
Abstract
Photoreceptors have recently been generated from mouse and human embryonic stem cells (ESCs), although ethics concerns impede their utilization for cell replacement therapy for retinal disease. Extra-embryonic tissues have received attention as alternative therapeutic sources of stem cells. Human and mouse amniotic fluid-derived stem cells (AFCs) have been reported to be multipotent and express embryonic and adult stem cell markers. Here, in vitro conditions that generate retinal cells from ESCs were used to analyze and compare the retinal potential of murine AFCs and ESCs. We show that AFCs express pluripotency markers (Nanog, Sox2, and Oct3/4) as well as retinal transcription factor genes (Et, Lhx2, Tll1, Six6, Otx2, Pax6, and Fgf15). AFCs from amniotic fluid of Fgf15.gfp, Nrl.gfp, and Crx.gfp embryos cultured in retinal proliferation and differentiation conditions failed to switch on these retinal transgenes. AFCs cultured in retinal-promoting conditions, effective on ESCs, showed reduced expression of retinal markers. Retinal co-cultures activated retinal genes in ESCs but not in AFCs, and migration assays in retinal explants showed limited migration of AFCs compared with ESCs. Unlike ESCs, AFCs do not express the early embryonic ectodermal gene Utf1 and Western analysis of AFCs identified only the B isoform of Oct3/4, rather than the isoform A present in ESCs. We conclude that AFCs have restricted potential and differ considerably from ESCs and retinal progenitor cells. Reprogramming to induce pluripotency or new differentiation protocols will be required to confer retinal potential to AFCs as expression of a subset of pluripotency and retinal markers is not sufficient.
Introduction
I
Extra-embryonic tissues, such as placenta [17 –19] and umbilical cord [20,21], have received significant attention as alternative sources of stem cells for therapeutic use as they avoid the ethical controversy associated with the use of human embryos. The amniotic fluid fills the amniotic cavity and provides a supportive environment for the developing fetus throughout pregnancy. Amniocentesis involves the collection of a sample of amniotic fluid and is a common screening tool used for the human fetus that also could provide a source of cells for regenerative medicine. Recently, a population of human and murine amniotic fluid-derived stem cells (AFCs) have been reported that express markers characteristic of both ESCs and adult stem cells [22]. AFCs, unlike ESCs, are particularly attractive for allogeneic transplantation, due to their immunoprivileged nature [21 –23]. Moreover, AFCs (unlike ESCs) are not tumorigenic and are capable of differentiating into cell types representing each embryonic germ layer, conferring them characteristics valuable for therapy [22,24]. De Coppi et al. [22] used retroviral marking to demonstrate that clonal AFCs can give rise to adipogenic, osteogenic, myogenic, endothelial, neurogenic, and hepatic lineages, inclusive of all embryonic germ layers [22]. In this respect, they meet a commonly accepted criterion for pluripotent stem cells. Several studies have demonstrated the capability of human AFCs to give rise to multiple differentiated cell types, including neuronal lineage differentiation [22,24 –32]. However, despite the value of mouse genetic models for elucidating cell potential and the possibility of generating retinal cells from this extra-embryonic cell source, little work has investigated mouse AFCs.
AFCs come either from the amnion or from the fetus [33,34] that is immersed in the amniotic fluid throughout pregnancy, and since several different precursor cell lineages have recently been isolated from the amniotic fluid [25,26,29,30,32,35], we evaluated whether committed retinal cells are present. Moreover, taking advantage of 3 different mouse transgenic reporter lines, each expressing green fluorescent protein (GFP) under the control of a retinal progenitor/photoreceptor gene promoter, we assessed whether or not AFCs could commit to a retinal progenitor and photoreceptor cell fate in defined culture conditions, and compared them with ESCs, which have previously been shown to have retinal potential.
Materials and Methods
Animals
Mice were maintained in the animal facility at University College London. All experiments were conducted in accordance with the Policies of the Use of Animals and Humans in Neuroscience Research, revised, and approved by the Society of Neuroscience in January 1995. Animal strains used included Nrl.gfp from A. Swaroop [36], Cba.gfp [37] and Crx.gfp from C. Cepko [38], and Fgf15.gfp kindly provided by N. Allen (Cardiff, United Kingdom).
AFC isolation
Amniotic fluid samples and c-kit+ AFCs were harvested from embryos at embryonic day (E) 11.5–12.5 as described in Supplementary Materials and Methods (Supplementary Data are available online at
ESC and AFC propagation
Mouse ESCs expressing GFP (ES cell line E14Tg2A [38 –40]) were expanded directly on 1% gelatine in NIWA medium as previously described [40]. AFCs were cultured on a modified mouse embryonic fibroblast (MEF) feeder layer producing leukemia inhibitory factor (LIF) in polystyrene (Petri) dishes coated with 0.1% gelatine in NIWA medium at 37°C with 5% CO2. The feeder layer was inactivated with Mitomycin-C (10 μg/mL; Sigma) at 37°C for 2.5 h. About 75% of the medium was changed every other day. AFCs were passaged every 15–20 days depending on the appearance of the feeder layer, at a dilution of 1:2 in trypsin-EDTA (0.25%; Sigma) for 10 min at 37°C. ESCs were passaged every week at a dilution of 1:3 in trypsin-EDTA.
Fluorescence-activated cell sorting
As described in Supplementary Materials and Methods.
Retinal proliferation medium
Embryonic retinal progenitor cells (RPCs) were isolated from E11.5-12.5 C57Bl/6, Fgf15.gfp, Crx.gfp, and Nrl.gfp litters and cultured in retinal proliferation medium as described in Supplementary Materials and Methods. AFCs were cultured in the same retinal proliferation medium with the following substrates: poly-L-lysine (100 μg/mL) or poly-L-lysine/laminin (5 μg/mL), in floating conditions (without substrate) or on the LIF producing MEF feeder layer.
Retinal differentiation medium
RPCs from Crx.gfp and Nrl.gfp lines were cultured for 2–3 days in retinal proliferation medium and then switched to retinal differentiation medium; cultures were treated for 4–6 days with N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butylester (DAPT) to activate GFP expression as described in Supplementary Materials and Methods.
