
Abstract
Bone tissue defects caused by trauma and disease are significant problems in orthopedic surgery. Human embryonic stem cells (hESCs) hold great promise for the treatment of bone tissue disease in regenerative medicine. In this study, we have established an effective method for the differentiation of osteogenic cells derived from hESCs using a lentiviral vector containing the transcription factor Cbfa1. Differentiation was initiated in embryoid body formation of Cbfa1-expressing hESCs, resulting in a highly purified population of osteogenic cells based on flow cytometric analysis. These cells also showed characteristics of osteogenic cells in vitro, as determined by reverse-transcription (RT)–polymerase chain reaction and immunocytochemistry using osteoblast-specific markers. We also evaluated the regenerative potential of Cbfa1-expressing cells derived from hESCs (hESC-CECs) compared with hESCs and the osteogenic effects of bone morphogenic protein-2 (BMP2) encapsulated in thermoreversible hydrogel in vivo. hESC-CECs were embedded in hydrogel constructs enriched with BMP2 to promote bone regeneration. We observed prominent mineralization and the formation of nodule-like structures using von Kossa and alizarin red S staining. In addition, the expression patterns of osteoblast-specific genes were verified by RT–polymerase chain reaction, and immunohistochemical analysis revealed that collagen type 1 and Cbfa1 were highly expressed in hESC-CECs compared with other cell types. Taken together, our results suggest that encapsulation of hESC-CECs with BMP2 in hydrogel constructs appears to be a promising method to enhance the in vitro osteoblastic differentiation and in vivo osteogenic activity of hESC-CECs.
Introduction
B
In a previous report, we have shown that the differentiation of osteogenic cells from hESCs can be achieved by coculture with primary human bone-derived cells through osteoblast-secreted morphogenetic factors. These cultures provide survival signals that result in the enhanced expression of osteogenic genes, which ultimately leads to cartilaginous nodule formation [22,24]. Although this coculture system increased the efficiency of osteoblastic differentiation, it was difficult to obtain highly purified osteogenic cells derived from hESCs. To improve the specificity and purity of this technique, hESCs can be genetically manipulated with specific osteogenic genes during differentiation [25]. A lentiviral vector system can be used to create stable genetic modifications of mammalian embryonic stem (ES) cells [26 –28]. Using a lentiviral vector system, we have previously shown that the elongation factor (EF)-1a promoter can drive gene expression more efficiently than can the CMV promoter in hESCs [29]. Cbfa1, a high-mobility-group domain transcription factor, is expressed in osteoblast cells. Cbfa1 functions downstream in the bone morphogenic protein (BMP) signaling pathway and is an essential transcriptional regulator of osteoblastic differentiation and bone formation [30,31]. Cbfa1 overexpression upregulates osteogenic gene expression in a wide range of cell types. Thus, this gene has the potential to promote the production of bone-specific products and extracellular matrix, which could ultimately result in the generation of new bone tissue [32 –38]. However, to our knowledge, the ability of Cbfa1 to affect hESC differentiation has not yet been investigated.
In the present study, we first investigated whether Cbfa1 overexpression during hESC differentiation could produce pure osteogenic cells with a high yield. We successfully generated highly purified osteogenic cells that could express Cbfa1-Venus from differentiating human embryoid bodies (hEBs), as determined by flow cytometry. We found that Venus-expressing cells possess characteristics similar to those of osteogenic cells in vitro. Next, we tested whether the osteogenic activity of hESC-CECs could be improved by a carrier scaffold containing BMP-2 (BMP2). Previous studies have indicated that Cbfa1 and BMP2 have distinct but complementary roles in osteogenesis and that their combined actions may be necessary for optimal bone formation [39,40]. Isolated and expanded hESC-CECs were encapsulated with BMP2 in hydrogel and implanted into the back subcutis of mice. The osteogenic potential of transplanted encapsulated hESC-CECs was examined by reverse-transcription (RT)–polymerase chain reaction (PCR), alkaline phosphatase activity assays, and immunohistochemistry of the implanted tissues.
