
Abstract
Brain damage through excitotoxic mechanisms is a major cause of cerebral palsy in infants. This phenomenon usually occurs during the fetal period in human, and often leads to lifelong neurological morbidity with cognitive and sensorimotor impairment. However, there is currently no effective therapy. Significant recovery of brain function through neural stem cell implantation has been shown in several animal models of brain damage, but remains to be investigated in detail in neonates. In the present study, we evaluated the effect of cell therapy in a well-established neonatal mouse model of cerebral palsy induced by excitotoxicity (ibotenate treatment on postnatal day 5). Neurosphere-derived precursors or control cells (fibroblasts) were implanted into injured and control brains contralateral to the site of injury, and the fate of implanted cells was monitored by immunohistochemistry. Behavioral tests were performed in animals that received early (4 h after injury) or late (72 h after injury) cell implants. We show that neurosphere-derived precursors implanted into the injured brains of 5-day-old pups migrated to the lesion site, remained undifferentiated at day 10, and differentiated into oligodendrocyte and neurons at day 42. Although grafted cells finally die there few weeks later, this procedure triggered a reduction in lesion size and an improvement in memory performance compared with untreated animals, both 2 and 5 weeks after treatment. Although further studies are warranted, cell therapy could be a future therapeutic strategy for neonates with acute excitotoxic brain injury.
Introduction
B
When survivors of preterm birth are assessed years later, they often perform less well in tests of cognition, attention, executive function, and perception than children born at term [13]. Despite the prevalence of perinatal brain damage and the personal as well as the economic burdens of long-term neurological morbidity, there is to date no effective treatment [14].
Cell therapy appears to be a promising neuroprotective and/or neuroregenerative strategy in various models of brain injury or disease (for a few examples, see [15 –19]). Over the last couple of decades, numerous studies have attempted to determine the efficacy and/or feasibility of transplanting stem cells or progenitor cells, including cells of both neural and non-neural origin, such as embryonic or mesenchymal stem cells, into the injured neonatal brain, to replace lost cells or to prevent damaged cells from dying [20,21]. Among these, neural stem cells (NSCs), self-renewing multipotent cells capable of generating the 3 main components of the nervous system—neurons, astrocytes, and oligodendrocytes—are present in the brain both during development and to a lesser extent in adulthood (for a review, see [22,23]), and have the advantage of being easy to isolate from mouse embryos and to propagate in culture in the form of multicellular aggregates called neurospheres [24]. However, despite the wide availability and use of these cells in animal models of injury, few studies have focused on their protective/regenerative effect in models of perinatal brain damage [25,26], and reports on functional recovery of the injured brain are practically nonexistent. Because clinical application of such cells as potential source of therapeutic cells in human is still impossible, it is of high importance to evaluate in adequate animal models the therapeutical potentiality of NSCs in vivo.
In the present study, we evaluated whether the implantation of neurosphere-derived precursors (NDPs) has a beneficial effect on the histopathological and behavioral outcome of ibotenate excitotoxicity in a neonatal mouse model of cerebral palsy. In this well-characterized model, ibotenate activates NMDA and metabotropic glutamate receptors and induces white matter cysts as well as cortical necrosis, similar to the lesions seen in newborns [27 –29].
Materials and Methods
All experimental protocols involving animals or human patients were carried out in accordance with local ethics guidelines after approval by the institutional review committee.
Animals
Wild-type mouse pups and embryos were obtained from pregnant Swiss mice (Janvier) housed at 24°C with a 12 h light/dark cycle and free access to food and water. To obtain green fluorescent protein (GFP)-positive NDPs, embryos were taken from C57BL/6 mice expressing GFP under an actin promoter (generously provided by Olaf Ninnemann, Institute of Cell Biology and Neurobiology, Charité, Berlin). Initial data were obtained using NDPs isolated from Swiss embryos and labeled with DiI staining. Because DiI is known to fade out, we have replicated the data with NDPs prepared from actin-GFP mice and extended the studies to longer time points (i.e P42). Animals used in the present study are detailed in Table 1.
Sizes of the different animal groups used in individual experiments performed at a given time are herein summarized. Numbers in parentheses indicate the different litters used to compose a given group.
GFP, green fluorescent protein; NDP, neurosphere-derived precursor; PBS, phosphate-buffered saline. Grey color indicated the infection time and therefore recapitulated the total number of animals used.
Isolation and culture of NDPs
Forebrains were dissected from E10.5 Swiss mouse or GFP mouse embryos as described previously [30,31] (Fig. 1). Cells were cultured in 10 mL Neurobasal medium (Gibco) supplemented with 20 ng/mL human recombinant fibroblast growth factor 2 (FGF2) (Sigma). The resulting neurospheres were dissociated into single cells by trypsinization and replated every 7 days. After dissociation, cell count and viability (>92%) were assessed by trypan blue exclusion. Culture media were supplemented with 10 nM pituitary adenylyl cyclase-activating polypeptide (PACAP; Calbiochem) and 1 ng/μL triiodothyronine (T3; Sigma) for 3 consecutive days before cells were harvested on in vitro day (DIV) 18 for implantation (see Supplementary Fig. S1; Supplementary Data are available online at
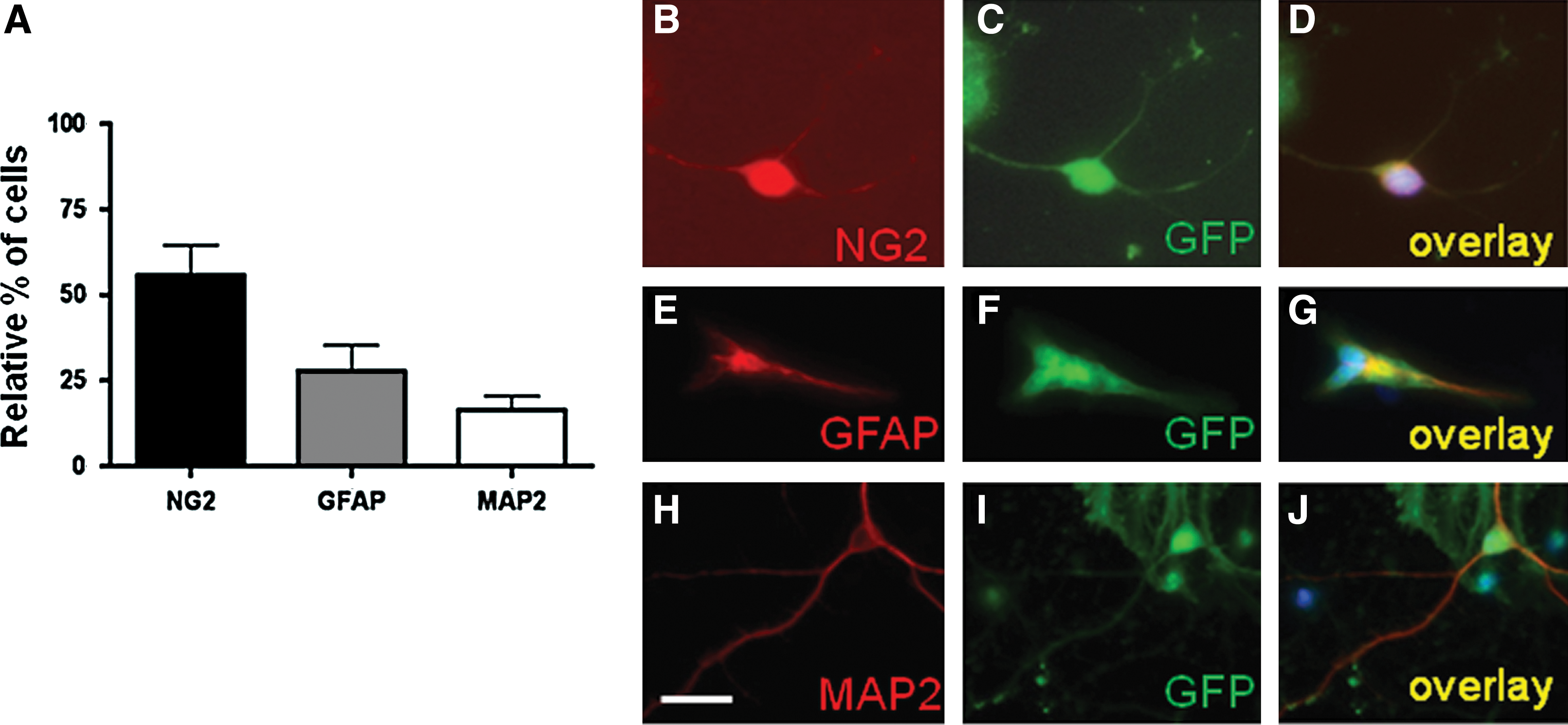
In vitro phenotype of neurosphere-derived precursors (NDPs) injected into ibotenate-lesioned brains.