Retinal lineage commitment in ESC and AFC cultures: 2-step differentiation protocol
ESCs were grown according to the Takahashi laboratory protocol [13,14] in serum-free floating cultures of embryoid body-like aggregates (serum-free embryoid bodies (SFEB)] with 5% of knockout serum replacement for 5 days. Neural retinal progenitors were induced using Dkk1, Lefty-A (DLFA), fetal calf serum, and Activin A treatment (SFEB/DLFA; 1° differentiation step) as described in Supplementary Materials and Methods.
ESC-derived retinal progenitors were subjected to a second step of differentiation (2° differentiation step) to induce photoreceptors as described in the Takahashi laboratory protocol [13,14] and in Supplementary Materials and Methods.
GFP-positive AFCs were differentiated using the same protocols as for ESCs in SFEB/DLFA conditions, or DLFA was added sequentially to AFCs on feeder layers as described in Supplementary Materials and Methods.
AFC and RPC co-cultures
Retinae from C57Bl/6 mouse embryos at E11.5-12.5 or P4 were co-cultured with AFCs derived from Nrl.gfp/Fgf15.gfp pregnancies as described [41] and detailed in Supplementary Materials and Methods. Reaggregated pellets of P4 retinae with Nrl.gfp AFCs were cultured with (+) or without (−) DAPT. Pellets of E11.5-12.5 retinae with Fgf15.gfp AFCs were cultured with (+) or without (−) DLFA. Alternatively, DLFA-treated ESCs were co-cultured with retinal cells at P4.
Immunohistochemistry and immunocytochemistry
As described in Supplementary Materials and Methods.
RNA isolation and reverse transcription
As described in Supplementary Materials and Methods.
Polymerase chain reaction
As described in Supplementary Materials and Methods. All polymerase chain reaction (PCR) experiments were performed at least 3 times on at least 3 independent samples. Representative images are shown of reproducible findings.
AFC, ESC, and RPC transplantation on adult retinal explants
Retina explant cultures were prepared as previously described [41] with minor changes as described in Supplementary Materials and Methods. AFCs from E11.5-12.5 Cba.gfp mice, GFP-labeled ESCs, or in vitro differentiated Nrl.gfp-positive photoreceptor precursors were placed onto adult retinal explants and cells migrating into the explant were analyzed after 7 days in culture by sectioning; all 12 μm sections (n = 40–60 per explant) were analyzed from each explant.
Protein isolation and Western blot
As described in Supplementary Materials and Methods.
Results
Isolation and characterization of AFCs
AFCs were collected from Cba.gfp E11.5-12.5 embryos and cultured in NIWA medium, seeded on an LIF producing modified feeder layer. Several alternative propagation culture conditions tested (Supplementary Table S1) did not support AFC survival and proliferation, and efforts to expand clonal cell populations by plating at clonal density on feeder layers in NIWA medium did not produce long-term cultures. In 3/5 experiments, clonal populations survived for only 30 days in vitro (see Supplementary Materials and Methods). In the feeder layer culture conditions, AFCs plated at high density could be maintained and expanded for at least for 18 passages (P18). AFCs preferentially grow at high density and proliferate slowly. Unselected AFCs show heterogeneous shapes after 3 days in culture: 51.8% ± 2.6% were small round cells, 42% ± 2.5% were mesenchymal-like cells, and 6.2% ± 2.2% were spindle-shaped cells (Fig. 1). Notably, we observed an increased number of round-shaped cells growing in aggregates to the detriment of the other 2 types from 2 weeks in culture.

AFCs in culture.
Previously, AFCs derived from mouse and human amniotic fluid that express the surface antigen c-kit (CD117) were able to give rise to lineages representative of the 3 embryonic germ layers cells under specific inducing conditions [22]. Here, we analyzed the c-kit+ population, in addition to the c-kit− population and unselected cells, to assess the retinal potential of AFCs. c-kit+ cells were isolated from the amniotic fluid of Cba.gfp embryos that ubiquitously express GFP (see Materials and Methods section). Approximately 1% of amniocytes collected at E11.5-12.5 age are c-kit+ [22]. To assess expression profile of AFCs, we performed reverse transcription (RT)-PCR analysis on cultured c-kit+ and c-kit− AFCs at P1 (passage 1) and P9 kept in propagation conditions (Fig. 2A, B), and on acutely isolated c-kit+ and c-kit− AFCs (data not shown), and compared these to that of ESCs, and E12.5 RPCs from the developing retina (RPCs) (Fig. 2C, D). Long-term maintenance of pluripotency in mouse ESCs is known to require expression of the stem cell pluripotency transcription factors Oct3/4, Sox2, and Nanog, and activation of tumor-related genes such as c-Myc and Klf4 [42]. Induction of pluripotency in somatic cells (induced pluripotent stem cells; iPSCs) was first achieved by combining 4 factors (Oct3/4, Sox2, c-Myc and Klf4) by the Yamanaka laboratory [43], and more recently different cocktails have achieved the same result. We found that all AFC populations, like ESCs, express Nanog and Oct3/4 as well as the tumor-related genes Klf4 and c-Myc (Fig. 2A, 2B left hand side). Unlike ESCs (Fig. 2C), either Sox2 was difficult to detect (c-kit− cells; Fig. 2A, upper left hand side; red arrow) or was not detected at all (c-kit+ cells). Notably, ESCs (Fig. 2C) and P1 AFCs (Fig. 2A) had very similar mRNA expression profiles, both for the pluripotency genes and retinal lineage genes (Fig. 2, right hand panels). Of particular interest was the observation that cultured P1 AFCs, and acutely isolated AFCs (data not shown) either c-kit+ or c-kit−, also expressed sets of transcription factors normally associated with a retinal lineage such as Et, Lhx2, Tll1, Six6, Otx2, Pax6, and Fgf15 (Fig. 2A, upper right hand side; differences between c-kit+ and c-kit− subpopulations are detailed in legend). The presumptive eye field in the anterior neural plate is defined by expression of a group of transcription factor genes, including Et, Lhx2, Tll1, Six6 Pax6, Six3, and Rax [44,45], all of which were detected by RT-PCR in embryonic E12.5 RPCs (Fig. 2D). In Xenopus embryos, simultaneous overexpression of these transcription factors and Otx2 is sufficient to promote eye development, including retinal cell differentiation [40]. Figure 2D shows expression of all the retinal markers tested, except the photoreceptor marker Nrl, in RPCs from the embryonic E12.5 retina.