Materials and Methods
Lentiviral vector constructs and virus production
The lentiviral vector constructs used in the present study were donated by Dr. Hiroyuki Miyoshi (Rinken Tsukuba Institute, Ibaraki, Japan). We constructed the SIN-EF-1a-Flag-Cbfa1-IRES-Venus lentiviral vector for osteoblastic differentiation (Fig. 1A). The virus stocks were produced by transient transfection into 293T cells. Briefly, 293T cells were transfected with the following plasmid DNAs using calcium phosphate transfection buffer: the envelope plasmid (pRSV-REV harboring the gene encoding VSV-G), the packaging plasmid (pCAG-HIVgp), and the transfer vector. The medium was replaced after 12–16 h with Dulbecco's modified Eagle's medium (Sigma) supplemented with 10% FBS (Gibco BRL). The medium containing viral particles was collected 48 h after transfection and concentrated at 50,000 g at 4°C for 4 h.
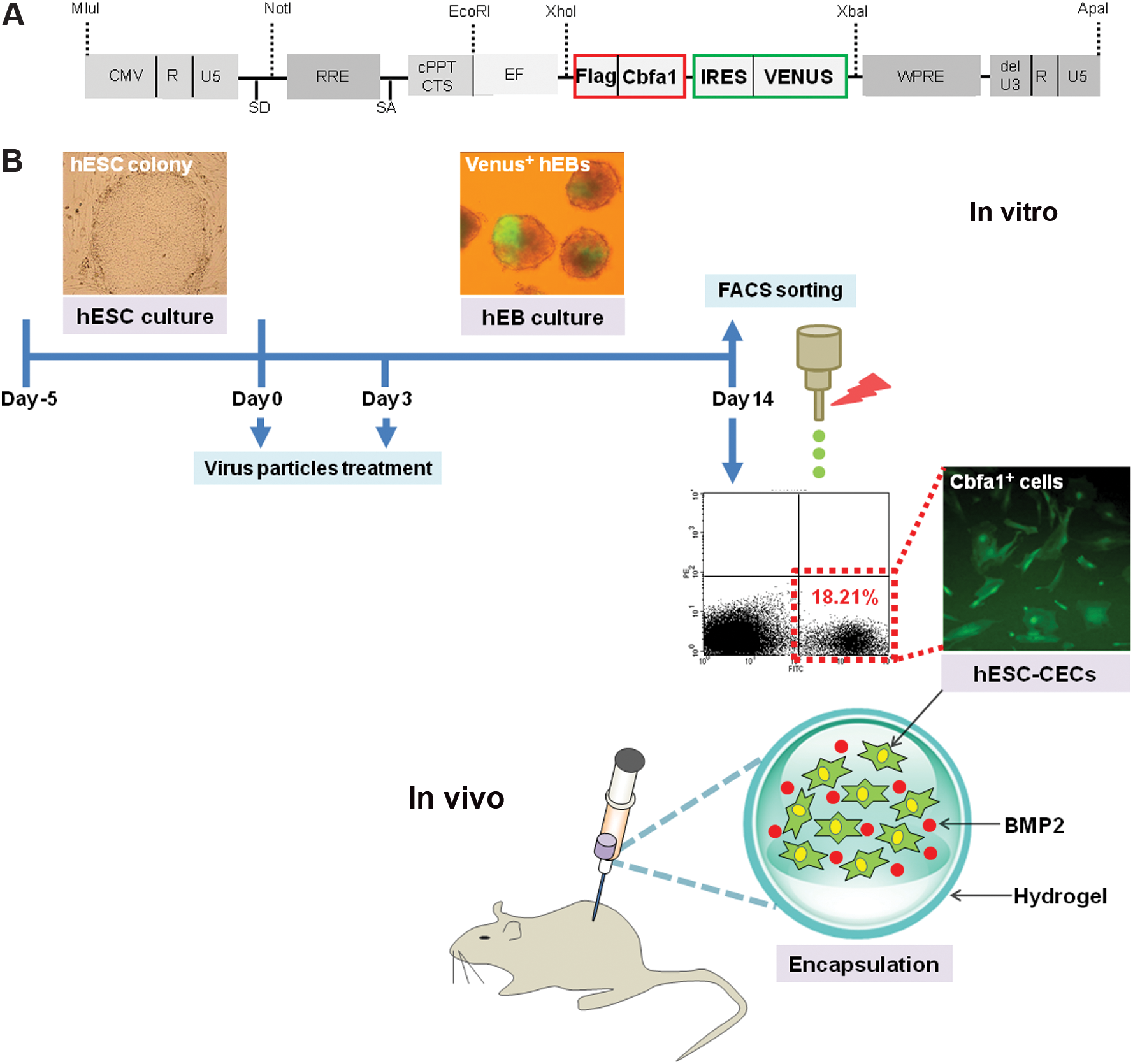
Cbfa1-overexpressing osteogenic cells derived from human embryonic stem cells.