RNA extraction and quantification of gene expression by real-time PCR
Total RNA was extracted on DIV18 according to a protocol previously described in detail [30]. Genes of interest were (1) the neuronal markers βIII-tubulin, nuclear receptor regulated 1 protein Nurr1 (dopaminergic neurons) and tryptophan hydroxylase (serotonergic neurons), (2) the astrocytic markers glial fibrillary acidic protein (GFAP) and S100, (3) the following markers for oligodendrocytes: 2′,3′-cyclic nucleotide 3′phosphodiesterase and platelet-derived growth factor receptor, alpha polypeptide, 2 transcripts specific to oligodendrocyte precursors; Notch and Delta1, known to promote the oligodendrocyte lineage; myelin proteolipid protein and myelin basic protein (MBP), expressed in mature oligodendrocytes, and (4) certain morphogens and neurotrophic (Table 2). Primer sets were designed using Oligo6.0 (Molecular Biology Insights Inc.) and M-fold software [32] (Table 2). The standard housekeeping gene used to normalize mRNA levels was glyceraldehyde-3-phosphate dehydrogenase (primer sequences in Table 2). Real-time PCR was set up using SYBR green-containing Supermix™ (Biorad) for 45 cycles of a 3-step procedure: a 20 s denaturation step at 96°C, a 20 s annealing step at 60°C, and a 20 s extension step at 72°C. Amplification specificity was assessed by melting curve and subsequent amplicon sequencing after subcloning into the TOPO-II vector (Invitrogen). Quantification was carried out using standard curves made from serial dilutions of control RNA samples, and transcript levels expressed as the ratio of the gene or interest to the housekeeping gene/genes.
Immunocytochemistry
On DIV18, neurospheres were dissociated into single cells by trypsinization and replated on glass coverslips coated with polyornithine. Sixteen days later (DIV34), cells were fixed with prewarmed 4% paraformaldehyde (PFA) for 20 min at room temperature. Fixed cells were identified/characterized by immunocytochemistry using the following primary antibodies: (1) NSCs: rabbit anti-Nestin (1:2,000; Sigma), (2) neurons: mouse anti-microtubule-associated protein 2 (MAP2, 1:10,000; Sigma), and mouse anti- neuron-specific nuclear protein [neuronal nuclei (NeuN), 1:1,000; Chemicon] (3) astrocytes: rabbit anti-GFAP (1:500; Sigma), (4) oligodendrocytes: mouse anti-NG2 (1:500; Sigma), mouse anti-O4 (1:500; Sigma). Cells were incubated in the appropriate primary antibody combinations overnight, and immunolabeling revealed with the following secondary antibodies for 2 h: Alexa Fluor 488 donkey anti-mouse IgG (1:500) or Alexa Fluor 488 donkey anti-rabbit IgG (1:500) (both from Invitrogen). Cells were then counterstained with DAPI (Boehringer) at room temperature for 1 min. Cell types were quantified by counting stained cells in 10 random fields per coverslip, and at least 3 coverslips per group, from 3 independent experiments.
Fibroblast culture
Cultured fibroblasts used as controls in our experiment were kindly provided by V. Paupe (Inserm U676, Paris, France) and originally derived from a forearm skin biopsy of a healthy male patient after written informed consent. Fibroblasts were cultured in Dulbecco's modified Eagle's medium (Gibco) supplemented with 10% bovine calf serum (Gibco) until they reached 80% confluency. Cells were then harvested by trypsinization and resuspended in 3–5 mL of phosphate-buffered saline (PBS) at a concentration of 3 × 105/mL.
DiI staining
NDP and fibroblasts not derived from actin-GFP mice were labeled with the red fluorescent dye DiI on DIV18 before infusion into the brain for later in vivo tracing. Cells were exposed to 7.5 mM DiI (Vybrant SE Cell Tracer DiI; Molecular Probes, Invitrogen) in their normal culture medium for 20 min at 37°C and then resuspended in PBS. The viability of NDPs after staining was evaluated by trypan blue exclusion in each experiment.
Excitotoxic lesions
Excitotoxic brain lesions were induced by a single intracerebral injection of 10 μg of the glutamate analogue ibotenate (5 μg/μL; Sigma) on postnatal day 5 (P5) isoflurane-anesthetized pups, as described previously [33,34] (Supplementary Fig. S2). This dose has been shown to consistently cause brain damage in P5 mice [35]. Ibotenate was injected into the neopallial parenchyma with a 26-gauge beveled needle adapted on 50 μL Hamilton syringe (Hamilton) and mounted on a calibrated microdispenser attached to a mechanically rigid holder. The needle was inserted 2 mm under the external surface of the scalp in the frontoparietal area of the right hemisphere, 2 mm from the midline in the mediolateral plane, and 3 mm from the junction between the sagittal and lambdoid sutures in the rostrocaudal plane. Histological analysis of brain tissue from 1,500 animals injected in our laboratory has previously confirmed that these visual coordinates result in highly reproducible injections that reach the periventricular white matter (P. Gressens, pers. comm.). Two pulses of 1 μL each were injected at an interval of 30 s. The needle was left in place for an additional 30 s. The correct positioning of the needle was verified by injecting some animals with toluidine blue. After the injections, pups were allowed to recover from anesthesia and were returned to their dams. Lesion induction by ibotenate was confirmed in a subset of pups (n = 5, see lesion size determination below).
Cell implantation
Within each litter, pups were first tattooed on the tail and then were randomly assigned to 1 of the following 6 treatment groups (see Fig. 3 and Table 1): (1) intracerebral ibotenate injection + DIV18 NDPs implantation (n = 12), (2) intracerebral ibotenate injection + fibroblasts implantation (n = 10), (3) intracerebral ibotenate injection + PBS injection (no cells implanted, n = 13), (4) intracerebral PBS injection + DIV18 NDP implantation (n = 12), (5) intracerebral PBS injection + fibroblasts implantation (n = 9), and (6) intracerebral PBS injection + PBS injection (no cells implanted; n = 8). At the given time, cells were implanted or PBS injected (negative control) 4 h after the ibotenate or first PBS (control) injection, intracerebroventricularly into the lateral ventricle contralateral to the ibotenate lesion, that is, the left lateral ventricle (A-P: 1 mm caudal to bregma, 0.5 mm left of the midline, and 2 mm below the surface of the scalp of isoflurane-anesthetized animals). The contralateral side was chosen to determine whether the implanted cells were capable of migrating toward the lesion site, rather than merely filling up the lesion when injected locally. For each pup, about 3 × 105 cells were slowly infused with a controlled flow of 100,000 cell/min using a NANOmite™ syringe pump (Harvard Apparatus), and the 26-gauge needle was slowly withdrawn after an additional 2 min. To evaluate the effect of later treatment initiation, pups were similarly treated on P8, that is, 72 h after ibotenate/vehicle injection, in the following groups: PBS + PBS (n = 10), ibotenate + PBS (n = 9), and ibotenate + NDP (n = 10).