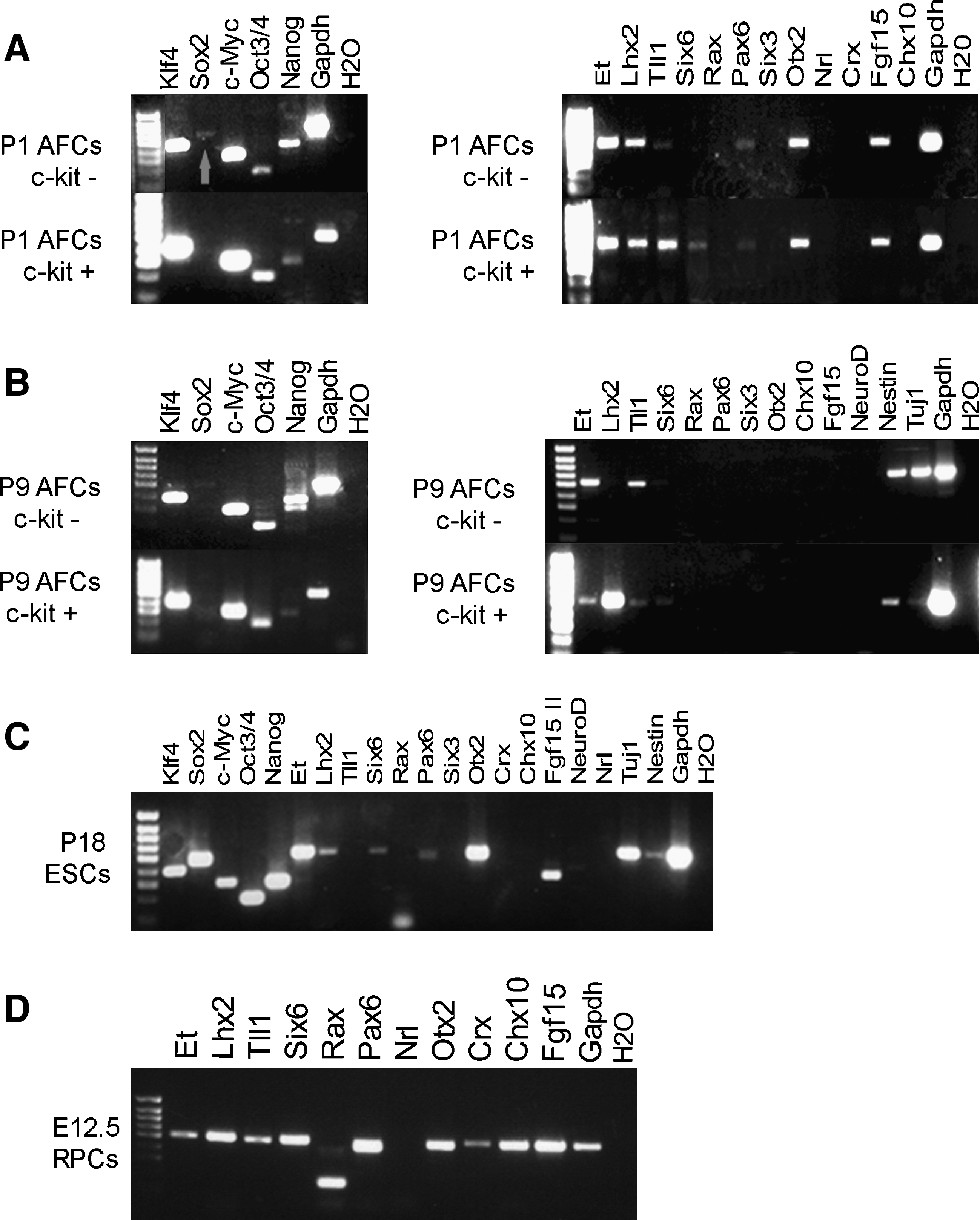
Comparative analysis of the RNA expression profile of AFCs, ESCs, and RPCs.
We found that if AFCs were maintained in ESC culture conditions, expression of the pluripotency markers Nanog, Oct3/4, Klf4, and c-Myc (Fig. 2B left hand side) and the neural markers Nestin and Tuj1 (Fig. 2B right hand side) were sustained (differences between c-kit+ and c-kit− subpopulations are detailed in Fig. 2 legend). However, expression of some of the retinal markers was lost over time; by P9 neither c-kit+ nor c-kit− subpopulations expressed Otx2, Pax6, or Fgf15, but Et, Lhx2, and Tll1 were detected (Fig. 2B right hand side).
RPCs proliferate and differentiate into photoreceptors in vitro, whereas AFCs do not
Since several studies have demonstrated the presence of different precursor cell lineages in the amniotic fluid [25,26,29,30,32,35], and our expression data indicated that at least some uncultured and early stage cultured AFCs may possess retinal potential (Fig. 2A, right hand side), we evaluated the possible presence of retinal progenitors within the amniotic fluid. We reasoned that if such cells existed then it might be possible to expand them in vitro using culture conditions that support the proliferation or differentiation of RPCs isolated from the embryonic retina. We tested AFCs derived from 3 retinal transgenic GFP reporter lines to provide a reliable readout of the presence of RPCs or photoreceptor precursors in the amniotic fluid before and after culture. In the developing retina, consistent with endogenous Fgf15 expression [46,47], the Fgf15.gfp transgene labels RPCs, whereas Nrl.gfp labels developing and mature rod photoreceptors [36] and Crx.gfp labels developing and mature cone and rod photoreceptors [38] (see also Supplementary Fig. S1 showing GFP expression in Fgf15.gfp retinal sections; the Fgf15.gfp transgenic line will be described in detail elsewhere).
