homing: Role in Survival and Maintenance of HSCs in BM
Apart from directed migration and homing, the SDF-1/CXCR4 signaling has also revealed a role in HSPC survival and repopulation, which ultimately facilitates engraftment of transplanted HSPCs into recipient BM. The myeloid progenitors from SDF-1 transgenic mice (endogenously expressing SDF-1 under a Rous sarcoma virus promoter) have shown enhanced survival and antiapoptotic potential in vitro relative to similar cells from wild-type mice. Further, progenitors from these SDF-1 transgenic mice displayed enhanced BM and spleenic myelopoiesis [82]. The antiapoptotic/survival-promoting role of SDF-1 was further substantiated by a study demonstrating that low concentrations (0.05–0.5 ng/mL) of exogenous SDF-1 alone promote survival of purified CD34+ cells in vitro [83]. Such survival-promoting activity has also been confirmed in vivo, wherein, in combination with other cytokines, SDF-1 increases the long-term survival of lethally irradiated mice [84]. Supporting this notion, retroviral-mediated CXCR4 overexpression in M07e cells have resulted in significantly increased cell survival [70]. In apparent agreement with these studies, SDF-1/CXCR4 signaling has been shown to promote HSPC proliferation under certain experimental conditions. Low concentration of exogenous SDF-1 triggers PB-derived CD34+ cells from quiescent G0 to G1 phase of cell cycle and, in combination with cytokines, act as a priming factor, making HSPCs more responsive to the effect of cytokines, which induce proliferation [23,83,85].
Although low concentrations of SDF-1 in synergy with early-acting cytokines enhance CD34+ cell proliferation, its high levels have been shown to maintain the HSC pool in BM via inducing the quiescence of human long-term culture initiating cells [86,87]. A study demonstrated the significant reduction of HSCs in BM of CXCR4 conditionally deficient adult mice when compared with wild-type mice. In addition, cell cycle progression of HPCs from G0 to G1 was substantially promoted in these mice compared with wild-type mice, in which most HPCs remained quiescent in G0 phase
WHIM syndrome is a congenital immune deficiency characterized by warts, hypogammaglobulinemia, recurrent bacterial infection, and myelokathexis [104 –107]. Many but not all cases of WHIM syndrome have been linked to autosomal dominant mutations in CXCR4, all of which cause truncations of the carboxy-terminus of CXCR4 leading to failure of mutant CXCR4 to downregulate, resulting in prolonged activation [108]. One of the prominent feature of WHIM is myelokathexis, which is clinically manifested by severe chronic neutropenia with marrow hyperplasia and inappropriate apoptosis of mature myeloid cells in the BM. To better explore the mechanism of myelokathexis, a study demonstrated that retroviral-induced transgene expression of mutated CXCR4 (WHIM variant) in human CD34+ cells resulted in significantly higher cell migration, calcium flux function, and in vivo engraftment in NOD/SCID recipient mice when compared with the expression of excess wt CXCR4. Moreover, mutated CXCR4 showed a significant increase in apoptosis of transduced cells in BM and reduced release of transduced leukocytes into peripheral blood of recipient mice, whereas the in vitro culture of these cells did not show any evidence of increased apoptosis. This suggests that neutropenia and increased apoptosis of mature myeloid cells in BM of WHIM patients is indeed secondary to a failure of marrow release and progression to normal myeloid cell senescence [109]. These studies provide a mechanism and rationale for WHIM treatment and suggest that AMD3100 might be efficacious to mobilize the neutrophils that are abnormally retained in the BM.
Dynamic Expression of the SDF-1/CXCR4 Axis: Impacts on Homing and Engraftment
The SDF-1/CXCR4 axis is regulated dynamically via their expression levels, which are tightly controlled by various factors or mechanisms. In BM, SDF-1 is predominantly expressed by endothelial cells and endosteal bone lining osteoblasts [37]. Moreover, in mouse studies, SDF-1 mRNA has been shown to be most highly expressed by osteoblasts in comparison to lower levels by endothelial cells in BM [110]. It has been shown that SDF-1 is immobilized on BMEC surface through binding to glucose aminoglycans [71,111 –113], protecting it from protease cleavage and thus increasing its local concentration gradient. This ultimately enables SDF-1 to be presented to migrating HSPCs with enhanced chemotactic activity contributing to increased homing [114]. This indicates that high concentration of SDF-1 protein in BM is likely due to its endogenous expression by these cells as well as capture through binding to glucose aminoglycans. Indeed, this could probably explain the specific SDF-1–induced HSPC engraftment in BM, whereas this chemokine is markedly expressed by a variety of tissues. Moreover, live in vivo microscopic study in murine calvaria has shown that SDF-1 protein is not homogenously present on all BMECs but primarily on sinusoidal BM vasculature, which being a prospective HSC niche is very relevant to BM function as well as HSPC engraftment [115].