Tissue processing
Injected mice were sacrificed by isoflurane inhalation followed by transcardiac perfusion of PBS (0.12 M TPO4, pH 7.4) before whole body fixation with 4% PFA in PBS. Brains were removed, postfixed overnight in 4% PFA in 0.12 M PBS at 4°C, and then cryoprotected in 10% sucrose in 0.12 M PBS for 2 days. Subsequently, brains were immersed in 10% sucrose and 7.5% gelatine in 0.12 M PBS for 1 h at 37°C, embedded in a block of the same solution for 1 h at 4°C, or flash-frozen in isopentane at −70°C, and stored at −80°C until further use. Ten-μm-thick parasagittal sections were cut using a cryostat, mounted on Superfrost Plus slides, and stored at −80°C. Brains dedicated to the determination of lesion size (see below) were fixed in 4% formaldehyde for 5 days and embedded in paraffin, and coronal sections of 16 μm were cut.
Lesion size determination
Lesion size was determined in randomly chosen pups from the ibotenate + PBS (n = 5), ibotenate + fibroblasts (n = 5), and ibotenate + NDP (n = 6) groups. Pups were sacrificed on P10, that is, 5 days after treatment, by transcardiac perfusion with 4% PFA, and the brains removed. After postfixation in the same solution for an additional 24 h at 4°C, the brains were dehydrated in alcohol and embedded in paraffin. Each brain was completely and serially sectioned from the frontal pole to the occipital lobes at 15 μm intervals in the coronal plane. Coronal sections were chosen because of the difficulty in accurately evaluating lesion extent along the radial axis, given the neuronal damage in the neocortical layers at the epicenter of the lesion, and our previous findings regarding the high level of correlation that exists between the maximal diameter of ibotenate-induced lesions along the radial and fronto-occipital axes [29]. An index of lesion volume was obtained after cresyl violet staining by multiplying the maximal diameter of the lesion along the fronto-occipital and sagittal axes by the number of sections in which the lesion was present and the thickness per section [28,36]. The size of the lesion in each brain was determined independently by 2 investigators blind to the treatment group.
NDP distribution and phenotype within the brain
To control the ability of NDPs to migrate toward the lesion, a subset of implanted animals was randomly chosen for brain analysis using either DiI or GFP to observe grafted cells on postinjury days 1, 2, 3, and 4.
To determine (1) the distribution of NDPs within the brain of ibotenate-lesioned and control pups and (2) the phenotype of NDPs at the level of the lesion, mice were sacrificed at 2 time points: at P10 (n = 10) and following behavioral assessment, at 6 weeks (n = 10) (see below). The following antibodies were used for immunohistochemical staining: (1) rabbit anti-Nestin (1:1,000; Sigma) for NSCs, (2) mouse anti-MAP2 (1:10,000; Sigma) and mouse anti-NeuN (1:1,000; Chemicon) for neurons, (3) rabbit anti-GFAP (1:500 dilution; Sigma) for astrocytes, and (4) mouse anti-NG2 (1:500 dilution; Sigma), mouse anti-MBP (1:500 dilution; Sigma) and mouse anti-O4 (1:500 dilution; Sigma) for oligodendrocytes, and (5) rabbit anti-caspase-3 (1:1,000; Sigma). Sections were incubated in the appropriate primary antibodies overnight, exposed to the secondary antibodies Alexa Fluor 488 donkey anti-mouse IgG (1:500) and Alexa Fluor 488 donkey anti-rabbit IgG (1:500) (Invitrogen), and stained with DAPI (Roche) at room temperature for 2 min. To assess transplanted mice for tumor formation, we autopsied the brain of the 2 transplanted females that died naturally before sacrifice, as well as the brains of 3 randomly chosen mice (1 female and 2 males) 6 months after transplantation. To evaluate abnormal proliferation of implanted cells, we also performed staining with rabbit anti-Ki67 (Invitrogen) on these and on randomly chosen younger mice (3 P10 and 3 P42).
Confocal microscopy
Confocal microscopy was performed using an oil-immersion objective (Plan-Apochromat 63x/1.4) on a Zeiss Observer inverted microscope equipped with an LSM 5 Exciter confocal scanning system (Carl Zeiss) and 488 and 543 nm lasers to observe the DiI or GFP signal (red and green, respectively) displayed by NDPs and the red signal displayed by various markers for stem cells, neurons, astrocytes, and oligodendrocytes. Images were processed using Adobe Photoshop CS software.
Behavioral assessment
The behavior of mice from the following groups was evaluated in an open field test [37,38] once per mouse at 3 weeks of age (Fig. 3): (1) intracerebral ibotenate injection + NDP implantation (n = 11), (2) intracerebral ibotenate injection + fibroblast implantation (n = 10), (3) intracerebral ibotenate injection + PBS injection (no cells implanted, n = 12), (4) intracerebral PBS injection + NDP implantation (n = 12), (5) intracerebral PBS injection + fibroblast implantation (n = 8), and (6) intracerebral PBS injection + PBS injection (no cells implanted; n = 7). The number of rears and contacts with objects as well as the total number of crossed squares and inside and outside crossings were scored. Motor activity included the number of rears and contacts with objects (testing exploratory behavior and sensorimotor activity, respectively) as well as the total number of crossed squares (motor activity) and the ratio of inside to outside crossings (inversely related to anxious behavior).
Memory function was studied in the same mice using the novel object recognition (NOR) test [39]. Indeed, under control conditions, mice explore a novel object more than a familiar object, and the degree of their preference for the new object is an indication of their memory of the familiar one [40 –42]. Tests were performed as described [39,43] with several modifications. Mice were allowed to habituate to an open field arena for 2 days (5 min per day) preceding the test. Two objects (A and B) were placed diagonally in the open field on day 3, and each mouse was allowed to explore them for 5 min. The time spent exploring each object individually and both objects were recorded, to evaluate levels of motivation, curiosity, and interest in exploring objects. Exploration of an object was defined as pointing the nose toward the object at a distance of <1 cm and/or touching it with the nose. Turning around, climbing, or sitting on an object was not considered exploration. After 30 min, object A was replaced by a replica (R: same shape but novel odor) and object B was replaced with a novel object, C, to test temporal memory. Alternatively, object B was replaced with a replica (R′) that was moved from its original position to test spatial memory. The mouse was again allowed to explore the objects for 5 min. Object recognition was scored by preferential exploration of the novel object using a discrimination index (DI) (novel object interaction/total interaction with both objects, ranging from 0 to 1; 0.5 = no preference) [39].
A subset of mice from the PBS + PBS (n = 7), ibotenate + PBS (n = 12), and ibotenate + NDP (n = 11) groups were re-tested with the NOR at 6 weeks of age to exclude the possibility that any recovery of behavioral function seen at 3 weeks was temporary. Pups included in the 72 h implantation procedure [PBS + PBS (n = 10), ibotenate + PBS (n = 9), ibotenate + NDP (n = 10)] were tested at 3 weeks of age. All behavioral assessments were carried out by 2 independent investigators blind to the treatment group.
