
Abstract
Mesenchymal stem cells (MSCs) are a very important adult stem cell population with a multitude of potential applications in regenerative medicine. The thorough characterization of the bone marrow MSC (BM-MSC) population derived from the BALB/c species was essential, considering the significance of the murine model amongst animal models. In the present study, we examined the effect of gender, age, and in vitro culture on the basic properties (proliferation, differentiation, and immunosuppressive potential) of BM-MSCs. We found a decline in the progenitor frequencies from the BM of adult mice, lower MSC frequencies in all female donors, and an increase in the BM-MSC proliferation rate upon in vitro propagation. We also examined BM-MSCs for the expression of the 3 major embryonic stem cell transcription factors, Oct3/4, Sox-2, and Nanog, as well as 2 mRNA binding proteins, coding region determinant binding protein/insulin-like growth factor 2 mRNA binding protein 1 (Crd-bp/Imp1) and Deleted in azoospermia-like (Dazl), which are expressed in primitive stem cells, umbilical cord blood-hematopoietic stem cells and amniotic fluid stem cells, respectively. Further, it has been reported that these 2 genes are critical for embryonic development. In this study, therefore, we report, for the first time, the expression of Crd-bp/Imp1 and Dazl in BM-MSCs. Dazl, Oct3/4, and Sox2 were detected in relatively low levels in contrast to Crd-bp/Imp1, its major target c-Myc, as well as Nanog, which were expressed redundantly, irrespective of sex, donor age, or in vitro passaging. These findings could further support the extrinsic theory of aging of the MSC population and the potential implication of embryonic genes in adult stem cell physiology.
Introduction
M
MSCs are self-renewing entities while generating multipotential progeny. Hence, this class of cells is of paramount importance for an organism during infancy and adolescence as well as for cellular homeostasis during adulthood [8]. However, controversy exists on whether these cells are either subject to, or causes of aging. Studies regarding hMSCs suggest that their frequency in the BM either declines with age progression or remains constant. Reports also state that there is an age-related deterioration of MSC physiology [9 –11]. A few sporadic, similar studies exist for mMSCs, and thus, a more thorough investigation of their properties is needed [12 –14].
So far, a single universal stem/progenitor marker to define MSCs does not exist; hence, a combination of characteristics, phenotypic and functional, is used to describe them [15]. In an effort to more precisely portray MSCs, the expression of various embryonic markers has been investigated. Recently, hMSCs were found to express embryonic stem cell (ESC) markers such as OCT3/4, NANOG, SOX-2, alkaline phosphatase, and stage-specific embryonic antigen 4 (SSEA-4) [16]. In the mouse BM, ESC markers were found to be expressed only by more primitive mMSCs such as multipotent adult progenitor cells, very small embryonic-like cells, and nonhematopoietic/endothelial SSEA-1+ cells [15,17 –19]. At present, the role of ESC markers in ASCs has not been clarified, and sometimes, controversial results are reported, for example, for Oct3/4. Lengner et al. recently rendered Oct3/4 dispensable for both self-renewal and maintenance of somatic stem cells in the ASC compartments of several tissues, including the intestinal epithelium, BM (hematopoietic and mesenchymal lineages), hair follicle, brain, and liver [20]. Undoubtedly, additional studies must be conducted to further confirm the role of ESC markers in ASC.
A protein expressed during mouse embryogenesis [21], in ESC and other stem cell types such as amniotic fluid stem cells, as previously described by Stefanidis et al., is Deleted azoospermia-like gene (Dazl) [21,22]. The exact role of Dazl has not been completely deciphered, but it is an important transcriptional regulator in germ cell development and could possibly play a role in pluripotency [23,24].
Our group has previously detected the expression of an embryonic protein, the insulin-like growth factor 2 mRNA binding protein 1 (CRD-BP/IMP-1), in umbilical cord blood (UCB) HSCs, but not in BM or mobilized blood CD34+ HSCs [25]. In the embryo, Crd-bp/Imp1 plays an important role—as exhibited by high perinatal mortality of knockout mice—particularly in intestinal morphogenesis via regulation of extracellular matrix formation [26]. Moreover, CRD-BP/IMP-1 expression in primitive HSCs suggests a normal role in the biology of stem cells with high renewal capacity. The major identified targets of Crd-bp/Imp1 are c-Myc, IgfII, h19, and tau mRNA, all known to be implicated in developmental processes [27].
Mice are very often used as model organisms in preclinical studies to test putative human therapeutic treatments. In this context, molecular and cellular changes, which alter MSC properties with age, are of particular interest in translational research, especially when elderly patients are the target group for such cell-based therapies. Therefore, in this study, we sought to evaluate the effects of both in vivo age and in vitro cellular aging on mMSCs initially with regard to their major phenotypic and functional properties and, second, on the expression of genes with an embryonic pattern of expression (Oct3/4, Sox2, Nanog, Dazl, and Crd-bp/Imp1). Although these genes have been detected in more primitive stem cell types, their expression in mMSCs would be of great interest as it could unravel new aspects of MSC biology.
Materials and Methods
Cell isolation
mMSC cultures were established from the BM of male and female BALB/c mice of different ages, according to a modified protocol of Peister et al. [28]. In this study, 1 mouse represents 1 MSC culture; at least 3 cultures were established for each experimental group. In total, 20 MSC cultures were used in the analysis of these results. In brief, mice were sacrificed and the BM of each tibia and femur was flushed using RPMI 1640 (Invitrogen) supplemented with 20% FBS (Biosera), 2 mM
hMSCs, isolated from the BM of healthy individuals, were cultured and characterized as previously described [29]. Passages 3–5 of hMSCs were used in gene expression studies.
Cell proliferation assays
Colony forming units-fibroblast assay
This assay was utilized at 2 different stages of mMSC culture: first, in freshly isolated BM mononuclear cells (BMMNCs), to determine MSC frequency. At this point, 105 BMMNCs/cm2 were seeded onto a 6-well plate (surface area: 10 cm2/well) and cultures were maintained as described earlier. Second, to confirm the self-renewing capacity of mMSCs during culture, 10 cells of mMSCs per cm2 were seeded at different passages onto 6-well plates in complete medium. At day 14, in both experimental settings, the medium was removed; the cells were washed twice with phosphate-buffered saline (PBS) and stained with 0.5% Crystal Violet (Merck & Co.) in methanol. Colonies larger than 50 cells were counted under a stereoscopic microscope. All experiments were performed in triplicates.