We were unable to detect GFP-positive cells in acutely isolated AFCs from any of the 3 transgenic lines, as assessed by fluorescent microscopy analysis (data not shown). Embryonic RPCs from the developing retina were cultured in conditions that permitted their expansion and maintenance (retinal proliferation medium, see Materials and Methods section). Under these conditions, we could maintain proliferating GFP-positive retinal progenitors from E11.5-12.5 Fgf15.gfp retinae for at least 6 passages, and these cells maintained an appropriate retinal progenitor expression profile (Supplementary Fig. S2A–D). For simplicity, primary retinal neurospheres were used for comparisons in the remainder of this study. AFCs from Fgf15.gfp pregnancies were cultured in the same medium either in floating conditions or on poly-L-lysine, poly-L-lysine/laminin, or feeder layers. In these first 3 conditions AFCs formed aggregates that died after 10–14 days of culture, whereas AFCs grown on feeder layers survived for at least 1 month although their potential declined as demonstrated by RT-PCR analysis, which showed loss of expression of retinal markers (Supplementary Fig. S2E, F). In these experiments we did not observe Fgf15.gfp fluorescence.
We next adapted a photoreceptor differentiation protocol, which has previously been used to differentiate ESC-derived RPCs into photoreceptors [13,14]. For these experiments, we used the Nrl.gfp and Crx.gfp transgenic lines in which GFP provides a read out for the percentage of cells adopting a rod photoreceptor fate and a photoreceptor precursor fate, respectively. Nrl-positive photoreceptor precursors were generated at high efficiency by treating RPCs derived from E12.5 embryos for 4–6 days with 10 μM of the Notch signal inhibitor DAPT in the differentiation medium (Supplementary Fig. S2G–J). Further application of Fgf1, Fgf2, Sonic hedgehog, taurine, and retinoic acid did not increase the number of Nrl-positive cells. We observed spontaneous GFP expression in Crx.gfp-derived spheres after the first day in culture even under RPC proliferating conditions (Supplementary Fig. 2C, D), something not observed in Nrl.gfp-derived spheres. Conversely, when these same conditions were applied to acutely isolated AFCs derived from the amniotic fluid of Nrl.gfp and Crx.gfp embryos, they failed to promote GFP expression (data not shown). These data show that cultures of AFCs express some immature retinal markers, but that conditions to maintain bona fide RPCs or to favor photoreceptor differentiation are not sufficient to confer retinal properties and photoreceptor differentiation potential to the AFCs.
Retinal differentiation protocol effective on ESCs does not promote differentiation of AFCs
Recently, several related protocols have been developed to promote retinal progenitor fate from ESCs [10 –16]. Here the most efficient of these (for mouse) [13,14] was applied to expanded c-kit+ and c-kit− AFCs to test whether it was sufficient to induce retinal potential. AFCs, and ESCs as positive controls were grown according to the first part of the Takahashi laboratory protocol in floating conditions or on feeder layer (See Materials and Methods section; SFEB/DLFA; 10 differentiation step) to promote neuralisation and to produce RPCs. In this protocol, cultures are exposed to a differentiation cocktail composed of the Wnt and Nodal antagonists (DLFA), followed by fetal bovine serum and Activin A. We examined the cultures for the presence of neural progenitors (Nestin), differentiated neurons (Tuj1), and retinal progenitors markers (Et, Lhx2, Tll1, Six6, Rax, Pax6, Six3, Otx2, Crx, Chx10, Fgf15, and NeuroD) by RT-PCR. Both P8 c-kit− and P9 c-kit+ AFCs failed to survive in free floating conditions and died after 2 weeks in culture. The same cells seeded onto feeder layers survived, but their competence became reduced as demonstrated by RT-PCR analysis. Only Nestin and Lhx2 mRNA were detectable in treated P9 c-kit+ cells; none of the other RPC markers were found in either AFC subpopulation (Fig. 3A, left hand side). Moreover, c-kit+- or c-kit−-treated cells no longer expressed the pluripotency markers Oct3/4 and Nanog, although they still expressed c-Myc and Klf4 (data not shown).
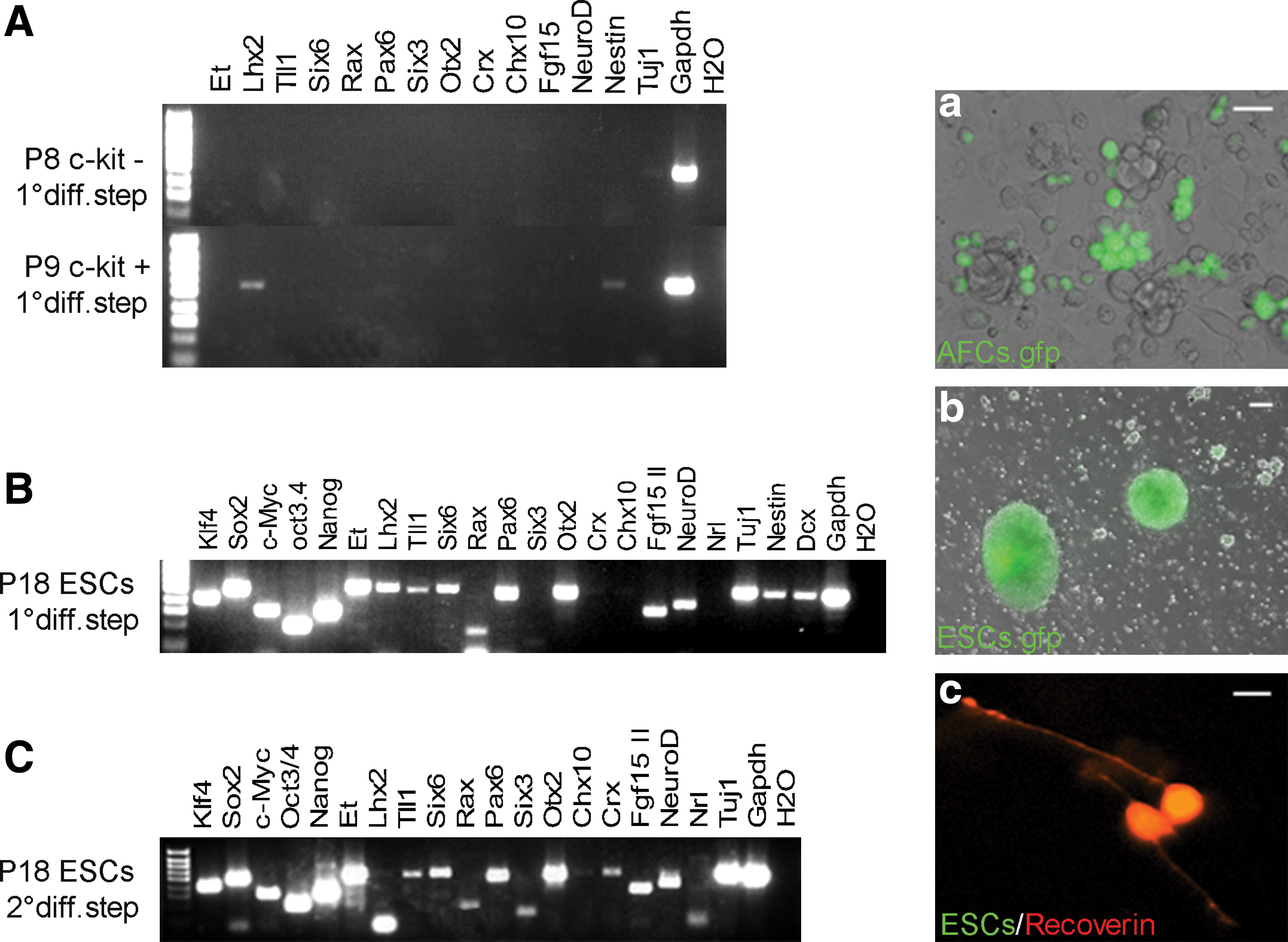
Comparison of retinal commitment of ESCs and AFCs after 2-step differentiation protocol.