Statistics
Data (mean ± standard error of the mean) were evaluated using the Student's t-test or standard analyses of variance with treatment as the between-subject factor. Statistical analyses revealed no significant influence of the litter of origin, gender, and weight of the animals on the results. The main within-subject effects and interactions are reported, together with P values based on the adjusted degrees of freedom.
Results
Phenotype of NDPs in vitro
The mouse embryonic forebrain NDPs used in this study presented the cardinal features of NSCs: (1) proliferation in FGF2 supplemented medium to form neurospheres, (2) differentiation into the different neural phenotypes (neurons, oligodendrocytes, and astrocytes), and (3) generation of secondary neurospheres after dissociation. After 2 weeks in culture, 55.8% ± 7.5% of the NDPs were NG2-positive cells, suggesting a high proportion of oligodendrocyte precursors. Neuroblasts (MAP2+ cells, 16.4% ± 4%) and astrocytes (GFAP+ cells, 27.8% ± 7.8%) were also detected (Fig. 1A–J). Quantitative PCR analysis confirmed the presence of the following mRNAs: (1) GFAP and S100 (astrocytes); (2) βIII-tubulin, Nurr1, and Tryptophan Hydroxylase (neurons); and (3) Notch, Delta1, 2′,3′-cyclic nucleotide 3′phosphodiesterase, platelet-derived growth factor receptor, alpha polypeptide, myelin proteolipid protein, and MBP (oligodendrocytes at various stages of differentiation; Supplementary Fig. S3).
NDPs migrate to the site of brain injury
NDPs were implanted into injured and control brains, contralateral to the site of injury (Fig. 2). As negative controls, fibroblasts or PBS solution were similarly injected. The fate of implanted cells was monitored 1, 2, 3, and 4 days after the ibotenate injury or control injection to analyze whether implanted cells were able to reach the lesion site. One day after implantation, cells had already migrated from the site of injection toward the contralateral hemisphere. Three days later, about 66% of the implanted cells were visible at the lesion site. In contrast, implanted fibroblasts were found only at the injection site and showed no evidence of migration (not shown).

In vivo fate of NDPs injected into ibotenate-lesioned brains. Injected NDPs migrate rapidly to the lesion site
NDP therapy reduces the size of the excitotoxic brain lesion
To study whether implanted NDPs reduced/attenuated ibotenate-induced brain damage, we determined lesion size in untreated and treated P7 and P10 mice. Mouse pups injected with ibotenate on P5 and treated with PBS or fibroblasts developed cortical and periventricular white matter lesions. The cortical lesion was typical, with dramatic neuronal loss in all neocortical layers and the almost complete disappearance of neuronal cell bodies along the ibotenate injection track (Fig. 3A, B). On the other hand, pups injected with ibotenate on P5 and implanted with NDPs showed a significantly smaller gray and white matter lesion than those treated with PBS or fibroblasts, both at P10, the usual time point for lesion size evaluation in this model (Fig. 3D), and, even earlier, at P7 (Fig. 3C), that is, 48 h after ibotenate injection, when the lesion was almost fully developed in control mice [36]. However, at this earlier time point, NDPs had not yet reached the lesion site, suggesting a significant and cell specific long-range effect of the implanted precursor cells, when compared with non- or fibroblast-implanted brains.
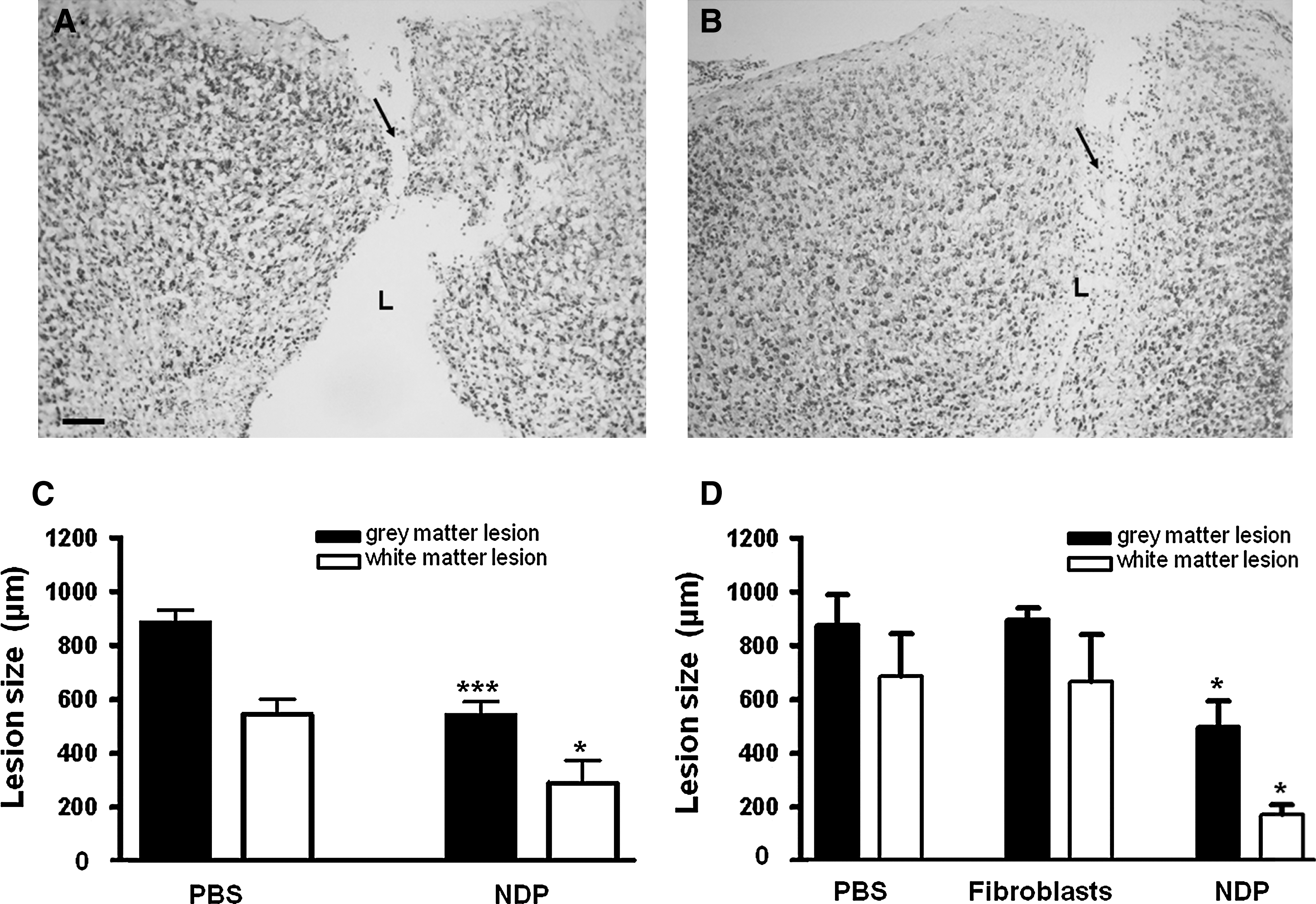
NDP implantation reduces the size of the ibotenate-induced gray and white matter.