Calculation of cumulative growth index
The growth index (GI) was calculated using the following equation: GI = N t /N0 , where N 0 is the inoculum cell number and N t is the cell harvest number. The cumulative GI (cGI) was calculated using the following equation: cGI = GI n × GI (n − 1) , where n is the passage number.
Calculation of doubling time
The population doubling time (T D ) was calculated using the following equation: T D = ln2 × Δt/(lnN t − lnN 0), where Δt is the time for which the culture was allowed to grow [30].
Population doubling
The population doubling (PD) was calculated according to Cristofalo et al. by the following equation: PD = (logN t − logN 0)/log2 [31].
Surface staining
Upon trypsinization, MSCs were washed in fluorescence-activated cell sorting (FACS) buffer (PBS, 2% FCS, 0.1% NaN3) and stained as previously described [29] for 15 min at room temperature. Cells were fixed in 1% paraformaldehyde (PFA). Flow cytometric analysis was performed on an FACSCalibur Flow Cytometer (BD Biosciences) using CellQuest software. The anti-mouse antibodies used were CD29-FITC (fluorescein isothiocyanate), CD73-PE (phycoerythrin), Sca-1 PE, H-2Dd-PE, I-A/I-E-PE, CD45-PerCP (peridinin chlorophyll-α protein), CD11b-APC (allophycocyanin), and CD44-APC (all from BD Biosciences) and CD105-PE (R&D Systems).
In vitro differentiation
In vitro adipogenesis assays were conducted according to Meirelles Lda and Nardi [32] and osteogenesis according to Peister et al. [28,32]. In brief, mMSCs were plated to confluence in 10% a-MEM and the following day the medium was replaced with an equal volume of differentiation or control medium. Cells underwent differentiation for 2 weeks. Media were replaced twice a week. To confirm their differentiation status, cells were stained with Oil Red O and Alizarin Red S, respectively, and 5 random areas were selected for observation under an inverted microscope. Quantification of differentiation specific genes was also performed as described later on.
Oil Red O staining
Oil Red O staining was used to evaluate lipid vacuole accumulation during adipogenic differentiation. Briefly, cells were fixed for 1 h with 10% natural formalin buffer, washed with 60% isopropanol, allowed to dry completely, and stained for 10 min with 0.18% Oil Red O solution in isopropanol. Lipid vacuoles were stained red.
Alizarin Red S staining
Alizarin Red S staining was used to evaluate Ca2+ deposition during osteogenic differentiation. Briefly, cells were washed twice with PBS, fixed with 10% natural formalin buffer, washed with ddH2O, and stained for 15 min with 2% Alizarin Red S (pH 4.1). Ca+2-rich extracellular matrix was stained red. All chemicals used in both staining were purchased from Sigma-Aldrich Co.
Immunosuppression
mMSCs were irradiated (30 Gy) and plated out in serial dilutions onto a flat-bottom 96-well plate (Corning). Splenocytes from BALB/c mice were isolated by mechanical dissociation through a 70-μm nylon cell strainer (BD Biosciences). Red blood cells (RBCs) were lysed by incubation for 5 min on ice with NH4Cl (0.15 M). Upon adherence of the irradiated mMSCs, 105 splenocytes and anti-mouse CD3ɛ antibody (BD Biosciences; 10 μg/mL) were added to each well. After 48 h of incubation, cells were pulsed with [methyl-3H]thymidine (Amersham; specific activity: 53.0 Ci/mmol, 1 μCi/well) for an additional 18 h, harvested, and counted on a liquid scintillation counter (Wallace Trilux, no. 1450 Microbeta; PerkinElmer Life Sciences).
RNA isolation and quantitative reverse transcriptase–polymerase chain reaction analysis
Total RNA was extracted from mouse and hMSCs using either the Nucleospin RNA II kit (Macherey-Nagel) or TriZOL reagent (Invitrogen), according to the manufacturer's instructions. rDNase treatment was performed to remove any residual genomic DNA. Total RNA (1.5 μg) was reverse transcribed using ImProm-II Reverse Transcriptase (Promega) and random hexamer oligonucleotides (Invitrogen). Quantitative reverse transcriptase–polymerase chain reaction (qRT-PCR) was performed on a Rotor-Gene 3000 (Corbett Research). PCR conditions were 95°C for 10 min, with 35 cycles of 95°C for 20 s, 58–63°C for 20 s, depending on primer pair, and 72°C for 30 s. cDNA solution (150 ng), Platinum SYBR Green qPCR Supermix-UDG reaction buffer (Invitrogen), and 100 nM of primers were used in a 25 μL reaction mixture of PCR. Primer pairs used are shown in Table 1.
All mMSC gene expression levels tested in this study were normalized to Theiler Stage 13 (E8.5). Theiler Stage 13 is one of the most primitive stages of the developing embryo that we could isolate. All genes tested are expressed at this stage as reported by Davis et al. [33] and in the Mouse Development handbook [34]. In particular, regarding Dazl, similar levels of expression were found in both mice spermatozoa and E8.5 tissues (data not shown). Human CRD-BP/IMP-1 levels were normalized to K562, a leukemic cell line overexpressing CRD-BP/IMP-1, and human DAZL was normalized to human sperm tissue; human and mouse GAPDH mRNA levels were used, respectively, for internal normalization. The ΔΔC t method was used for data analysis of gene expression levels.
Regarding lipoprotein lipase (Lpl) and osteocalcin, also known as bone gamma-carboxyglutamic acid-containing protein (Bglap), adipogenic and osteogenic differentiation specific genes, respectively, samples were tested at the time of initiation (D0) of the differentiation assay and on the termination day (D14). Gene expression levels of D14 were normalized versus D0. Expression at D0 was arbitrarily defined as 100. Adipose and bone tissue were utilized as positive controls in this assay.