By contrast, ESCs after the first differentiation step expressed the majority of the RPC markers tested (Fig. 3B, left hand side). After the second differentiation step the photoreceptor-specific markers Crx and Nrl, and Recoverin were induced in ESCs (Fig. 3C, c). These data show that the previously described ESC retinal differentiation protocol is not sufficient to commit the AFCs toward a retinal fate.
Co-culture experiments to induce RPCs and photoreceptor differentiation from AFCs
To investigate the hypothesis that other conditions, additional to the Takahashi protocol, may be required to commit AFCs to a retinal fate, we performed co-culture experiments. A combination of exogenous factors plus optimal explant conditions was explored [41]. Both c-kit+-sorted and unsorted AFC populations were tested. Retinae were provided from 2 stages of development: E11.5 when RPCs are proliferating and P4 during rod photoreceptor genesis. To assess retinal progenitor potential Fgf15.gfp AFCs were co-cultured with E11.5 embryonic retinal cells and either exposed to the SFEB/DLFA 10 differentiation protocol (see above) or left untreated (Fig. 4A). Similarly, to test photoreceptor potential Nrl.gfp, Crx.gfp AFCs were co-cultured with embryonic retinal cells with or without DAPT (Fig. 4B).
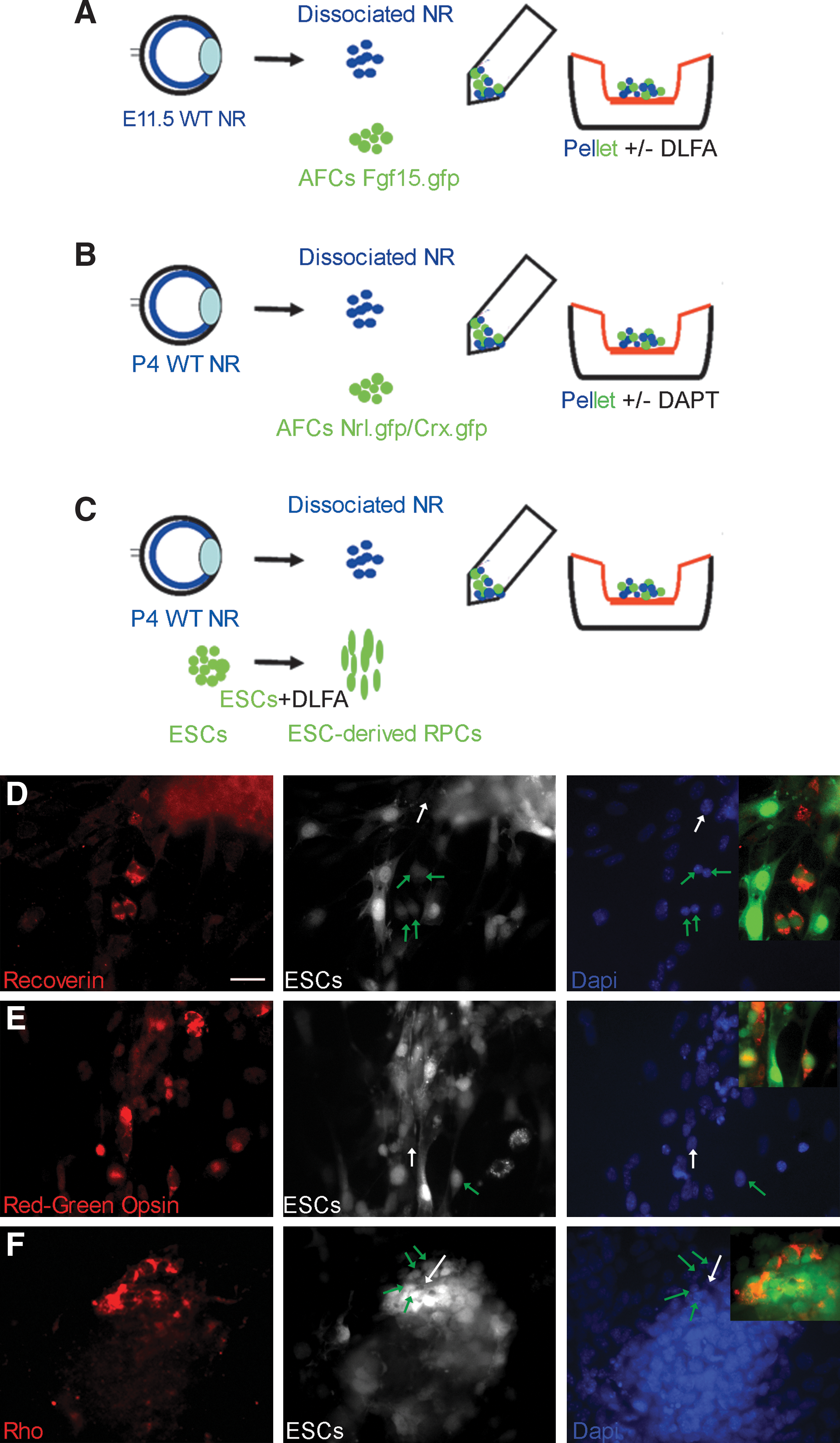
Retinal co-cultures with AFCs and ESCs.