To determine the fate of NDPs at the lesion site, we studied their phenotype using immunohistochemistry. Implanted DiI-positive or GFP-positive NDPs were localized at the level of the lesion in P10 mice (5 days after implantation). They were positive for Nestin (marker for NSCs or precursors) but not for GFAP, NG2, O4, MAP2, NeuN, or activated caspase-3, indicating that the NDPs reaching the lesion were still undifferentiated.
Improvement of memory performance through NDP implantation
To test the hypothesis that implanted cells could play a role in reducing the behavioral deficits caused by an ibotenate-induced brain injury, we performed behavioral tests in NDP-implanted and control 3- and 6-week-old mice (Fig. 4). Motor activity as evaluated by the open field test did not differ between the experimental groups. We further evaluated memory performance in our mice using the NOR test. In our study, the total amount of time spent exploring object A or B at time 0 did not differ significantly between control mice, lesioned NDP-implanted mice, and lesioned untreated mice. This indicates that mice had the same level of motivation, curiosity, and interest in exploring objects. Next, we compared the amount of time spent exploring the novel object as a fraction of the total object-exploration time (DI). All the ibotenate-lesioned mice injected with saline or fibroblasts showed a significant deficit in memory function (spatial and temporal) at 3 weeks (Fig. 4A). This deficit was not observed in mice implanted with NDP that performed as good as control PBS injected animals in memory tasks (Fig. 4A).
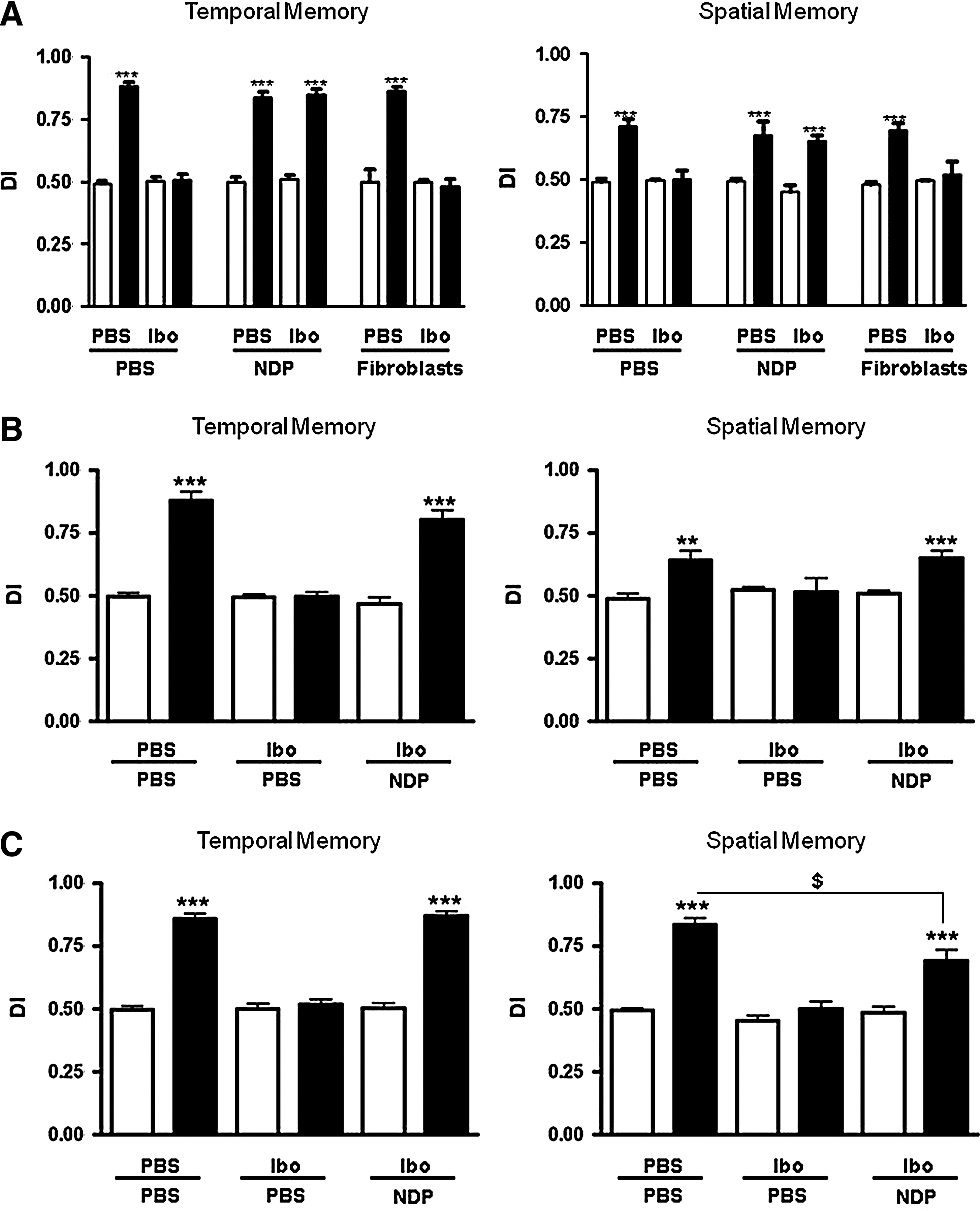
Improvement of memory performance after NDP implantation in ibotenate-lesioned mice.
To determine whether or not the beneficial effect of NDPs was transient, NOR evaluations were repeated with 6-week-old mice. Similar to our observations at 3 weeks, ibotenate-lesioned mice implanted with NDPs performed significantly better than those infused with PBS (Fig. 4B), suggesting that the behavioral improvement persisted throughout the developmental period. We observed a significant improvement in memory performance even when NDP implantation was delayed, that is, when cells were implanted 72 h after the excitotoxic lesion. The improvement in spatial memory in these animals was partial (DI = 0.78; Fig. 4C) and limited to the DIV18 PACAP + T3-treated NDPs, whereas the control DIV18 NDPs failed to improve the spatial memory defects (DI = 0.54, data not shown).
NDPs undergo cell death at the lesion site
To evaluate the long-term fate of implanted NDP, we determined their cellular phenotype in 6-week-old mice after behavioral evaluation. At this time point, the majority of the NDPs that had migrated to the lesion site were positive for neuronal (MAP2 and NeuN) or oligodendrocytic markers (NG2 and MBP). We were unable to find any NDP-derived GFAP-positive astrocytes (Fig. 5). Similar results were obtained when NDPs derived from actin-GFP mice were implanted (not shown). Since cell-type markers stained the entire cell rather than being localized in their respective antibody-specific compartments, we asked whether the implanted cells had retained their integrity. Activated caspase-3 staining did indeed show that all the Dil-labeled cells surrounding the lesion site were actively dying (Fig. 6G–L). Similar results were obtained with GFP-positive implanted cells (Fig. 6M–R). However, no actin-GFP cells were found positive for activated caspase at P10 when still in the vicinity of the ventricular zone where they have been originally grafted (Fig. 6A–F). The results indicate that the grafted cells retain their integrity until they have reached the lesion site where they finally undergo cell death.