Statistics
The Student's t-test with a 95% confidence interval was performed using GraphPad Prism version 5.00 for Windows, GraphPad Software (
Results
Proliferative and self-renewal potential
In the present study, the BALB/c strain was used for the isolation and expansion of mMSCs. BALB/c is a well-known inbred mouse strain and among the most widely used in animal experimentation [35]. Initially, we determined the clonal capacity, growth potential, and proliferation rate of mMSCs. All parameters tested in this report were analyzed according to donor age, sex, and passage. Hence, BM samples were divided, as reported recently [36], into 3 categories according to their developmental stage in the postnatal organism: in infants (<4-week-old mice), adolescent (5–12-week-old mice), and adult (13–34-week-old mice). Moreover, cultured cells were categorized, arbitrarily, into 4 different groups according to the number of passages: (a) 3–7, (b) 8–12, (c) 13–17, and (d) 18–24. It should be noted here that mMSC properties were not studied in later passages because of a recent publication that deemed MSCs to be tumorigenic, upon spontaneous malignant transformation with long in vitro culture [37]. Further, we tested whether the gender of the mice affects the properties of MSCs as more studies either use female mice or do not take sex into account [2,28].
Initially, the MSC frequency in the BM was determined by the colony forming units-fibroblast assay. A decrease in the frequency of progenitors was observed as we moved from the infant to the adult group. The number of colonies obtained from 106 BMMNCs plated and expanded for 14 days was 61.83 ± 21.77 (1 per 16,173 nucleated cells) in infant donors and 17.7 ± 8.8 (1 per 56,947 nucleated cells) in adult donors. Therefore, the calculated frequency of MSCs in the BM of the adult organism was found to be reduced by approximately 3.5 times compared with that in early postnatal development. As demonstrated in Fig. 1A, this decrease was extremely significant in all groups. More importantly, MSC frequency was sex dependent, with female BM consisting of an extremely significant lower number of progenitor cells in all age groups (Fig. 1B).
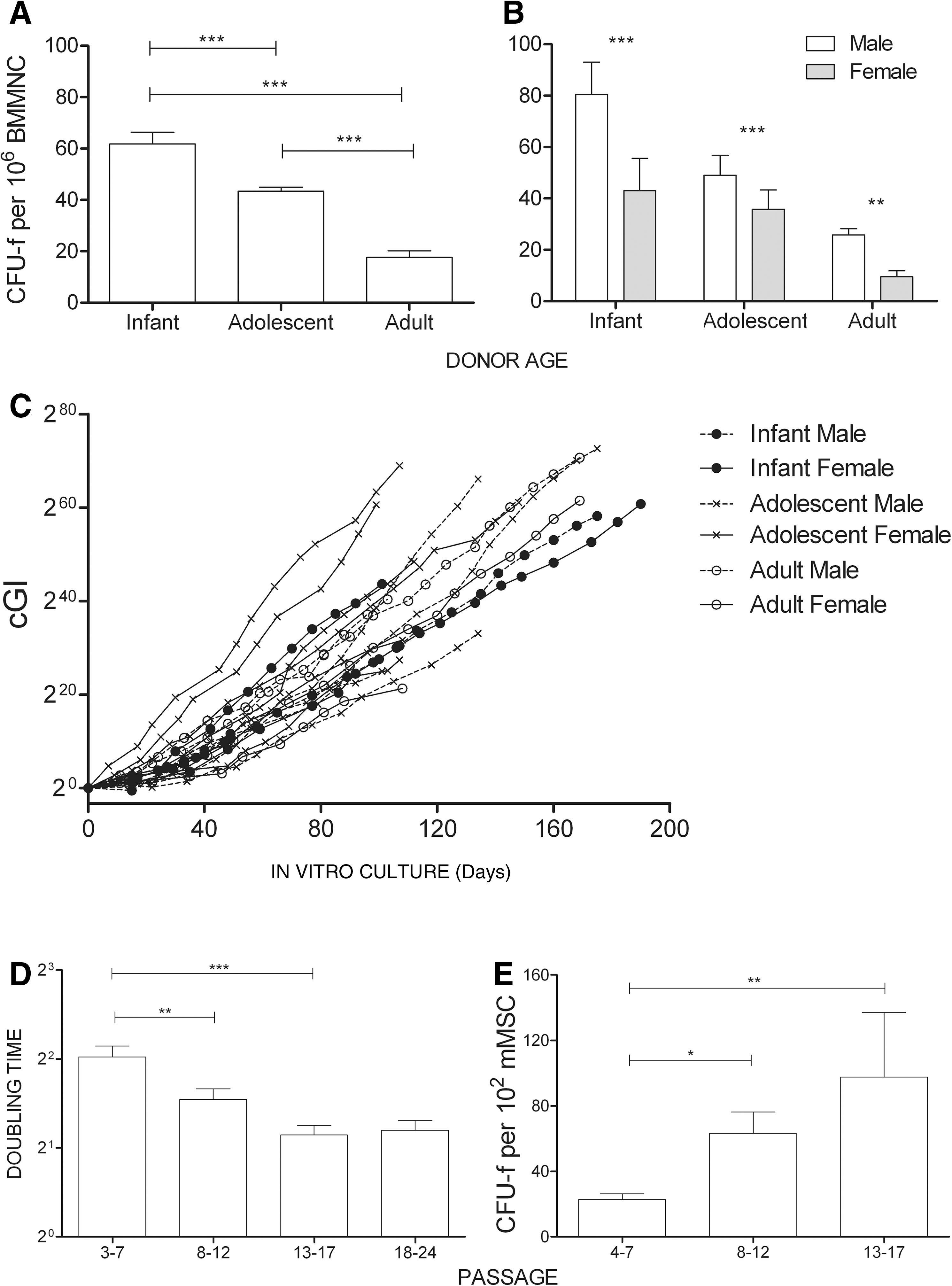
Growth potential of BM-derived mMSCs. Differential mMSC frequency in the BM according to age
Upon selection and in vitro expansion, all preparations, independent of the age and sex of the donor, showed a similar proliferation capacity as demonstrated by the calculated cumulative GI (Fig. 1C). Further, mMSCs seemed to be able to proliferate “indefinitely”—for more than 70 PDs—showing a tendency for increased proliferation rate with passage as determined by population T D (Fig. 1D). The increase in proliferation rates was further confirmed with the colony forming units-fibroblast assay. In vitro propagation resulted in an increased number of colonies (Fig. 1E).