DNA-damaging agents (irradiation or cytotoxic drugs) have been shown to increase mRNA as well as protein level of SDF-1 in BM, which in turn augments the homing and engraftment of transplanted HSPCs following irradiation of recipient [37]. The SDF-1/CXCR4 axis is negatively regulated by several proteases present in the BM hematopoietic microenvironment. Chemotactic inactivation of SDF-1 due to its N-terminal cleavage has been demonstrated by neutrophil elastase, cathepsin G, cathepsin K [60,93,116,117], matrix metalloproteinase-2/9 [118], and C-terminal cleavage by carboxypeptidase N [119]. In addition, neutrophil elastase and cathepsin G have shown to cleave CXCR4 on human T cells and CD34+ cells [60,120]. It has been shown that GCSF administration for stem cell mobilization actually induces increased BM concentration of proteases such as neutrophil elastase, cathepsin G, and matrix metalloproteinase-9, all of which enzymatically degrade SDF-1–mediated HSPCs homing to BM, ultimately resulting in HSPC mobilization to peripheral blood [93,121]. Similarly, the cell surface peptidase-CD26 expressed on CD34+ cells negatively regulates the homing and engraftment by causing proteolytic degradation of SDF-1 and thus its inhibition in CD34+ has resulted in increased efficiency of HSPCs homing and engraftment [122].
TGF-β1 in BM has been shown to down-regulate SDF-1 expression, both at mRNA and protein level, which in turn control CD34+ cell migration and homing to BM [123]. Further, study has revealed TGF-β1 as a modulator of long-term responsiveness of HSPCs to SDF-1. It has been shown that in the presence of low level of TGF-β1, CD34+ cells preexposed to SDF-1 remain chemotactically responsive to SDF-1 for a prolonged period of time and retain sustained adhesion response mediated by SDF-1. In fact, this might explain as one of the possible reasons for the responsiveness of HSPCs to constant exposure of SDF-1 in BM [124]. A recent study indicated the CXCR4-mediated transcytosis of circulating SDF-1, wherein circulating SDF-1 bind to CXCR4 expressed on endothelial and stromal cells of BM, followed by SDF-1/CXCR4 complex internalization and subsequent active translocation of SDF-1 to BM stroma, a mechanism termed as “transcytosis” [61].
CXCR4 expression is highly dynamic and regulated by both autocrine and paracrine mechanisms. It has been shown that hypoxia upregulate the gene expression of SDF-1 [125] and CXCR4 by causing the stabilization of hypoxia-inducible factor-1α, subsequently leading to the transcriptional activation of CXCR4 gene. Indeed, hypoxia further contributes to increased CXCR4 gene expression through stabilization of CXCR4 transcripts, likely by hypoxia-regulated RNA-binding factors, which may interact with and stabilize the CXCR4 mRNA at posttranscriptional level [126]. Supporting this notion, incubation of human BM-derived CD34+/CD38− cells in hypoxic conditions resulted in hypoxia-inducible factor-1α stabilization and significantly increased in vitro survival and repopulation as well as their in vivo homing and engraftment in immune-compromised recipient mice [127]. A recent study revealed that histone deacetylase inhibitors such as valproic acid increase CXCR4 expression on HSPCs by chromatin remodeling [128]. CXCR4 functionality crucially depends on its incorporation into lipid rafts. Several priming agents, such as C3a, hyaluronic acid, and fibrinogen, prime the homing-related cellular responses of HSPCs to SDF-1 gradient by increasing the CXCR4 incorporation into lipid rafts [129,130]. Apart from ligand-induced endocytosis [131], a study revealed the ligand-independent constitutive endocytosis of CXCR4 in CD34+ cells, which accounts for its low surface expression, while a large part of protein is sequestered intracellularly. The reasons for this phenomenon are currently unknown, but several possible explanations have been envisioned. As SDF-1 is produced at a high level within BM, it might be necessary to tightly regulate CXCR4 surface expression on CD34+ HSPCs. Alternatively, the intracellular pool of CXCR4 may serve as a reservoir, allowing for a rapid response to SDF-1 [114,132]. Overall, a complex picture that regulates the activity of SDF-1/CXCR4 axis is emerging (Fig. 3). Hence, a better understanding of SDF-1 and CXCR4 trafficking in BM will be helpful to understand the molecular regulation of homing, engraftment, and hematopoiesis.