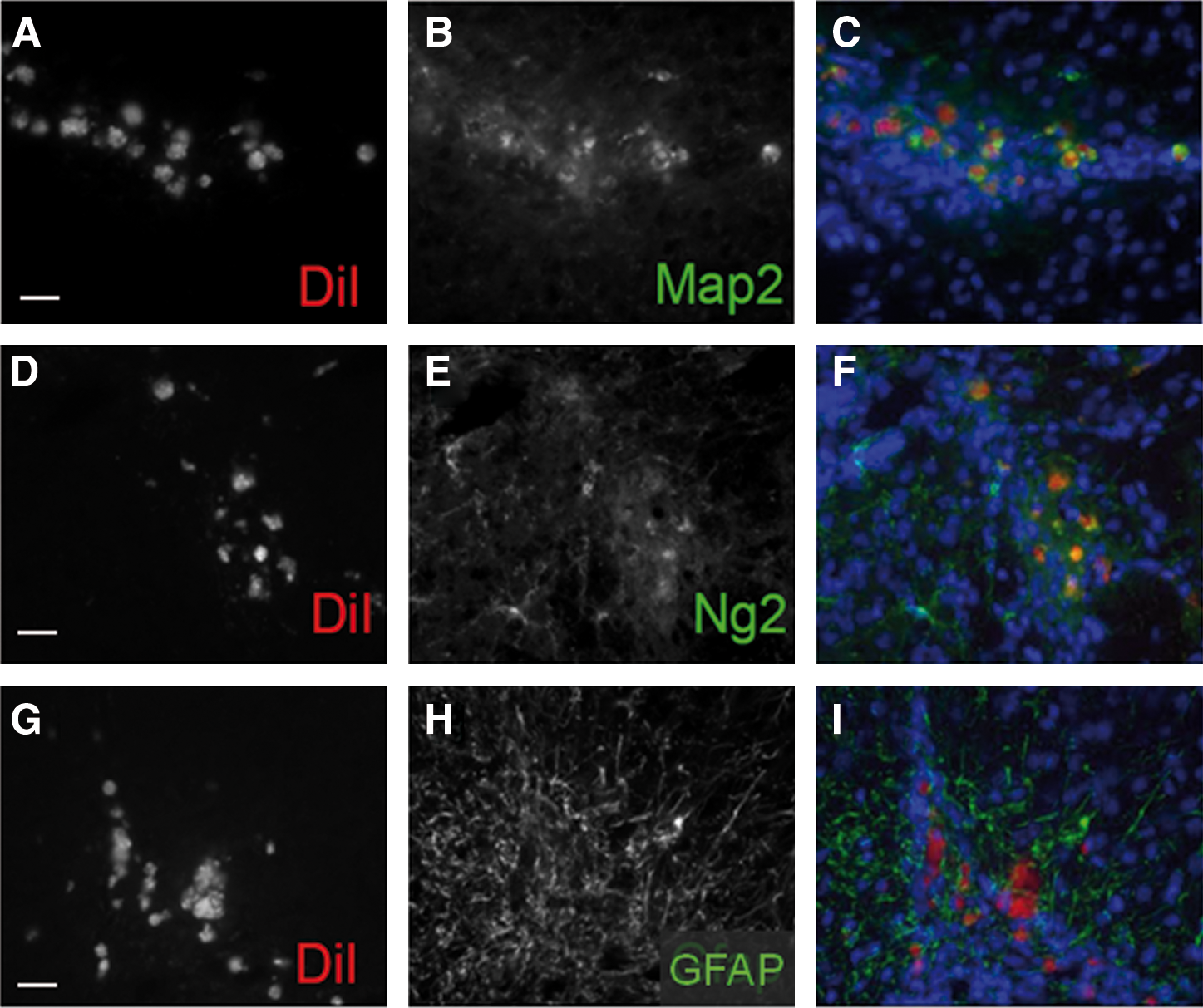
Phenotype of NDPs in vivo in 6-week-old ibotenate-lesioned mice. Cellular fate of NDPs at the lesion site determined using immunohistochemistry 5 weeks after implantation: DiI-positive NDPs (red) are immunopositive for the neuroblast marker MAP2
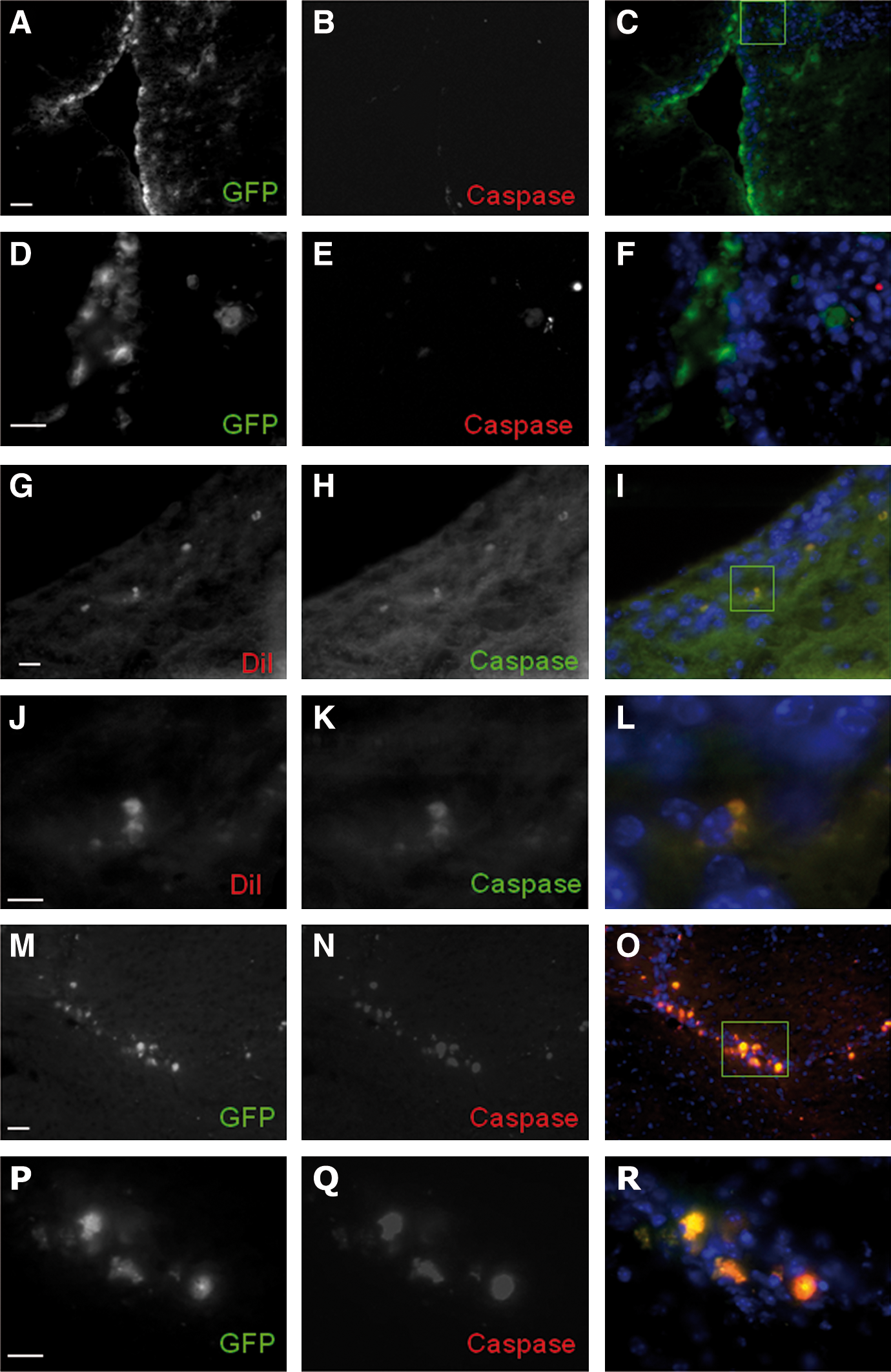
NDPs undergo cell death at the lesion site. Confocal microscopy shows no activated caspase-3 in the GFP-positive
No tumor formation was observed at the macroscopic or microscopic level in tested mice. Staining with anti-Ki67 antibodies did not detect any abnormal cellular proliferation (data not shown).