Phenotypic characterization
In vitro expansion of mMSCs caused a notable reduction of cell size values and complexity (granularity) as depicted by the cell distribution of the same BM preparation at passages 3 and 19 according to forward and side scatter values (Fig. 2A, B). mMSCs of different preparations were analyzed by FACS and their forward and side scatter values were plotted against passage. Statistically significant differences were observed in both parameters (Fig. 2C, D). It is important to note that statistical significance could not be reached at passages 18–24 because of the small sample number analyzed (n = 4). On the other hand, the size and complexity of the cells did not differ significantly with donor age (data not shown).
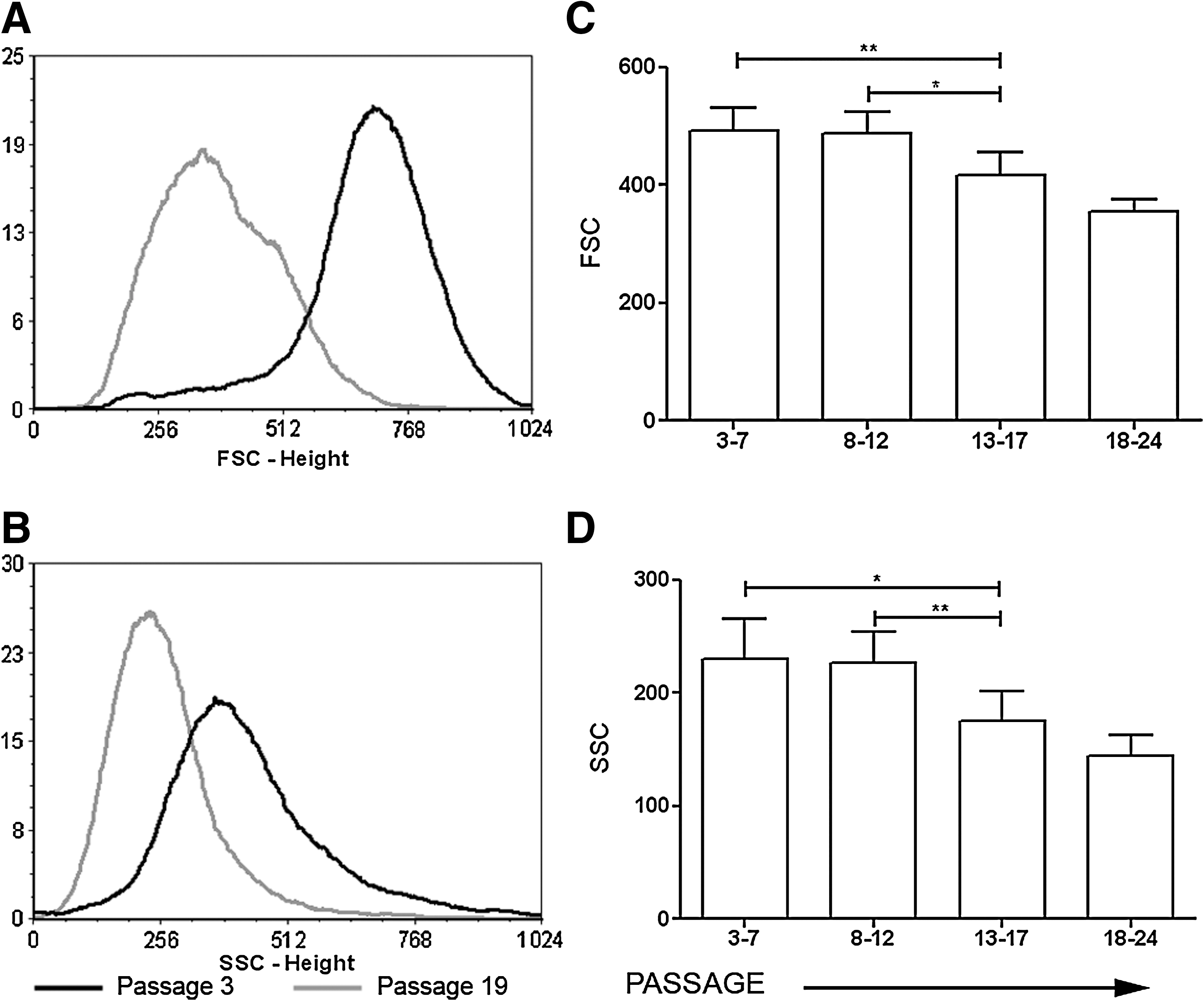
Effect of extensive subcultivation of MSCs on cell size and granularity. Representative histograms of the shift between passages 3 and 19 in FSC
As a single antigen to directly define mMSCs does not exist, a panel of surface antigens is commonly used to characterize this population [38]. We adapted our analysis from previous studies by Anjos-Afonso et al. [15], who examined different types of cell surface antigens such as hematopoietic antigens, adhesion molecules, integrins, etc., and determined a more precise immunophenotype of mMSCs. In our experimental setup, selection of the mMSC population (CD45− and CD11b−) was achieved by short trypsinization from passages 0 to 2 as recommended by Peister et al. [28]. We started our analysis of the mMSC phenotype at passage 3 onwards. A representative experiment of FACS analysis is depicted in Fig. 3A. MSCs were stained negative for CD45, CD11b (not shown), and MHC class II molecules. CD105 expression was at low/negative levels, whereas Sca-1, CD29, CD44, and MHC class I molecule expression was scored positive. CD73 expression ranged from 0% to 88%. In contrast to CD73 and Sca-1, CD29 and CD44 expression was independent of donor age and in vitro culture (Fig. 3B, C). A statistically significantly higher percentage of the mMSC population stained positive for CD73 during adolescence and adulthood in comparison to mMSCs isolated from infant mice; no changes in the fluorescence intensity of the positive fraction of mMSCs were observed. In contrast, Sca-1 levels demonstrated a decline in extreme passages. Its high expression (91.77% ± 3.6%) detected up to passage 12 was significantly reduced (52.93% ± 13.45%) at late (18–24) passages. The decrease in the percentage of expression was also accompanied by a decrease in its density on the cell surface as this was detected by a decrease in the fluorescence intensity of the positively staining population.