Retinae and AFCs were centrifuged to produce reaggregated pellets as described by others [41]. The reaggregated cell pellets were cultured for 12 days on culture inserts (porous filter membrane) in the co-culture medium. No positive GFP signal was detected. In parallel, ESCs were grown in the SFEB/DLFA medium for 10 days to induce retinal progenitors, before being collected and induced to differentiate into photoreceptors using the same reaggregation co-culture assay [41]. By this assay, ∼20% of aggregates contained ESC-derived cells expressing photoreceptor markers (4/16 and 3/15; n = 2 experiments) as shown by the presence of positively stained Recoverin, Red-green opsin (4%–5%), and Rhodopsin (1%–2%) cells. The percentage of photoreceptors produced ranged between aggregates (Fig. 4C–F). These experiments provided no evidence in support of AFCs giving rise to RPCs and photoreceptors even with appropriate and proven environmental stimuli.
Transplantation of AFCs onto adult retinal explants
We next evaluated the migratory capability of AFCs transplanted onto adult retinal explants and compared integration levels with transplanted undifferentiated ESCs, and with in vitro differentiated Nrl-positive photoreceptor cells (Fig. 5A–D). Undifferentiated Cba.gfp-derived AFCs (5 × 105 per eye) were transplanted onto retinal explants and analyzed for integration of cells within the explant after 1 week in culture (Fig. 5A). For each transplanted cell type, 6 retinal explants were analyzed by sectioning. AFCs rarely migrated into the retinal outer nuclear layer (only 2 cell aggregates were observed within the 6 explants). The majority of the cells remained outside, along the edge of the explant (Fig. 5B; Supplementary Fig. S3). Surviving cells exhibited an immature morphology appearing as small round undifferentiated cells expressing proliferating markers (pH3; Supplementary Fig. S3B) and neural markers (Nestin; Supplementary Fig. S3E), but not expressing markers of mature neurons (Tuj1) nor photoreceptors (Rhodopsin and Recoverin) or the forebrain and retinal marker (Pax6) (data not shown). Conversely, in vitro differentiated Nrl.gfp cells transplanted onto adult retinal explants showed survival, migratory, and integration potential (Fig. 5C) and expression of a typical photoreceptor marker, Recoverin (data not shown). ESCs displayed extensive migratory capability, as aggregates (Fig. 5D) or as single cells, extensively colonizing the adult retinal explant. Thus, AFCs have a limited migratory capability compared with ESCs, and even when rare cells were observed within the retinal explant, they maintained an immature phenotype.
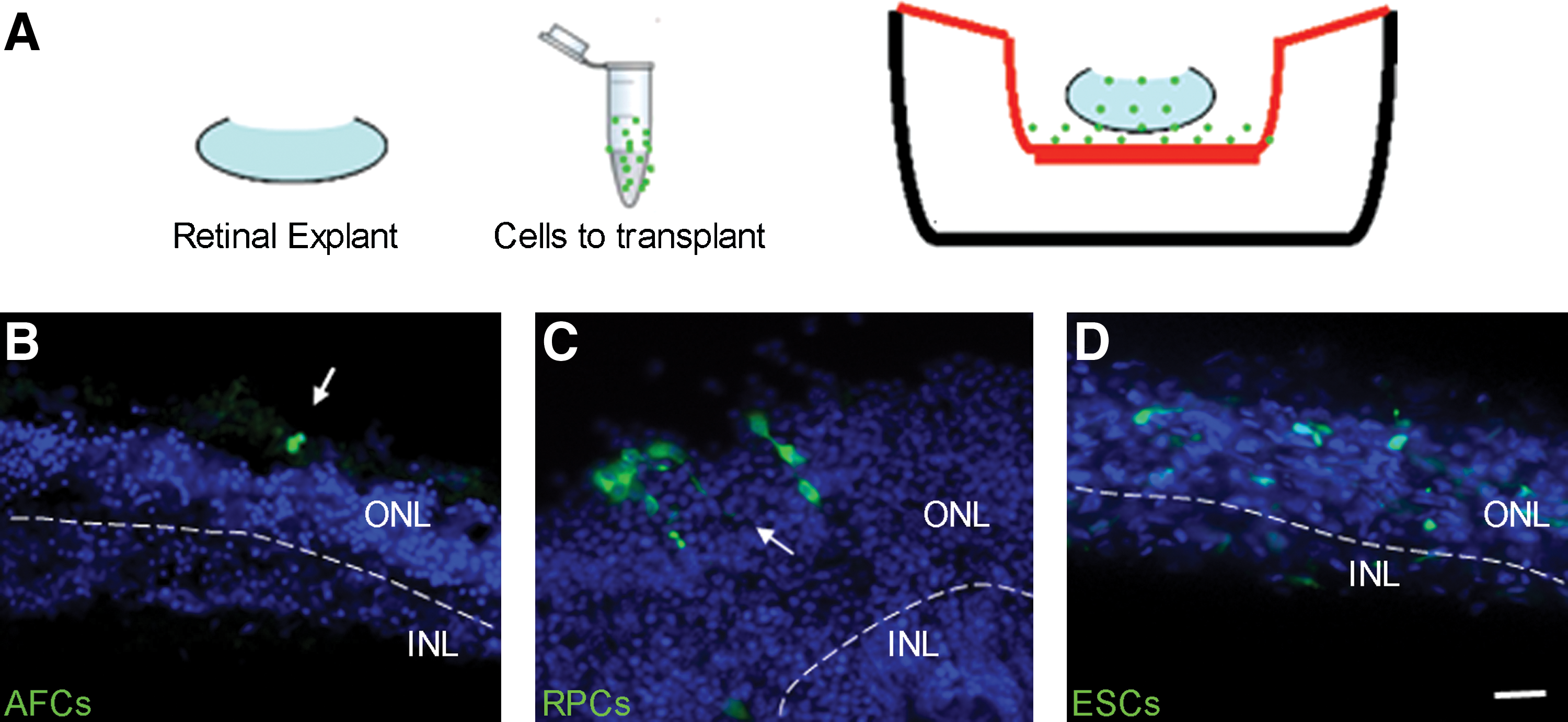
Analysis of AFCs, ESCs, and photoreceptor precursors transplanted onto adult retina explants.