Transduction of the Cbfa1 gene into hESCs
Undifferentiated hESCs (CHA3-hESC cell line) were cultured as previously described [24,29,41]. For hESC differentiation, hESCs were cultured in suspension in basic fibroblast growth factor-free hES cell culture medium. To transduce the Cbfa1 gene into hESCs, viral particles expressing Cbfa1-driven Venus were applied twice during hEB differentiation (Fig. 1B). To obtain pure hESC-CECs, transduced cells were sorted using a FACSVantage flow cytometer (BD Bioscience) at day 14 hEB.
Immunocytochemistry and histochemistry
To detect mineralized nodules, cells were fixed in 4% paraformaldehyde and incubated for 10 min in 1% alizarin red S (Sigma) solution. To detect osteoblast-specific markers, cells were fixed in 4% paraformaldehyde and permeabilized with 0.1% Triton X-100 in phosphate-buffered saline (PBS; Gibco). After treatment with 1% normal goat serum, the cells were incubated with collagen type 1 (COL1; Sigma) and osteocalcin (OCN; Biogenesis; Poole, England) at 4°C for 24 h. After being washed, the bound primary antibodies were detected by incubation with fluorescein isothiocyanate (FITC)-conjugated (Santa Cruz Biotechnology) or rhodamine-conjugated secondary antibodies (Molecular Probes), respectively. The slides were then mounted with Vectashield mounting solution containing DAPI (Vector Laboratories). Images were analyzed with a digital camera (DXM1200C; Nikon) and a fluorescence microscope (ECLIPSE TE 2000; Nikon). For von Kossa staining, fixed and washed cells were incubated in a 2% silver nitrate (Sigma) solution with simultaneous exposure to a 60-watt ultraviolet light bulb. Following a 1-h incubation at room temperature and washing with distilled water, the reaction was stopped by the addition of a 3% sodium thiosulfate (Sigma) solution for 5 min at room temperature. The cells were incubated in an alkaline dye mixture for 15 min at room temperature and then counter-stained with 0.1% nuclear fast red. For the immunohistochemical assays, cells embedded in hydrogel sections were fixed in 4% paraformaldehyde solution, dehydrated, and embedded in optimal cutting temperature (OCT) compound. Nonspecific binding was blocked with normal horse serum diluted 1:10 in 0.3% bovine serum albumin. Sections were incubated with COL1 (Sigma) or Cbfa1 (Santa Cruz Biotechnology) antibodies for 24 h at 4°C. After being washed in PBS, the sections were incubated with FITC-conjugated secondary antibodies (Amersham) and then stained with a 1:100 dilution of DAPI (Molecular Probes) for 5 min. An isotype control monoclonal antibody was used to estimate the levels of nonspecific binding of target primary antibodies to tissue sections (Supplementary Fig. S1; Supplementary Data are available online at
Transplantation of hESC-CECs encapsulated in hydrogel
The poly(N-isopropylacrylamide-co-acrylic acid) (p(NiPAAm-co-AAc)) copolymer was synthesized as previously reported [42]. To prepare the composite gel, a cell pellet (5 × 106 cells) was suspended in a solution of p(NiPAAm-co-AAc) mixed with or without BMP2. All reagents were cell culture grade. Briefly, p(NiPAAm-co-AAc) was embedded with the suspension of cells (5 × 106 cells) and mixed by vigorous vortexing. Five-week-old nude mice were purchased from Clea Japan. All animal experiments were approved by the Animal Care Committee of CHA University. The animals were divided into four groups for the implantation experiments. Dissociated single cells of nontransduced hEBs or hESC-CECs encapsulated in hydrogel with or without BMP2 were transplanted into the back subcutis of the mice. At 1 and 3 weeks posttreatment, mice were sacrificed (n = 4 for each time point) using an overdose injection of an anesthetic, and a region of the skin that included the transplanted site was carefully removed for subsequent biological examination.
RT-PCR analysis
RNA extractions of all samples were performed using Trizol (Invitrogen) according to the manufacturer's instructions. Total RNA (0.5 μg) was reverse-transcribed in a 20 μL reaction mixture using MMLV reverse transcriptase and random hexamers according to the manufacturer's protocol (Invitrogen). The PCRs were performed using 2 × SeeAmp™ ACP™ Master Mix II (Seegene). PCR products were confirmed in a 1.2% agarose gel visualized under UV light with ethidium bromide staining. The primer sequences and sizes of the amplified products are shown in Supplementary Table S1.