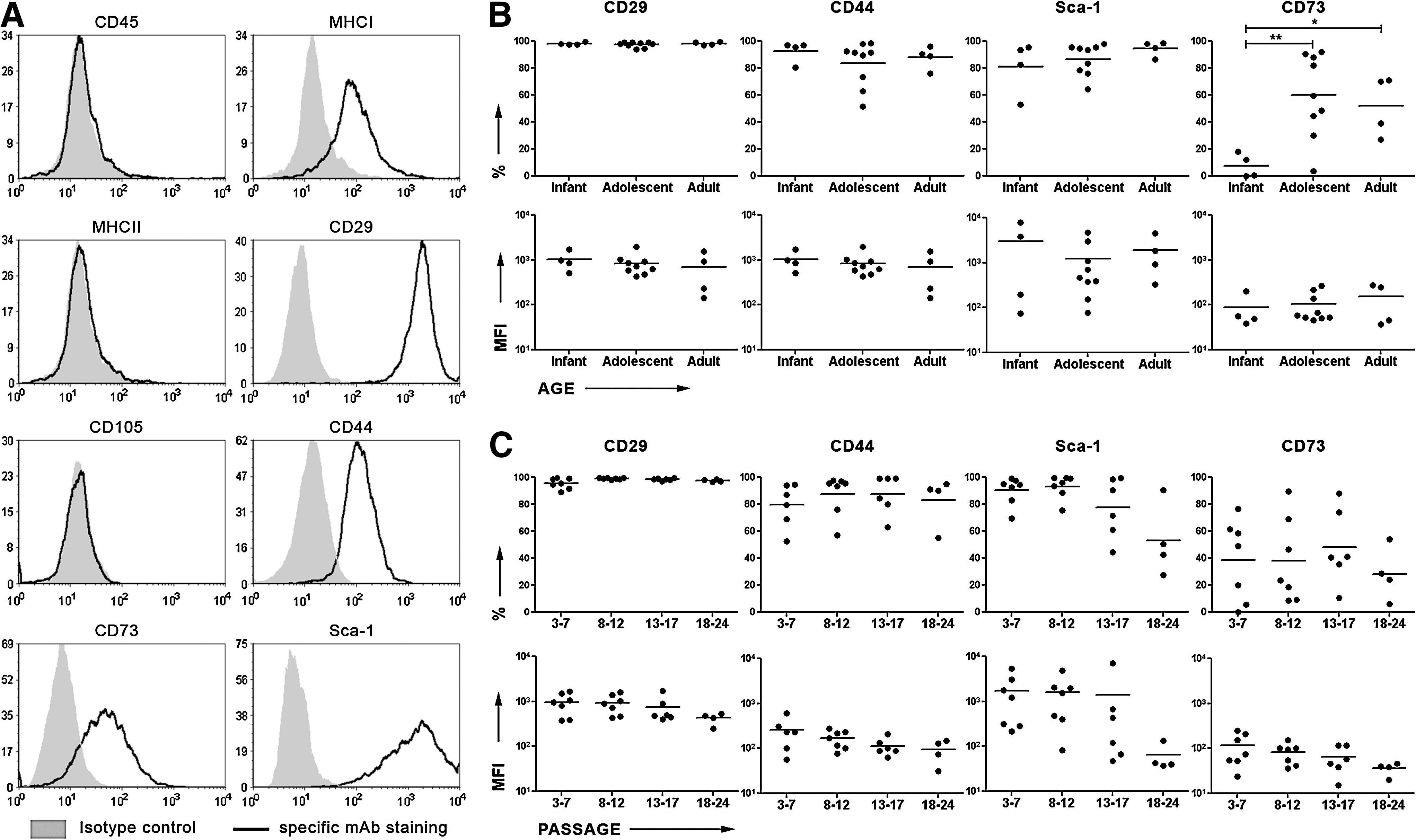
Immunophenotypic characteristics of mMSCs.
Functional characterization
There is currently little consensus, and in many cases, conflicting reports regarding the effect of donor age and passage progression on MSC function exist. A number of studies have previously shown no age-related differences in in vitro differentiation using hMSCs [39]. Similar results were obtained in our experimental setup, by optical evaluation of the number of differentiated cells. mMSCs independent of gender, age, or passage differentiate into both adipocytes and osteocytes (Fig. 4A), although differences in the percentage of differentiating cells were observed among all groups, ranging from 40% to 100%. As this technique would only reveal gross variations, we decided to examine the expression levels of 2 established differentiation markers, Lpl and Bglap, by qRT-PCR. Lpl is an enzyme that hydrolyzes lipids in lipoproteins [40]. On the other hand, Bglap is a noncollagenous protein found in bone and dentin. It is secreted solely by osteoblasts and is implicated in bone mineralization and calcium ion homeostasis [41]. Thus, mMSCs isolated from male subjects express higher levels of both markers (Fig. 4Bi, 4Ci). In addition, the levels of expression of Lpl seem to be independent of donor age in contrast to Bglap levels, which show a tendency to decrease with age progression (Fig. 4Bii, 4Cii). Moreover, Lpl expression tends to decrease with in vitro culture progression in contrast to Bglap levels, which significantly increase upon in vitro expansion (Fig. 4Biii, 4Ciii).