Grafted cells may produce a wide range of neurotrophic factors that could actively participate in functional recovery
Since implanted NDP cells did not survive after their migration to the injury site, we hypothesized that they could deliver neurotrophic factors, which in turn would enhance the recovery of endogenous cells after excitoxic brain injury. To test this, we cultured NDPs under medium conditions similar to those used for cell engrafting (FGF2 vs. PACAP+T3), before extracting total RNA and performing quantitative PCR to analyze the expression levels of several trophic factors that could promote recovery of the injured brain. Our results reveal that NDPs express several morphogenetic factors such as BMPs and shh; the neurotrophins NT3, NT4/5, and BDNF; neuropeptides, including VIP and PACAP (Fig. 7 and Table 3); and many other neurotrophic factors, including IGFs, NGF, GDNF, and CNTF (not shown). In addition, we observed that differences in culture conditions (FGF2 vs. PACAP+T3) resulted in a variation in expression levels of a few of them, including an increase in transcripts coding for the morphogens BMP6 and 7, the neuropeptide VIP, and the neurotrophic factors artemin, persephin, GDNF, and PDGF in the presence of PACAP+T3 (Fig. 7). Conversely, neurturin and LIF levels were found to be significantly decreased under these culture conditions. This indicates that NDPs express many growth factor transcripts in vitro and show that PACAP+T3 treatment is able to enhance this phenomenon. Although further experiments are required to fully demonstrate that implanted cells secreted recovery-promoting trophic factors in vivo, these results suggest that the implanted cells likely produce a wide range of neurotrophic factors that could actively participate in functional recovery.
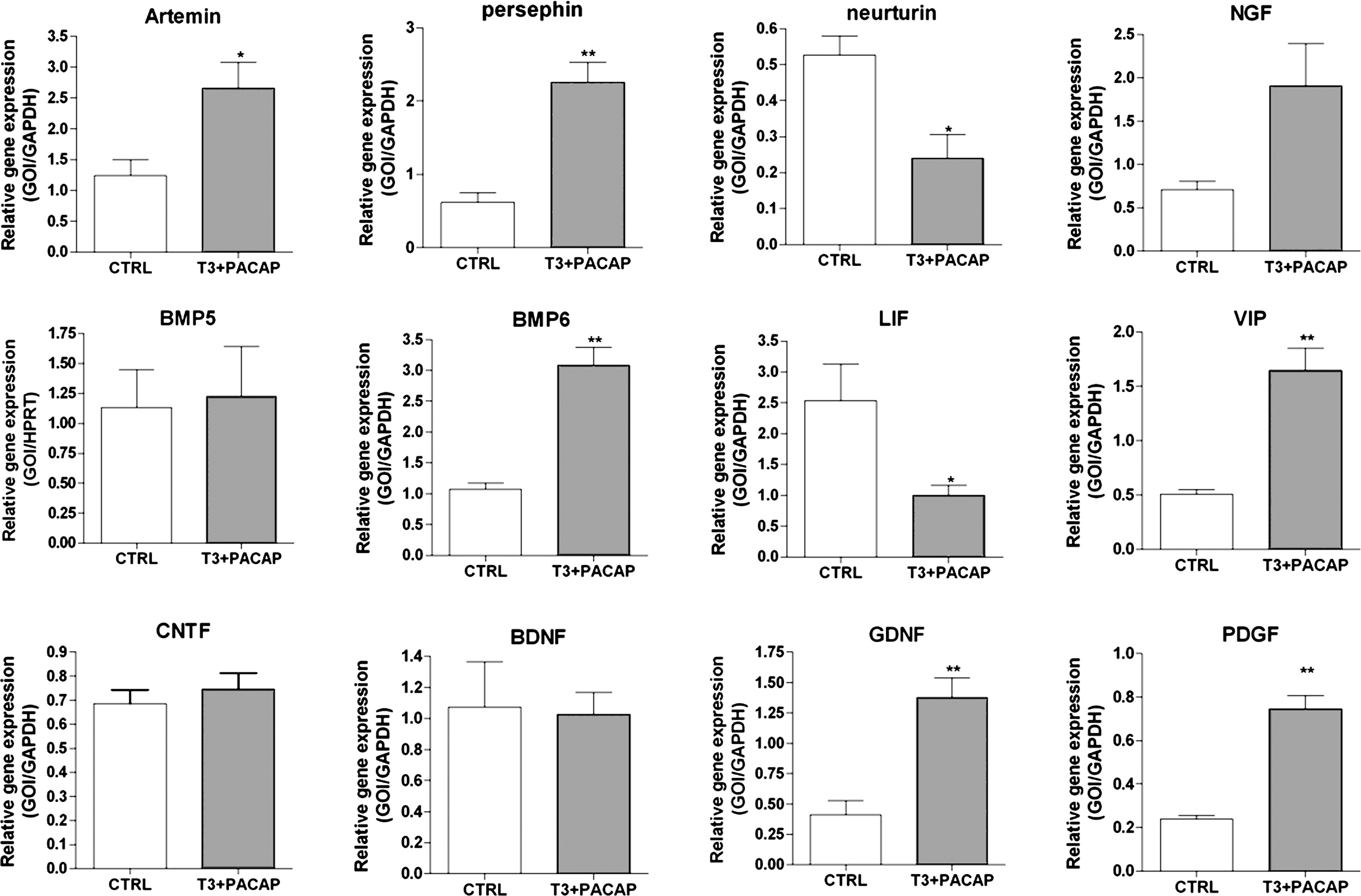
NDPs express a wide range of morphogenetic/neurotrophic factors in vitro. Gene expression (see Table 3 for details) in NDP cultures under standard conditions or with PACAP and T3 supplementation (see Supplementary Fig. S1) was quantified by real-time polymerase chain reaction (PCR) using total RNAs from cellular extracts. Data are presented as means ± SEM of the ratio of the gene of interest (GOI) to GAPDH. * P < 0.05, ** P < 0.01, *** P < 0.001; Student's t-test.
Data obtained in fibroblast growth factor 2 or in PACAP+T3 culture conditions shown on Fig. 6 are summarized using the following symbols: minor increase (↗) or decrease (↘); major increase (↗↗); no change (0).
PACAP, pituitary adenylyl cyclase-activating polypeptide; T3, triiodothyronine.
Discussion
In the present study, we evaluated the therapeutic potential of NDPs in acute excitotoxic injury of the neonatal mouse brain. The ibotenate-lesioned infant mouse is a classic excitotoxic injury model for the study of white and gray matter damage in humans, a major characteristic of cerebral palsy. The ibotenate-induced white matter lesion progressively increases in size during the first 24 h after the insult and then remains stable for 3–4 weeks, eventually being replaced by a glial scar [36]. Excitotoxicity-induced inflammation plays a central role in the formation of this lesion. Indeed, there is a significant increase in the density of activated microglia at the level of the lesion as soon as 4 h after ibotenate injection, before detectable cell death or axonal degeneration [36]. In addition, blockade of this microglial activation leads to significant neuroprotection [44]. The ibotenate-induced lesion mimics several key aspects of human perinatal brain damage, including its periventricular location, the initial cystic appearance, the secondary evolution of PWMD toward a glial scar, the associated appearance of gray matter damage, the deleterious effect of inflammatory cytokines, and the discrete ontogenetic window of sensitivity of the brain to damage [35,36]. These gray and white matter abnormalities tend to go hand in hand [7,12]: preterm infants who develop magnetic resonance imaging (MRI)-defined focal and diffuse white matter damage tend to have smaller cerebral cortical and deep gray volumes [45] than their preterm peers who do not have white matter damage. Expression of Nestin, a cytoskeletal protein involved in normal development and a marker for NSCs, is often reduced in the cortical gray matter during the subacute stage of PWMD but is increased at later stages, suggesting acute impairment with subsequent compensation leading to repair and plasticity [46].