Alkaline phosphatase activity assay in vivo
To analyze the osteoblastic differentiation of hESC-CECs, intracellular alkaline phosphatase (ALP) activity was determined using p-nitrophenyl-phosphate (pNPP) hydrolysis as measured by an ALP Assay Kit (Wako Pure Chemical Industries Ltd.). The collected scaffolds were washed three times with PBS, cut down with scissors, and homogenized in lysis buffer (0.2% Igepal CA-630, 10 mM Tris–HCl, 1 mM MgCl2, pH 7.5). The lysate (2 mL) was then centrifuged at 12,000 rpm for 10 min, and the supernatant was assayed for ALP activity using pNPP as a substrate. To each well of a 96-well plate (well area = 28.26 mm2; Corning), an aliquot (2.5 μL) of supernatant was added to 25 μL of 56 mM 2-amino-2-methyl-1,3-propanediol (pH 9.8) containing pNPP with 1 mM MgCl2. The mixture was incubated at 37°C for 30 min. Next, 250 μL of 0.02 N NaOH was added to the wells to stop the reaction before the absorption was measured at 405 nm using a spectrophotometer. ALP was determined as the number of millimoles of pNP released per scaffold after 30 min of incubation.
Statistical analysis
The statistical significance of differences between experimental groups was assessed using a two-tailed Student's t-test. As shown in Fig. 2, multiple comparisons were generated using the Bonferroni adjustment to calculate the P values [42]. A P value of <0.05 was considered significant.
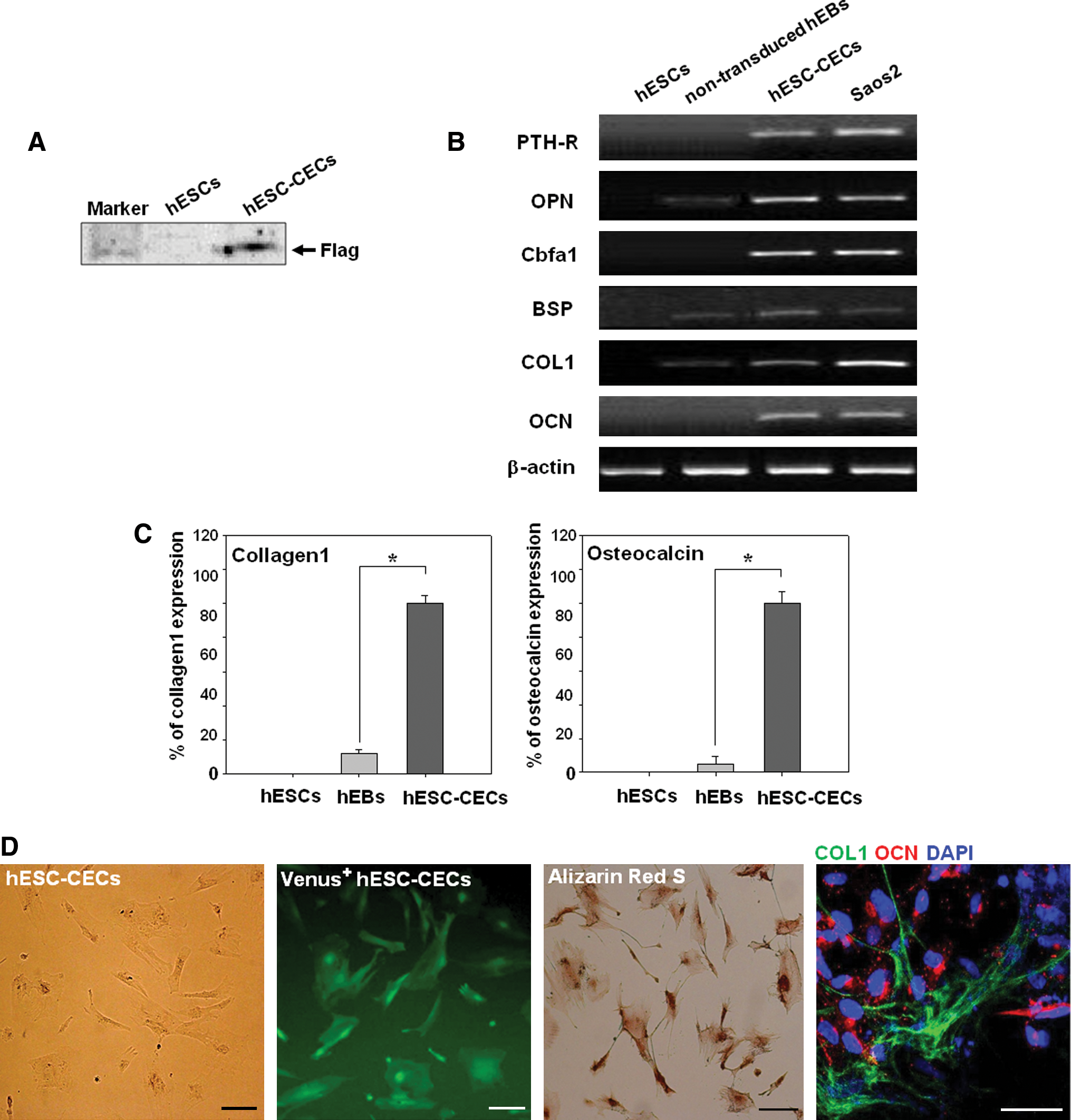
Characterization of Cbfa1-expressing cells derived from hESCs (hESC-CECs).
Results
Cbfa1 gene transduction and isolation of hESC-CECs
To obtain highly purified hESC-derived osteogenic cells with a high yield, we used genetic modification techniques and molecular tools. First, we constructed an expression vector containing the osteoblast-specific gene Cbfa1. The vector also drives the expression of Venus fluorescence gene and contains common domains that include a central polypurine tract, a central termination sequence, a promoter region, a gateway reading frame cassette that contains Cm, ccdB, attR1, and attR2 (RfA), a Venus transgene [43], and a woodchuck hepatitis virus posttranscriptional regulatory element (Fig. 1A). The viral particles expressing Cbfa1-driven Venus were produced using a lentivirus system. To genetically modify the differentiation of hESCs, we first assessed the spontaneous differentiation of hESCs under hEB suspension culture conditions. hESCs were also transduced