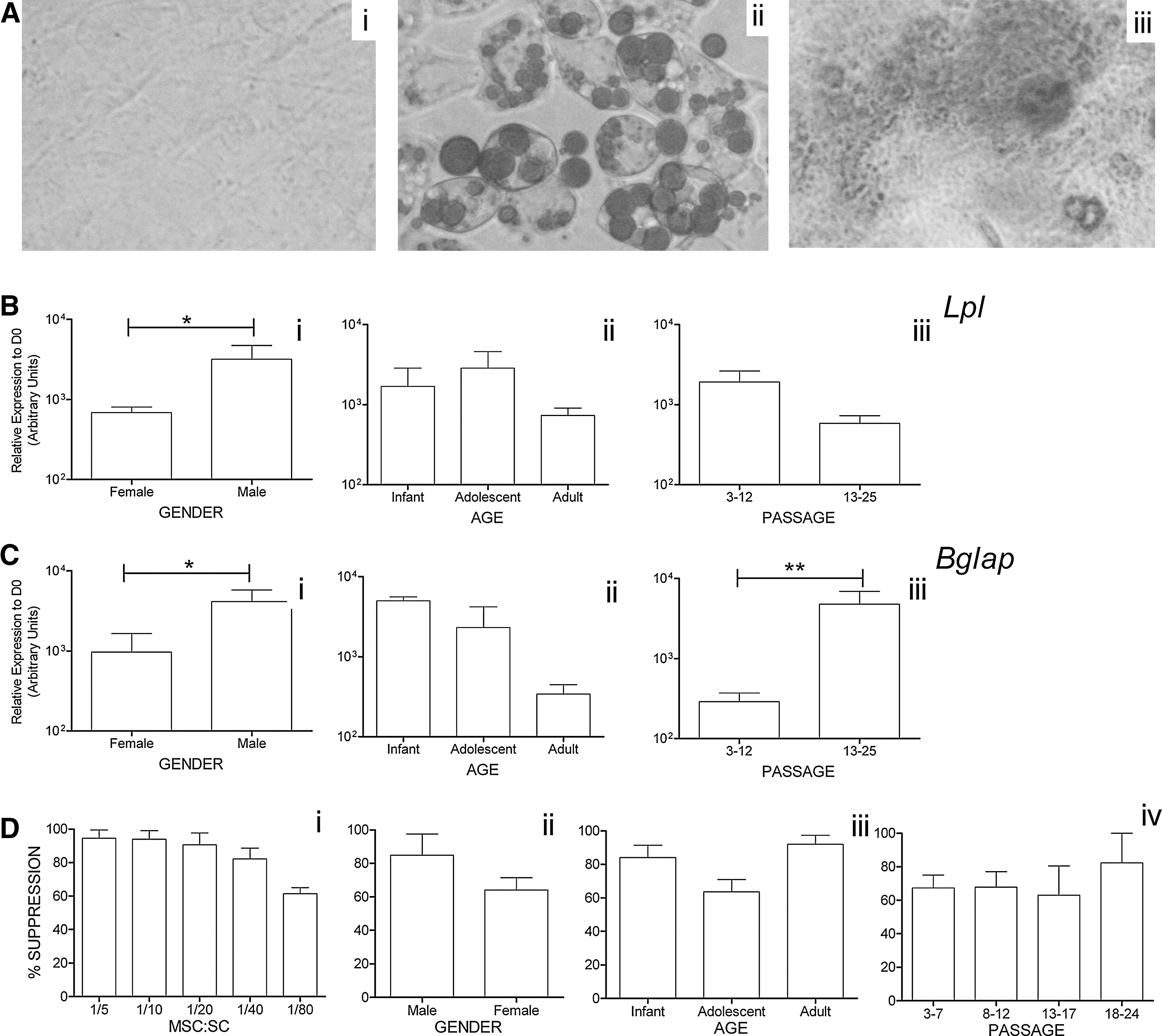
Functional properties of in vitro cultured mMSCs.
MSC-mediated inhibition of T-cell proliferation was utilized throughout our experiments as a standard assay characterizing a functionally intact MSC population. T cells were activated by CD3e crosslinking at coculture, set up at different MSC-to-spleen cell (SC) ratios, and it was found that mMSCs were immunosuppressive even at a very low cellular MSC-to-SC ratio (1:80) (Fig. 4D). Because all samples exhibited ≥95% suppression at high MSC-to-SC ratios (1:5 and 1:10), we chose to use suboptimal conditions (1:20 MSC-to-SC ratio) to unravel any possible differences among the various tested parameters. Percentage of suppression of mMSCs at 1:20 MSC-to-SC ratios ranged from 60% to 100%. Slight variations were detected, but with no statistical significance between male and female donors (Fig. 4E), among the different age groups examined (Fig. 4F), as well as in relation to passage progression (Fig. 4G).
Molecular characterization
hMSCs have been found to express several embryonic markers [16]. They share a multitude of characteristics with their mouse counterparts but also retain important species-specific differences [9,42]. Further, model organisms such as mice are frequently used to decipher various biological phenomena. In this context, we proceeded to determine whether mMSCs have a similar molecular profile to hMSCs. Three important ESC markers (Oct3/4, Sox2, and Nanog) were examined, as well as 2 other developmentally important genes (DazL and Crd-bp/Imp-1), not previously investigated in these populations.
Initially, various randomly selected samples from different passages of mMSCs were tested for the expression of Oct3/4, Sox2, and Nanog (Fig. 5A). All samples were found positive for Nanog, whereas Oct3/4 and Sox2 mRNAs were infrequent. Additionally, the levels of expression of these 3 genes were determined by qRT-PCR, in relation to donor age, passage progression, and lineage commitment.

Gene expression profile.
Nanog was expressed redundantly in all samples tested, with levels of expression almost uniformly higher to those in the developing embryo E8.5 as well as to those in the total BM cell population. It should be noted that Nanog levels seemed to gradually decline in relation to donor age, reaching statistical significance between the adolescent and adult population. Statistical significant differences between the infant and adult groups could not be detected, possibly because of the low sample number in the infant group (n = 4) (Fig. 5B). However, “in vitro cellular aging” (passage progression) did not affect Nanog expression levels. Additionally, a statistically significant decrease was observed during adipogenesis but not during osteogenesis (Fig. 5D). We could barely detect any Oct3/4 and Sox2 mRNA expression in the majority of the tested samples. Donor age, passage progression, and lineage commitment did not affect their levels of expression (data not shown).