We found that early (4-h) and late (72-h) NDP implantation significantly reduces the extent of the brain lesion after an acute excitotoxic injury to the neonatal mouse brain. Moreover, these cells are capable of migrating across the midline [47] toward the lesion site even when implanted contralateral to the lesion, similar to the long-distance migration of NSCs seen in a hypoxic-ischemic model of brain injury [48]. At the lesion site, they undergo transient differentiation into neurons and oligodendrocytes but not astrocytes. This strongly suggests that fate specification toward oligodendrocytic lineage is achieved by the culture conditions. Various protective and/or regenerative mechanisms by which NSCs could exert their effects have been described, and several of these could be involved in our model [49 –53]. Such mechanisms include, but are not limited to, an attenuation of central nervous system inflammation, the secretion of survival-promoting neurotrophic factors, the stimulation of the plastic response or neural activity in the damaged host tissue, and the restoration of synaptic transmitter release by providing/promoting local re-innervation. The latter mechanism that refers to the integration of NDPs into existing neural and synaptic networks to re-establish functional afferent and efferent connections is obviously impracticable in our model.
Therefore, our data address the legitimate question whether NDP implantation improves cognitive defects through a neuroprotective or a neuroregenerative mechanism or again through a simpler mechanism of wound healing. In this context, lesion can first be identified under the microscope as early as 4 h after injection, and the lesion size remains stable from P6 to P30 [36]. In our study, at P7, the lesion was already smaller in NDP-treated mice than in controls, and remained so at P10. At the latter time point, NDPs were still undifferentiated, suggesting that wound healing is a plausible mechanism to reduce brain lesion size. In addition, implanted NSCs have been reported to remain undifferentiated in other brain injury models (eg, for 30 days in ref. [53]). At P42, corresponding to the last behavioral evaluation (ie, in 6-week-old mice), implanted cells were undergoing cell death. Our data thus support the hypothesis that in this model, NDPs exert their beneficial effect through wound repair and neuroprotection rather than neuron and oligodendrocyte regeneration. However, in absence of any therapeutic intervention, the wound is known to heal but also to result in cognitive defects, suggesting that wound healing per se is not sufficient for full recovery.
This effect is probably mediated by a non-local immunomodulatory or trophic mechanism, since NDPs need about 9 days to reach the lesion site. For instance, a decrease in microglial activity has previously been shown to be induced by grafted stem cells (for review, see [54]). The active secretion of neurotrophic factors by NDPs (as shown in Fig. 7) may also play a key role in the present study. Indeed, we demonstrate expression of mRNAs for several morphogens and neurotrophic factors known to be associated with neuroprotection, such as BMPs, members of the neurotrophin family, GDNF, CNTF, and VIP [33,35,55 –57], or reported to be beneficial to oligodendrocyte development, such as PDGF, PACAP, IGF1, and GDNF [58 –61]. In addition, we observed that preconditioning medium containing PACAP+T3 further enhanced expression of VIP, BMP6, artemin, persephin, and GDNF compared with medium containing FGF2. This may have played a role in the improved neuroprotective activity of NDPs when grafted 72 h after injury.
In parallel with the reduction in lesion size, injured mice implanted with NDPs displayed a persistent and marked improvement in temporal and spatial memory, performing significantly better in the open field and NOR tests at 3 and 6 weeks of age than littermates that received intracerebroventricular injections of PBS or fibroblasts. The behavioral impairment seen in control mice was no longer apparent at 3 weeks in injured mice that were implanted with NDP, even when the implantation was delayed to 72 h after ibotenate injection. This result is quite intriguing because memory performance in adult mice is mainly controlled by adult neurogenesis occurring in dentate gyrus. Because we did not observe changes in adult neurogenesis in both P10 and P42 mice after NDPs implantation (using Ki67 immunostaining; data not shown), we have no further explanation for such a phenomenon, although 2 recent reports using a different experimental brain damage paradigm have described similar behavioral improvements in 9-day-old mouse pups and 7-day-old rat pups when subjected to intracerebral implantation of mesenchymal stem cells [62,63]. Our results suggest that even in the absence of the survival or differentiation of implanted cells or enhanced adult neurogenesis in NDPs implanted P10 and P42 mice (data not shown), the functional impact of brain injury in neonates can be modified.
The use of preorientated neural progenitors such as oligodendrocyte precursors for the treatment of neurological diseases characterized mainly by myelin loss is slowly becoming a realistic option [64,65]. At 6 weeks, implanted NDPs expressed neuronal and oligodendrocytic markers but were also undergoing cell death, as shown by their immunoreactivity for activated caspase-3. Our current data do not allow us to state conclusively whether the in vitro exposure of NDPs to PACAP and T3 modifies their in vivo fate or whether this plays a role in the behavioral improvement shown by implanted mice. This supplement cocktail was chosen because previous studies have shown that both PACAP [30,58] and T3 [66,67] are important extrinsic cues regulating the proliferation and differentiation of oligodendrocyte progenitors in vitro and in vivo.
Both embryonic stem cell and mesenchymal stem cell implants from various sources have been shown to reduce lesion size in animal models of neonatal brain damage, but few if any of these cells differentiate into neural cells in vivo [21]. Besides, tumor development remains a serious concern with regard to embryonic stem cell use [68]. We did not observe tumor formation in a subset of our treated mice, although more animals will need to be analyzed to confirm these data.
From a clinical and ethical point of view, treating infants suffering from brain insults using NSCs is still highly questionable. However, in view of future therapeutic interventions, we also chose to implant NDPs 3 days after lesion induction as a proof of concept. The efficacy of a therapeutic intervention is directly related to the rapidity with which it is implemented in acute human brain disorders such as stroke [69]. Mice receiving NDPs 72 h after the ibotenate injury showed a partial recovery of impaired memory function. This suggests that intervention at a later time point after an acute excitotoxic injury could also be effective. A better definition of this therapeutic window, as well as the pathophysiological mechanisms involved, could lead to optimal behavioral improvement.
In this study, we have demonstrated the occurrence of functional recovery induced by precursor cell therapy in a neonatal model of cerebral palsy. In humans, the potential benefits of such an approach could extend to other neurological disorders of childhood, glutamate excitotoxicity being the final common pathway for many diseases such as hypoxic-ischemic encephalopathy, stroke, herpes virus encephalitis, and shaken baby syndrome [70]. However, further studies are needed to fully understand the mechanisms involved in stem cell-mediated neuroprotection before cell therapy can be considered for acute diseases of the developing human brain.
Footnotes
Acknowledgments
This work was supported by INSERM, Université Paris-7, Fondation pour la Recherche Médicale, the ELA foundation (ELA 2007-024I4), APHP (Interface contract to P. Gressens), the Niemann-Pick Association—Italy, and the PremUp Foundation. The authors thank Olaf Ninnemann and Robert Nitsch, Institute for Cell Biology and Neurobiology, Charité, Berlin, for providing the actin promoter GFP mice. We are very grateful to Gap Junction for their proficient English editing service and useful comments (
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.