with the Cbfa1- and Venus-expressing lentiviral particles. Viral particles were introduced twice (on days 0 and 3) to increase the transduction efficiency of hEB differentiation (Fig. 1B, virus particle treatment). Cbfa1-expressing hEBs were observed during hEB differentiation (Fig. 1B, Venus+ hEBs). Next, to isolate the Cbfa1-Venus–positive cells from differentiating hEBs, fluorescence-activated cell sorting (FACS) was performed, which revealed that the Venus-positive cell population accounted for 18.21% of the total cell population (Fig. 1B, Cbfa1-expressing cells). Finally, to investigate whether encapsulation of BMP2 with hESC-CECs in hydrogel accelerates bone tissue regeneration, hydrogel-encapsulated hEBs or hESC-CECs with or without BMP2 were transplanted into the back subcutis of mice (Fig. 1B, encapsulation).
Characterization of hESC-CECs
We first assessed the expression of transduced Cbfa1 by western blot analysis. A Flag epitope tag was fused to Cbfa1 in the transduced construct (Fig. 1A). Thus, the detection of Flag expression using an anti-Flag antibody revealed that Cbfa1 was transduced and expressed within the cells (Fig. 2A). Next, to investigate whether Cbfa1-overexpressing cells possess osteogenic cell characteristics in vitro, we examined the expression of osteoblast-related genes and proteins by RT-PCR, qRT-PCR, and immunostaining. The expression of osteoblast-specific genes such as parathyroid hormone receptor, osteopontin, Cbfa1, bone sialoprotein (BSP), COL1, and OCN was analyzed at each stage of hESC differentiation by RT-PCR (Fig. 2B). The expression patterns of osteoblast-specific genes were not detected or were weakly expressed in undifferentiated hESCs and in nontransduced hEBs, respectively. hESC-CECs in which the Cbfa1 gene was overexpressed in hEBs (Fig. 1B) strongly expressed all osteoblast-specific genes. This expression pattern was similar to that detected in the osteoblastoma cell line Saos2 (Fig. 2B). The relative expression levels of the osteoblast-specific genes COL1 and OCN were examined in undifferentiated hESCs, nontransduced hEBs and Cbfa1-overexpressing hEBs by qRT-PCR. As expected, the expression patterns of COL1 and OCN were increased by 7- and 100-fold in Cbfa1-overexpressing hEBs compared with undifferentiated hESCs or nontransduced hEBs, respectively (Fig. 2C). Based on these results, we conclude that the Cbfa1 overexpression construct functions efficiently.