AFCs only express the Oct3/4 isoform B
It was previously proposed that AFCs have neurogenic/ectodermal potential [22,24,29,32]. Here, we have shown that a number of protocols, each with proven efficacy for ESCs, were unable to induce a retinal fate in AFCs. Although our mRNA analysis indicated similarities between AFCs and ESCs (Fig. 2A–C) with respect to the expression of pluripotency markers, analyses using immunocytochemistry indicated a lack or reduced level of the corresponding proteins (Fig. 6A–D). Whereas cultured ESCs or RPCs transcribed and translated Oct3/4, Nanog, and Sox2 Nestin or Otx2, Pax6, and Tuj1, respectively (Fig. 6E–H; Supplementary Fig. S4), we were only able to detect the presence of the corresponding proteins in a small number of AFCs: Nestin 7%, pH 3 3% and rare Oct3/4 and Nanog-positive cells (Fig. 6A–D). With confocal analysis we observed the presence of unexpected Oct3/4 and Nanog-positive signals localized in the cytoplasm as well as in the nucleus (Fig. 7A–F). As a number of articles have reported the presence of 2 different Oct3/4-spliced variants either in human or in mouse [48,49] and as the Oct3/4 antibody we used recognizes both isoforms, we performed RT-PCR and Western blot analysis to identify the isoforms present in AFCs.

AFC and ESC protein expression profile.

Confocal and Western blot analysis of Oct3/4 and Nanog protein in AFCs.
The mouse Oct3/4 transcript lacking exon 1 and encoding isoform B is expressed in somatic tissues, and its function is still under investigation, whereas the isoform A that includes exon 1 is transcribed and translated in actively proliferating ESCs and in embryonic carcinoma [39,40]. RT-PCR analysis performed on c-kit+, or c-kit− AFCs revealed the presence of Oct3/4 A and B isoforms, which were also present in ESCs (Fig. 7G). As expected, Western blot performed on 2 different ESC lines (data shown only for E14Tg2A line) confirmed the presence of the Oct3/4 A isoform, but not the B isoform. By contrast, AFCs were positive for the B isoform but not for the A isoform (Fig. 7H). The same analyses were performed for the Nanog transcription factor, which, like Oct3/4 A, is involved in the safeguard of ESC pluripotency and in germline development [50]. No different isoforms have been described for Nanog so far. As already demonstrated by RT-PCR, ESCs and AFCs transcribed Nanog (Fig. 2A–C), but by Western blot analysis only ESCs translate the corresponding protein (Fig. 7I).
Finally, we conducted RT-PCR analysis for Fbx15, Rex1, and Utf1, 3 transcription factors known to be involved in pluripotency, and early lineage commitment [51 –56]. Rex1 deletion in mouse results in the loss of ability to differentiate into visceral endoderm, whereas Fbx15 is a novel F-box-containing protein activated by Oct3/4 and involved in the ubiquitin-proteasome pathway [51,52]. Utf1, encoding the undifferentiated embryonic cell transcription factor 1, is expressed mostly in the primitive ectoderm, and is able to increase cell reprogramming efficiency in induced pluripotent stem cell (iPS) protocols when transfected in place of c-Myc [51,53,54,56]. We found that although Rex1 and Fbx15 are transcribed in both AFCs and ESCs, Utf1 is only transcribed in ESCs, indicating a reduced ectodermal potential of AFCs (Fig. 7J). These differences in expression provide further evidence in support of the restricted potential of AFCs compared to ESCs.
Discussion
In recent years AFCs have been investigated as an alternative source of stem cells for therapeutic use as they avoid the ethical controversies associated with ESCs derived from surplus human embryos donated for research. Amniotic fluid obtained from routine clinical amniocentesis specimens has been shown to contain multiple types of progenitor cells [25,26,29,30,32,35], as well as pluripotent stem cells capable of differentiating into cell types representing each embryonic germ layer [22,24,27,28,31,32]. Recently, 2 different groups argued against the neural capability of human AFCs [57,58], following the proposal in an earlier study that dopaminergic neurons could be generated from AFCs based on limited immunocytochemistry and PCR analyses [22,24]. To date the majority of studies have focused on investigation of human AFCs and little use has been made of mammalian model systems to investigate AFC potential; in particular, murine genetic reporter systems provide a powerful analytical tool. In this study, we used mouse transgenic lines Fgf15.gfp, Crx.gfp, and Nrl.gfp that express GFP in developing retinal progenitors and photoreceptors. This novel approach based on the activation of retinal and cone/rod photoreceptor reporter lines has allowed us to assess the potential of AFCs to generate retinal cells.
We analyzed the capability of 2 different subpopulations of AFCs (c-kit− and c-kit+) to give rise to retinal progenitor and photoreceptor cells. AFCs were analyzed at E11.5-12.5 for 2 reasons: by this stage of mouse development optic cup formation is completed; the number of c-kit+ cells, demonstrated to be broadly multipotent by De Coppi et al., is maximal. AFCs are thought to derive from the amniotic membrane and from the developing embryos [33,34] and as neurogenic cells and the presence of neural stem cell markers have previously been described [31,34], we reasoned that retinal progenitors could be present in the amniotic fluid at the end of the optic cup formation. To compare the potency of AFCs with cells that genuinely have retinal potential, we employed protocols to differentiate bona fide retinal progenitors into photoreceptor cells and protocols to differentiate pluripotent ESCs into retinal progenitors in parallel.