In the course of our studies, we also investigated the expression of 2 developmentally important genes in mMSCs: DazL and Crd-bp/Imp-1 (Fig. 5A). As it was the first time that these genes were investigated in the MSC population, we also examined their expression in hMSCs. Dazl was detected in mMSCs, although in relatively low levels and always lower than that in the total BM cells tested. As shown in Fig. 5B–D, no significant alterations in gene expression were observed with donor age, with passage progression, or during MSC differentiation. In contrast, overall Crd-bp/Imp-1 levels of expression in mMSCs were rather high and almost always similar to the embryo E8.5 levels (Fig. 5B–D). It is worth noting that Crd-bp/Imp-1 levels tend to decrease in adult-derived mMSCs, whereas total BM cells were found to be negative. In addition, the levels of Crd-bp/Imp-1 remained high throughout in vitro expansion. There was a statistically significant increase in the expression levels between early passages (3–7) and passages 18–24 (Fig. 5C). Moreover, Crd-bp/Imp-1 levels increased significantly when mMSCs were triggered to differentiate toward the osteogenic lineage but remained unaffected during adipogenesis (Fig. 5D).
The high expression levels of Crd-bp/Imp-1 prompted us to investigate 3 of its major targets—c-Myc, insulin-like growth factor 2 transcript 3 (IgfII-leader3′), and h19. c-Myc exhibited a constant, high expression level among all ages and in vitro passage. The c-Myc levels were similar or higher than these detected in the E8.5 and total BM. In addition, c-Myc followed a pattern similar to Crd-bp/Imp-1 upon osteogenic differentiation, with statistically significantly increased levels of expression (Fig. 5D). In contrast, IgfII-leader3′ and h19 levels were very low or even undetectable, similar to those in total BM cells. No alterations in the expression of these genes were detected, with the exception of IgfII-leader3′ levels, which were further significantly reduced during adipogenic commitment (Fig. 5D). As far as hMSCs are concerned, DAZL could not be detected in any sample isolated from the BM, in contrast to CRD-BP/IMP-1, which was expressed in relatively high levels, similar to those of its mouse counterpart (Fig. 5A).
Discussion
The characterization of the MSC population has been the focus of numerous studies in the past decades. In this report, we thoroughly investigated the properties of mMSCs in relation to donor age and in vitro expansion as well as the expression pattern of well-established embryonic markers.
In our experimental setup, mMSCs were isolated from both genders of BALB/c mice during different developmental stages of postnatal life. Our finding that female BM, from infancy to adulthood, contained significantly lower MSC numbers—in comparison to the corresponding male—is in accordance with that by Strube et al., who conducted their experiments on Sprague–Dawley rats and observed a sex-specific compromised bone healing in female rats due to a decrease in MSC quantity [43]. These observations could possibly propose a reason why women are more prone to osteoporosis, as one of the mechanisms involved in its development is the inadequate formation of new bone during remodeling.
Further, the intrinsic growth potential of explanted MSCs diminished with age progression. However, MSCs showed similar proliferation capacities in vitro, independent of age or sex, and—upon overcoming the resting state and, subsequently, the low turnover exhibited within the BM—retained their proliferative capacity for up to 5 months in culture, that is, for more than 70 PDs. Similar observations have been previously reported in various studies [32,44,45]. In these studies, extended in vitro expansion of mBM-MSCs for up to 1 year has been documented. This could not be attributed to strain specificity or methodology variations, because both BALB/c- and C57BL/6-derived BM-MSCs behaved similarly as the isolation–expansion methodologies used do not significantly differ from the ones presented herein. This proliferative capacity exhibited in vitro does not necessarily represent the in vivo potential of MSCs but confirms that the cells possess the ability to proliferate indefinitely if the necessary signals are provided. Therefore, these findings could possibly support the extrinsic theory of aging—which states that cells are driven into proliferative silence by changes within the surrounding tissue [9].
In addition, we observed that mMSCs decrease their T D with in vitro culture and diminish in cell size as well as in granularity. In contrast, hMSCs cease to proliferate within 20–50 PDs, and cells enlarge and become more granular. Ultimately, they stop cell division [46]. This significant difference between human and mouse BM-MSC could be attributed to either technical reasons (ie, in vitro conditions may affect them differently) or diversity between species, including lifespan, tissue renewal requirements due to environmental differences, etc.
Regarding the mMSC phenotype, neither donor age nor in vitro culture affect CD29 and CD44 expression, whereas CD73 expression is significantly increased in correlation to donor age. We report that mMSCs isolated from adolescent and adult mice express at least 5 times more CD73 than infant-isolated MSCs. The physiological role of CD73/ecto-5′-nucleotidase differs in various organisms and tissues and extends beyond its enzymatic activity. Resta et al. have stated that mature versus immature mouse T and B lymphocytes show a 10-fold greater activity of CD73, which is necessary for their adhesion and migratory capacity [47]. Nevertheless, its actual role in MSC populations needs to be further clarified. Further, Sca-1 cell surface expression declined with passage progression, although statistical significance was not reached. As mentioned earlier, passage progression correlated with decreased cell size and granularity. Our results are in accordance with Meirelles Lda and Nardi, who observed low Sca-1 expression in an MSC subpopulation with low cell size and complexity [32]. These observations support the notion that BM-MSCs are heterogeneous, including subpopulations of different proliferative capacities. It is conceivable that one of these subpopulations becomes predominant under long-term ex-vivo expansion conditions. It would be interesting to further study this possibility.
mMSCs exert a very strong suppressive activity in vitro even in very low numbers in contrast to hMSCs, which exert their in vitro immunosuppression at relatively high numbers. mMSCs' suppressive ability was tested throughout in vitro culture and in relation to sex and age and was not significantly affected by either of these parameters. Immunomodulation by MSCs is mostly exerted via soluble factors, including transforming growth factor-β, hepatocyte growth factor, and nitric oxide [48], which differentially affect each immune system compartment. Our study addressed only a part of the suppressive ability of MSCs—their effect on T cells—which remains unaltered by donor age and in vitro propagation. Although individual factors produced by MSCs may be affected during postnatal life and/or in vitro expansion, it is possible that the end effect (immunosuppression) remains unaltered, as MSC-mediated immune regulation is the result of the cumulative action displayed by several molecules [48].