Cbfa1-overexpressing cells demonstrated osteogenic-like cell characteristics at the gene level during hEB differentiation. The Cbfa1-overexpressing cells had the potential to express Venus fluorescence following Cbfa1-driven IRES activation in the vector construct (Fig. 1A). Thus, we sorted Cbfa1-overexpressing cells by FACS. The sorted cells displayed a spindle-like structure that is characteristic of osteoblast cells (Fig. 2D, hESC-CECs), and Venus fluorescence was observed in all of the sorted cells (Fig. 2D, Venus+ hESC-CECs). The hESC-CECs stained positive for COL1 at the cell junctions, whereas OCN was detected in the cytoplasm of the cells (Fig. 2D). Next, hESC-CECs were further examined for osteoblast-specific functions in vitro. Dense mineralized deposits of phosphate and calcium were detected in hESC-CECs by alizarin red S staining. All of the cells showed strong staining with alizarin red S, which indicated that hESC-CECs possessed osteoblast characteristics in vitro (Fig. 2D).
Expression pattern of osteogenic genes after cell transplantation
To investigate whether cultures isolated from differentiating hESCs possess the characteristics of osteoblasts and whether treatment with BMP2 affects osteoblastic differentiation in vivo, we transplanted hESC derivatives into the back subcutis of four groups of mice. Group I served as the control group (n = 12) and was transplanted with hydrogel constructs with hEBs. In Group II (n = 12), BMP2 and hEBs were mixed with the hydrogel constructs. Group III (n = 12) was transplanted with hydrogel constructs containing hESC-CECs, and Group IV (n = 12) was transplanted with BMP2 and hESC-CECs in hydrogel constructs. To confirm the osteoblastic differentiation of injected cells at 1 and 3 weeks posttransplantation, we first analyzed the effects of Cbfa1 overexpression and BMP2 treatment on the expression of osteoblast-specific markers (BSP, OCN, and COL1) by RT-PCR (Fig. 3A). After 1 week, the hEB-embedded constructs with or without BMP2 did not express detectable osteoblast-specific markers, but the hESC-CEC–embedded constructs with or without BMP2 displayed weak expression. By the 3-week time point, the hESC-CEC–embedded construct in the BMP2 group showed the strongest expression of osteoblast-specific genes. This result suggests that the implanted hESC-CECs rapidly promoted the formation of bone tissue and that treatment with BMP2 increased osteogenic activity.