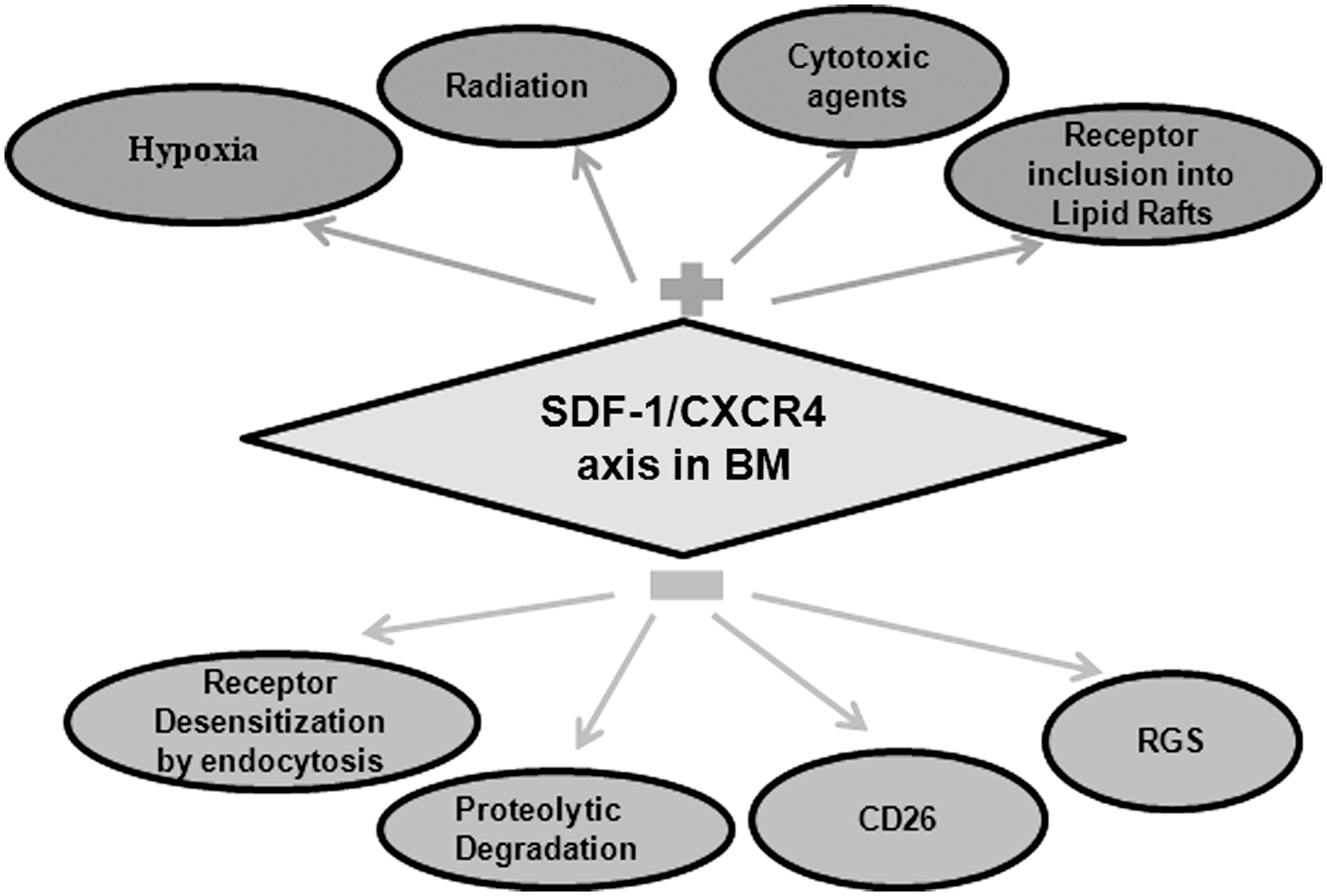
SDF-1/CXCR4 regulation in BM. Upregulation: (1) radiation, cytotoxic agents, and hypoxia upregulate SDF-1 and CXCR4 expression; (2) CXCR4 inclusion into lipid rafts increases its functionality. Downregulation: (1) proteolytic degradation of SDF-1/CXCR4 by BM proteases and CD26; (2) CXCR4 endocytosis downregulates its surface expression; (3) regulators of G-protein signaling (RGS) negatively regulate CXCR4 signaling.