Detection of several, but not all, of the eye field transcription factors in AFCs that are known to be essential for retinal development in Xenopus [45] provided some support for the existence of retinal potential in the murine amniotic fluid. However, we could not consistently detect expression of the retinally expressed Fgf15.gfp, Nrl.gfp, and Crx.gfp transgenes in acutely isolated cells or in retinal culture conditions, even though endogenous Fgf15 expression could be initially detected by RT-PCR. Two protocols were tested. The first was proven to maintain the proliferation of Fgf15.gfp-positive retinal progenitors in culture, and the second was proven to differentiate Crx.gfp and Nrl.gfp-positive photoreceptor precursors. Using these same culture conditions, AFCs expressing the transgenes were not generated. In conclusion, these experiments provide no evidence for the proposal that the amniotic fluid contains retinal progenitors.
Second, because of the numerous articles highlighting the presence of multipotent stem cells in the amniotic fluid [22,24,27,32], we addressed the possibility of committing putative broadly multipotent cells present into the amniotic fluid [22] to the retinal lineage. For this purpose, we adapted the Takahashi protocol to differentiate an ESC line (E14Tg2A) available in our laboratory. The protocol applied to ESCs allowed us to generate Crx/Nrl and Recoverin-positive cells in <3 weeks. In contrast, cultured c-kit+ and c-kit− AFCs treated with the same protocol, or with a modification where cells were differentiated on feeder layers, not only failed to induce the expression of the retinal markers but also reduced expression of the pluripotency markers Nanog and Oct3/4. To rule out the possibility that additional factors not present in the media were required to induce the differentiation of progenitors present in the amniotic fluid, we also tried to direct them toward a retinal fate by performing reaggregation co-culture assays between wild-type retinae at appropriate developmental stages and AFCs from Fgf15.gfp, Crx.gfp, and Nrl.gfp transgenic lines. Even in these experiments no GFP-positive cells were detected, despite induction of retinal differentiation in parallel control experiments using ESCs.
Migratory assays confirmed the different properties of AFCs compared with ESCs and with in vitro generated photoreceptor precursors. As previously demonstrated [7], immature photoreceptor cells at an appropriate developmental stage can migrate and integrate into the adult retina. ESCs also exhibit migratory capability, and migration has previously been reported after transplantation into the retina [59,60]. Rarely, AFCs expressing the proliferating marker pH3 and the neural marker Nestin migrated into the adult retinal explants. The absence of retinal/photoreceptor marker expression leads us to conclude that immature photoreceptors are not present in AFCs.
Our analyses showed that murine AFCs display only limited translation/stability of the pluripotency markers Nanog and Oct3/4 and also that they express a different Oct3/4 isoform compared with the pluipotency-associated isoform present in ESCs. These observations may explain the failure of retinal induction in our experiments and indicate that AFCs may have different potency compared with the ESCs. Notably, AFCs do not transcribe Utf1, indicating a reduction in ectodermal potential compared with ESCs. In wild-type mouse embryos, Utf1 mRNA is present in the inner cell mass, the primitive ectoderm, and the extra-embryonic tissues [54]. During the primitive streak stage, the induction of mesodermal cells is accompanied by the downregulation of Utf1 in the primitive ectoderm, highlighting the importance of this gene in the commitment of ectodermal-derived tissues. A further experimental approach also supports the conclusion that AFCs lack retinal potential. Attempts to make chimeric embryos by blastocyst injection of both c-kit+/− cells at passage 1 were not successful, and we found no evidence for AFC contribution to retinal tissue (Decembrini et al., unpublished observations). It proved impossible in the conditions tested to isolate single AFC colonies, and this means that we cannot completely rule out the possible existence of rare cells with retinal potential. Future studies could test alternative culture conditions such as low oxygen or overexpression of Utf1.
In conclusion, in this work we analyzed AFCs from at least 300 Nrl.gfp, Crx.gfp, and Fgf15.gfp transgenic embryos without obtaining any evidence for the presence of retinal/photoreceptor precursors in the amniotic fluid at the analyzed stages. Moreover, in experiments that took advantage of protocols effective at driving ESCs toward a photoreceptor fate and all analyses conducted to evaluate the AFCs expression profile indicate that mouse AFCs are cells with a restricted competence.
Recent work demonstrated the ability of uncultured AFCs to give rise to the hematopoietic lineage [26], renal cells [28], osteogenic tissues [31], and the myogenic lineage [27]. These cell types share a mesodermal or endodermal origin consistent with the idea that AFCs have a restricted competence. Recent progress in the iPS field suggests that AFCs could be a suitable source of donor cells to make iPS quickly and with a high frequency (100-fold higher efficiency was achieved using AFCs compared with fibroblasts cells) [61 –63] since the AFCs transcribe pluripotency genes, and somatic cells do not. In summary, investigations to date suggest that although the amniotic fluid is a potentially rich source of multipotent cells, these are not akin to ESCs and merit further investigation of their neural potential.
Footnotes
Acknowledgments
We gratefully thank Constance Cepko for providing the Crx.gfp mouse line, Anand Swaroop for the Nrl.gfp mouse line, Juan-Pedro Martinez Barbera for the LIF-producing MEF feeder line, and the UCL Institute of Child Health flow cytometry and confocal facilities for technical support. This work was supported by the Medical Research Council UK (G03000341 and G0901550); the Macula Vision Research Foundation; the Ulverscroft Foundation; the Wellcome Trust (082217); the Royal Society; the NIHR Biomedical Research Centre for Ophthalmology at Moorfields Eye Hospital and UCL Institute of Ophthalmology; and NIHR Biomedical Research Centre for Paediatric Research at Great Ormond Street Hospital for Children and UCL Institute of Child Health. R.A.P. is a Royal Society University Research Fellow; S.D. has a long-term EMBO Fellowship. J.C.S. is funded by Great Ormond Street Hospital Children's Charity.
Author Disclosure Statement
The authors declare no competing interests with the work presented in this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.