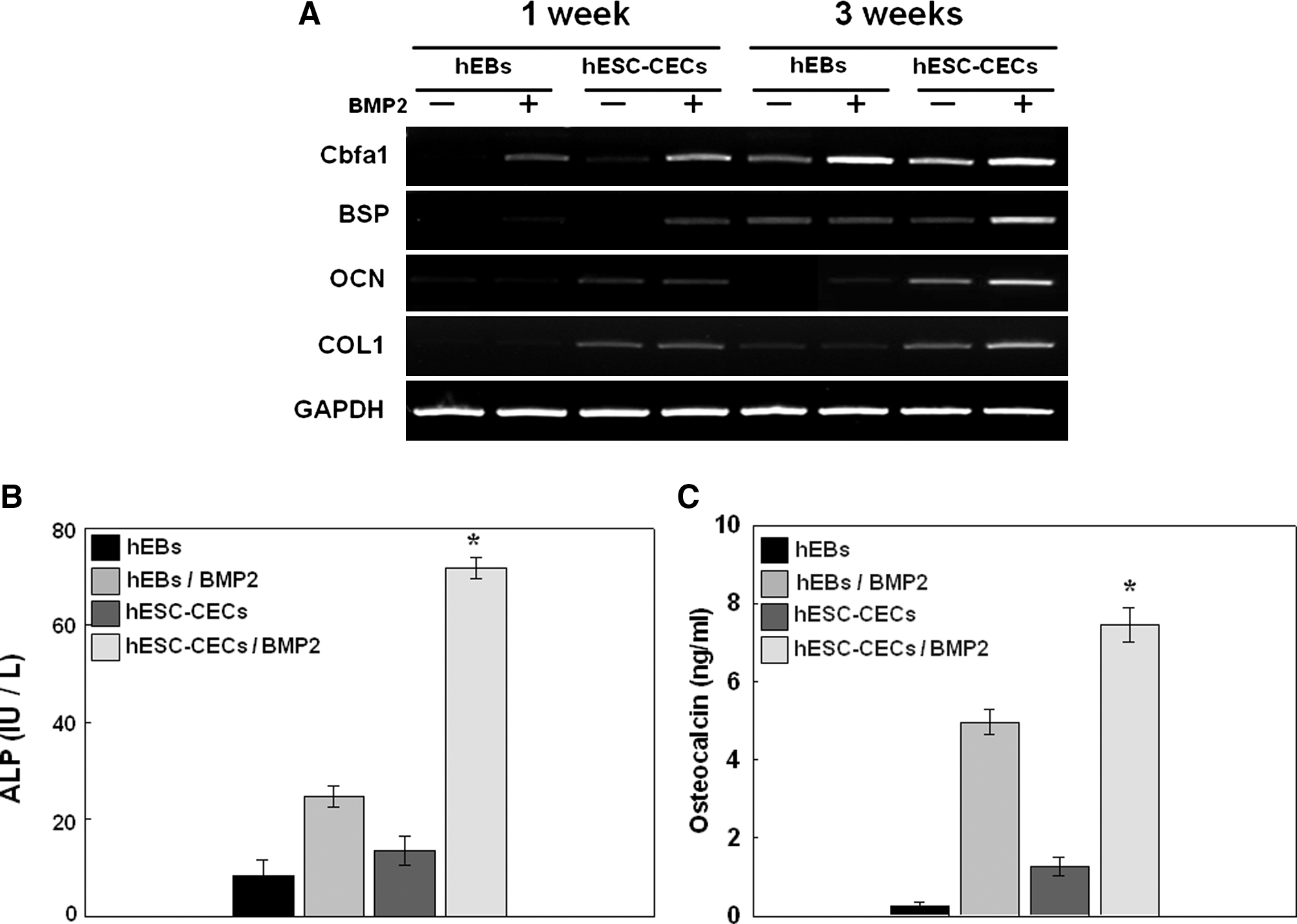
Characterization of hESC-CECs after transplantation. RT–PCR analysis of gene expression levels of Cbfa1, COL1, BSP, and OCN (with GAPDH as an internal control) at 1 and 3 weeks posttransplantation in vivo
Measurement of ALP activity in transplanted cells in vivo
To examine the functional effect of hESC-CECs on osteoblastic differentiation in vivo, we next measured the biochemical activity of ALP at 3 weeks posttransplantation. The BMP2-treated groups showed higher ALP activity compared with the nontreated groups. The ALP activity in BMP2-treated hESC-CECs was significantly higher than in the other groups, indicating that BMP2 plays a role in accelerating ALP activity in transplanted cells in vivo. ALP activity increased with time during the initial 2 weeks and plateaued thereafter, although the levels remained high compared with those in the control group (data not shown).
Analysis of mineral deposition in each group
To examine the efficacy of osteoblastic differentiation from injected cells after implantation, groups of cells embedded with or without BMP2 in the hydrogel were examined histologically by staining with hematoxylin and eosin, alizarin red S, and von Kossa (Fig. 4A-a–l). Hematoxylin and eosin staining revealed significant differences between the cell populations that were incubated in the hydrogel construct with and without BMP2. The BMP2-containing hydrogel constructs increased the proliferative potential or survival of injected cells compared with hydrogel constructs that lacked BMP2. In addition, we measured the mineral deposition in the hydrogel constructs by von Kossa and alizarin red S staining. Significantly higher calcium deposition was observed in hESC-CECs with the BMP2-containing hydrogel construct at 3 weeks posttransplantation (Fig. 4A-h, l). On the other hand, the levels of calcium deposition were not clearly detectable in hEBs in the mixed hydrogel construct group (Fig. 4A-e, i). The hEBs in the BMP2-containing hydrogel construct and those in the construct-only group showed similar patterns of weak von Kossa and alizarin red S staining, whereas the hESC-CECs embedded in the BMP2-containing hydrogel showed robust staining (Fig. 4A-f, g, j, and k).