The SDF-1/CXCR4 Signaling Cascade
Although our understanding of significant roles played by SDF-1/CXCR4 axis in the regulation of hematopoiesis has improved substantially in recent years, relatively little is known about its downstream signal targets. Site-directed mutation studies of CXCR4 revealed that its N-terminus and extracellular loop domains contain critical binding sites for SDF-1 [56,133]. Posttranslational sulfation of its amino-terminal tyrosine21 and glycosylation at asparagine11 are found to be critical for SDF-1 binding [134,135]. SDF-1 binding triggers a domain-specific conformational change in CXCR4, switching it from an inactive to active state, and as a result, its cytoplasmic loops and C-terminal end of CXCR4 are made available for binding to Gαi and other downstream effectors. Mutational studies on GPCR family members have revealed that it is primarily the intermolecular interaction between conserved motifs in TM 3, 6, and 7 that regulates the conformational isomerism of the receptor [58,136]. Although several studies have explored the activation of specific downstream signaling mediators of CXCR4, how these pathways together elicit the biological effects are still complex. What is known is that SDF-1 binding to CXCR4 triggers G protein coupling to ICL3 domain and subsequent dissociation of the heterotrimeric G protein into Gβγ and Gαi subunits, which in turn bind to several downstream effectors, resulting in activation of PI3K, protein kinase C (PKC), and MAPK-mediated pathways [57,137,138].
Studies in a model hematopoietic progenitor cell line (CTS) and human CD34+ HSPCs have revealed that SDF-1–triggered CXCR4 downstream signaling induces the transient intracellular Ca2+ flux [139] and increases tyrosine phosphorylation of several focal adhesion components, including the proline-rich tyrosine kinase 2 (Pyk-2), RAFTK, focal adhesion kinase, adaptor molecules Crk, Crk-L, and p130 Cas, and the cytoskeleton protein paxillin. Further, using specific inhibitors, both PKC and PI3K activation has been found to be essential for SDF-1–mediated phosphorylation of these focal adhesion proteins and cell migration, whereas the mitogen-activated protein kinases ERK-1 and 2 were not. These findings delineated the functional roles of PKC- and PI3K-dependent downstream effectors, connecting downstream signaling of CXCR4 to HSPCs trafficking and adhesion [140]. PKCzeta (PKCζ), an atypical PKC isoform, has been reported to be a crucial downstream component of the CXCR4 signaling. Interestingly, PKCζ inhibitors prevent CD34+ cell migration toward SDF-1 gradient in vitro as well as BM repopulation, but homing was unaffected, suggesting its crucial role in CXCR4-mediated engraftment rather than homing. Moreover, PI3K has been identified as a PKCζ activator, whereas Pyk-2 and ERK1/2 have been identified as downstream targets of PKCζ in the CXCR4 signaling cascade [141].
Although the role of Rac-1 and Rac-2 as crucial downstream targets of CXCR4 signaling in the regulation of homing and engraftment has been established [73 –75], how and where they act in the signaling network has not been defined precisely. Small Rho GTPase inhibitors have been shown to suppress the SDF-1–induced Ca2+ influx in HPCs, an essential molecular event for chemotaxis [142]. Interestingly, the Gβγ subunit of Gi protein coupled to CXCR4 is involved in SDF-1–induced Rac activation and cell migration, whereas CXCR4 can also stimulate Rho in a Gi-independent manner through the α subunit of G13, leading to Rho kinase-mediated phosphorylation of myosin light chain. Hence, coordinated activation of both Rac and Rho are required for CXCR4-induced cell chemotaxis [73].