As far as the differentiation ability of MSCs is concerned, multitudes of controversy exist about the effect of age and passage on their potential. Kretlow et al. [14] reported that the osteogenic and adipogenic differentiation potential of murine BM-MSCs decreased with donor age. Gou et al. have recently reported that long-term–cultured mBM-MSCs could differentiate into adipocytes only but not into osteocytes or chondrocytes [44]. In a review article, Sethe et al. [9] reported that the osteogenic potential of MSCs decreased with age in almost all the studies included, irrespective of species origin, which is in line with the data presented herein. Regarding the adipogenic properties of the cells, their potential was found either to remain unaltered or to increase with age [9]. The high heterogeneity of our data with respect to the adipogenic potential of mMSCs with age could not lead us to conclusive results, although a tendency toward reduced differentiation potential was also observed in older donors. This could possibly reflect the diminished regenerative potential with age [49]. In agreement with previously reported data [50,51], we show that the adipogenic potential of mMSCs tends to decrease with passaging in contrast to their osteogenic potential, which is significantly increased in high passages. Notably, in our study, male MSCs exhibit stronger differentiation potential toward both the osteogenic and adipogenic lineages, which, to our knowledge, has never been previously reported. This observation should be further investigated under the prism of gender physiological differences.
Developmental alterations are guided by the orchestrated, differential expression of genes. ASCs should in some way retain a part of the ESC characteristics as descendants of those initial stem cells that create the fetus and in continuity the postnatal organism [52]. Oct3/4, Sox2, and Nanog are considered the key regulators of pluripotency in ESCs of both human and mouse origins [16,53 –57]. As shown in the present study, Nanog is abundantly expressed in mMSCs, in accordance with previous studies on hMSCs of different origin [16] and unpublished observations (Mahaira et al., unpublished data). Nanog is a homeodomain protein whose intrinsic regulators of expression expand beyond its control by Oct3/4 and Sox2. Specifically, Nanog is controlled by Forkhead box D3 and is a direct downstream effector of the leukemia inhibitory factor-signal transducer and activator of transcription-3 pathway in maintaining ESC pluripotency. In addition, stringent control of Nanog expression is necessary for guided differentiation toward a specific lineage. As demonstrated by Suzuki et al. [58], t brachyury (T) and signal transducer and activator of transcription-3 coordinately bind to a regulatory element in the mouse Nanog promoter, resulting in increased Nanog expression in early mesoderm-specified progenitors. MSCs could possibly retain the molecular signature of early mesoderm-specified progenitors, part of which is high Nanog expression levels. This hypothesis as well as Nanog downregulation during adipogenesis opens an interesting field of investigation. The role of the major ESC factors Oct3/4 and Sox2 remains an issue of debate. BM-derived hMSCs were found to be positive for OCT3/4, but SOX-2 was not detected [16], whereas the expression of both was reported in a different study [59]. Little is known about the functional role of these pluripotency markers in ASCs [16]. All these controversial results may suggest that the major ESC factors exhibit a highly differential expression pattern, probably reflecting the significant divergence existing between embryonic and adult stem populations or stem populations of different origins.
Two novel genes (Dazl and Crd-bp/Imp-1) were explored in the MSC population. Dazl expression was recently reported in stem cells originated from amniotic fluid [60] and in ESCs [23]. We could not detect DAZL in hMSCs. On the contrary, mMSCs were positive for Dazl, although in low but constant levels of expression. It is important to report that we also detected Dazl in total murine BM cells in levels similar to embryonic and higher than those in mMSCs. Hua et al. reported that hMSCs from fetal lung can give rise to germ cells in vitro [61]. Additionally, Johnson et al. reported that a putative germ stem cell reservoir that supports oogenesis in adult female mice appears to reside in BM [62]. Nayernia et al. achieved the derivation of male germ cells from BM stem cells [63]. Thus, it remains to be clarified whether mMSCs are a potential germ cell source and what the implication of Dazl expression is.
Crd-bp/Imp-1 expression is documented here for the first time in both mouse and human MSCs. mMSCs isolated from the BM were found to express high levels of Crd-bp/Imp-1, independent of donor age or in vitro passage. This mRNA-binding protein has at least 4 identified targets: c-Myc, IGFII, H19, and tau mRNA (which is a neuron-specific gene). It has been postulated that CRD-BP/IMP1 affects the posttranscriptional fate of its targets by determining their subcellular localization, by modulating their half-life, or by affecting their translatability [27]. To elucidate the possible role of Crd-bp/Imp-1 in MSC physiology, we examined the pattern of expression of its targets. Both IgfII-leader3′ and h19 mRNA levels were relatively low to undetectable. Nevertheless, a statistically significant reduction in IgfII-leader3′ expression was detected during adipogenic commitment, which is in line with the finding by Jones et al. [64], who detected a reduction in IgfII-leader3′ expression accompanied by increased fat deposition in adult mice fat pad.
A major target of Crd-bp/Imp1 is c-Myc, a multifunctional, nuclear phosphoprotein involved in cell cycle progression, apoptosis, and activation of many ESC-specific genes with a role in metabolic regulation during the early steps of dedifferentiation [65]. In our study, c-Myc expression was detected in all samples tested, and its levels were significantly higher than in embryos. This high expression was influenced by neither donor age nor passage. Our data along with the well-established role of c-Myc in cell cycle progression may support the notion that Crd-bp/Imp1 constant high expression leads to constant c-Myc levels required for cell population maintenance [66,67].
Additionally, both Crd-bp/Imp1 and c-Myc expression followed the same pattern upon differentiation to the osteogenic lineage and a statistically significant induction of their expression was detected. This could be attributed to the fact that increased apoptosis is necessary to enhance the progress of bone formation, thus justifying the elevated expression of c-Myc during osteogenesis, as c-Myc is a major apoptosis inducer [68,69]. All the above suggest that Crd-bp/Imp1 gene possesses a coordinative role in stem cell physiology.
In summary, the data presented herein show that mMSCs exhibit high similarity to their human counterparts in terms of in vitro and in vivo aging: their frequency in vivo declines significantly during postnatal life, but this population per se retains most of its functional and phenotypic properties when expanded in vitro. However, essential differences between mMSCs and hMSCs, such as their in vitro proliferative capacity, need to be taken into serious consideration when translating data from mouse models into humans. The great similarities, including Nanog and CRD-BP/IMP1, and discrepancies (eg, Oct3/4 and Sox2) observed in the expression profile of important stemness regulatory genes could possibly unravel mechanisms underlying different cellular physiology and account for species variation.
Footnotes
Author Disclosure Statement
No competing financial interests exist.