Although ICL3 is identified to be the only cytoplasmic loop of CXCR4 that binds Gi protein, SDF-1–induced cell chemotaxis seems to be more complex, as it is dependent on ICL2, ICL1, and the C-terminus of CXCR4 along with ICL3 [57]. Indeed, this indicates that CXCR4 regulates HSPCs homing and engraftment not only through G-protein–dependent pathways, but also through G-protein–independent downstream signaling. Supporting this notion, studies on hematopoietic progenitor cell lines reported that SDF-1 stimulates tyrosine phosphorylation of Janus kinase 2 (JAK2) and other members of the JAK signal transduction, including JAK1, tyrosine kinase 2, STAT2, and STAT4, all in a G-protein–independent manner. Nevertheless, studies using a JAK2 inhibitor revealed JAK2 to be an upstream target of PI-3K, ultimately resulting in tyrosine phosphorylation of multiple focal adhesion proteins. Inhibition of JAK2 has been shown to decrease the SDF-1–induced cell migration in both CTS and CD34+ progenitor cells. These data provide new insight into the role of JAK kinases in CXCR4-mediated cell chemotaxis. However, the molecular mechanism that mediates the interaction between JAK proteins and CXCR4 needs to be further delineated [143,144]. Taken together, these studies illustrate the complexity of the SDF-1/CXCR4 downstream signaling cascade (Fig. 4) that regulate homing and engraftment (Fig. 5). Further studies will be necessary to decode the complex and interwoven signaling network completely.
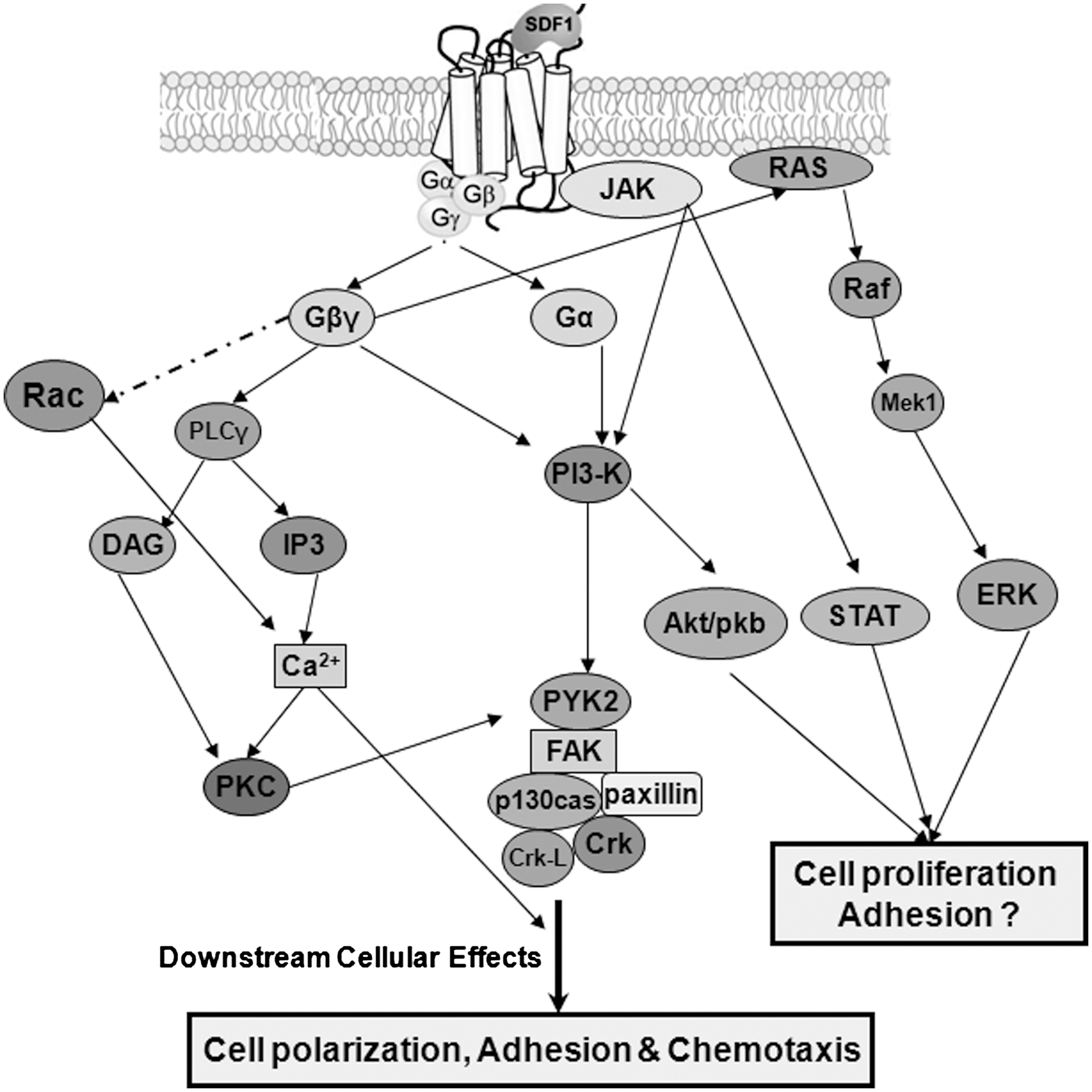
SDF-1/CXCR4 signaling cascade. SDF-1 binding to CXCR4 triggers Gi protein-mediated downstream signaling and PI3K/Akt, PKA, PKC, RAS/MAPK, as well as Gi-independent JAK/STAT activation. Altogether, these regulate the cell adhesion, locomotion, and chemotaxis of HSPCs to home BM. However, its direct effect on cell proliferation remains controversial to date. PI3K, phosphoinositide-3 kinase; PKC, protein kinase C.
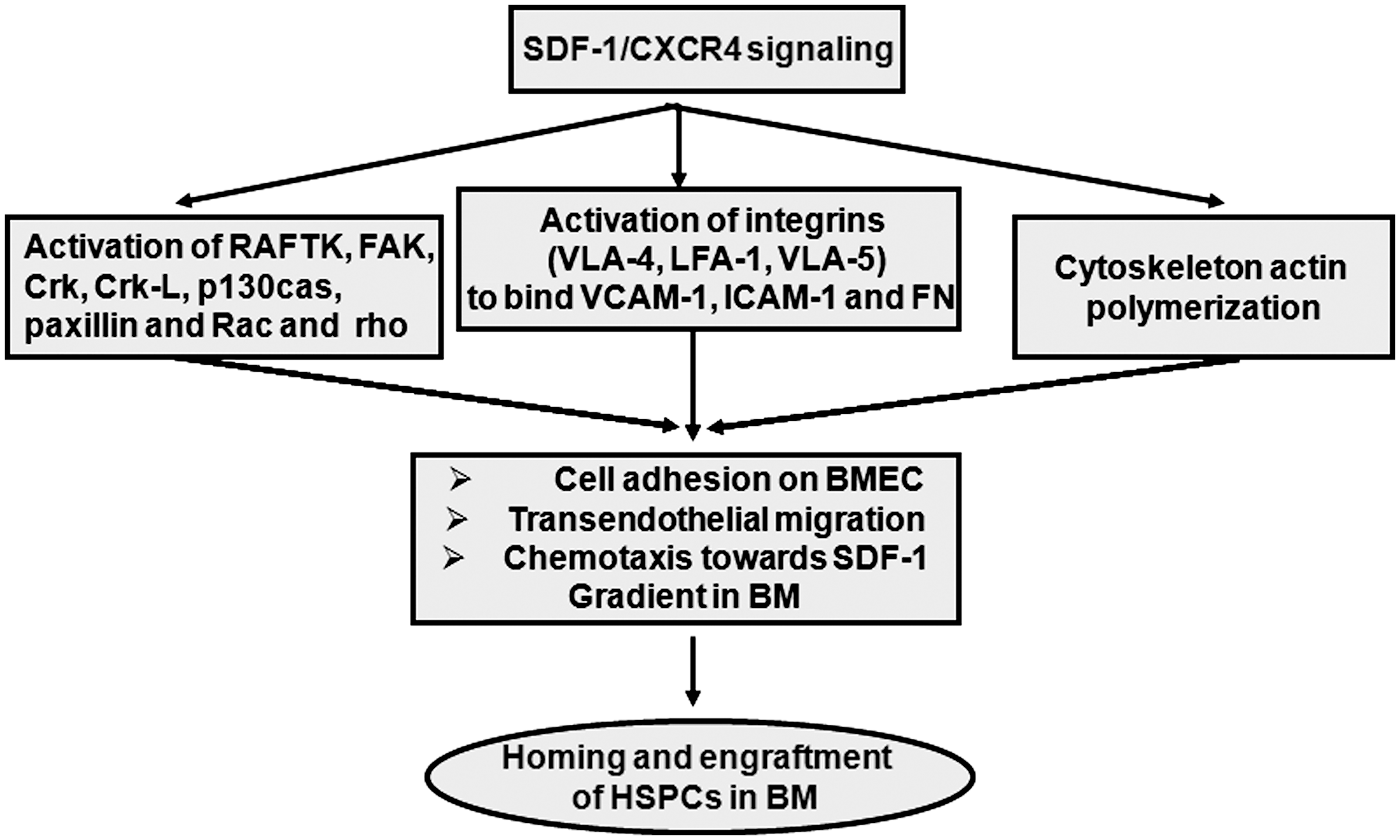
Downstream mechanistic effects of SDF-1/CXCR4 signaling in the regulation of homing and engraftment of HSPCs. The SDF-1/CXCR4 interaction induces downstream molecular events, activation of integrins, focal adhesions, and actin polymerization, which altogether elicit different cellular responses of HSPCs (cell adhesion transendothelial migration and chemotaxis), essentially implicated in homing and engraftment.
Conclusion and Future Perspectives
This review highlights recent findings that emphasize the newly appreciated role of SDF-1/CXCR4 signaling as a principal regulator of homing and engraftment. However, several aspects are still ambiguous and need to be addressed further. Much of the current research on transendothelial migration and homing of HSPCs in BM is focused on CXCR4 expressed on HSPCs and its ligand SDF-1 expressed by endothelial and stromal cells of BM. Studies looking from the other direction, that is, examining the functional significance of CXCR4 expression on stromal and endothelial cells of recipient BM, in the homing and engraftment of transfused HSPCs need to be examined further.
Additionally, studies indicating the role of SDF-1/CXCR4 signaling in HSPC survival and proliferation leave us with a quandary of conflicting observations. Whether SDF-1/CXCR4 signaling is directly involved in cell cycle progression and survival or indirectly regulates by inducing cell adhesion remains a source of contention. Similarly, while the role of SDF-1/CXCR4 signaling in HSCs quiescence has been recently explored, the molecular mechanism remains unclear. Hence, a wide spectrum of multidirectional study addressing these issues is certainly needed to establish a notion without any ambiguity as well as to explore its hidden roles in regulation of hematopoiesis.
In conclusion, the SDF-1/CXCR4 signaling cascade acts as a key player in the journey of intravenously infused HSPCs, influencing their homing and engraftment to reestablish hematopoiesis in recipient BM. However, its functional relationships to homing, engraftment, as well as HSC survival, quiescence, and retention in BM have yet to be fully understood. We are at the beginning of a long journey and have miles to go before we understand the complex molecular regulation involved. Certainly new strategies are bound to emerge, not only to increase engraftment efficiency in recipient through upregulation of SDF-1/CXCR4 signaling cascade but also to increase HSPC mobilization in donor peripheral blood by disruption of this axis. This will ultimately contribute to greater clinical success of HSCT and gene therapy.
Footnotes
Acknowledgments
The authors thank the director of the Institute of Nuclear Medicine and Allied Sciences (INMAS-DRDO) for his encouragement and support. M. Sharma thanks Defense Research and Development Organization (DRDO), Delhi (India), for the award of Ph.D. research fellowship.
Author Disclosure Statement
No competing financial interests exist.