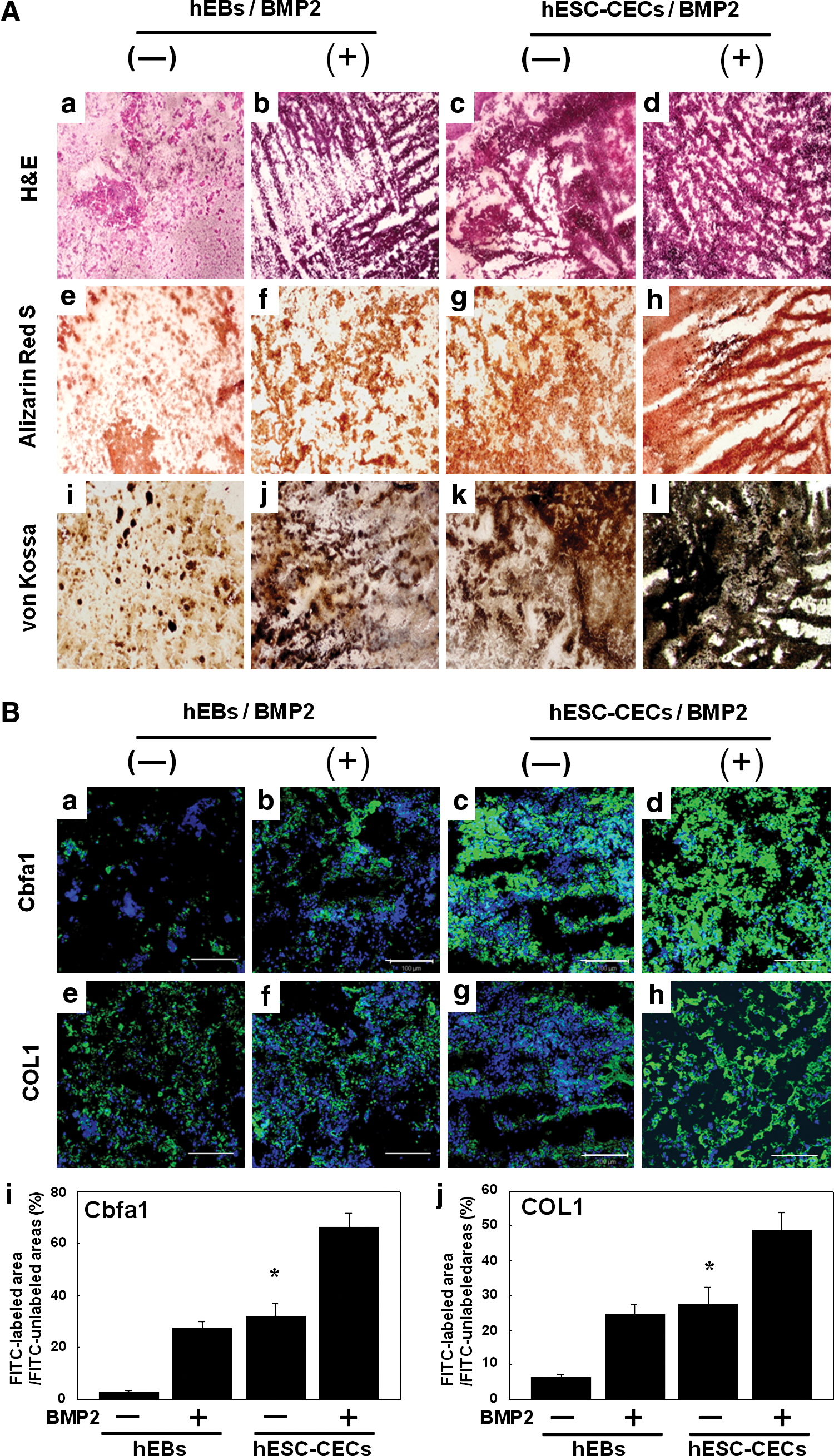
Histochemistry
Expression patterns of COL1 and Cbfa1 in vivo
To examine the osteogenesis of bone tissues formed in hydrogel constructs, we evaluated the expression patterns of osteoblast-specific markers such as COL1 and Cbfa1 by immunohistochemical staining. The hESC-CECs embedded in the BMP2-containing hydrogel showed strong staining with both COL1 and Cbfa1 antibodies (Fig. 4B-i, j), whereas hEBs embedded only in the hydrogel construct showed weak expression of both Cbfa1 and COL1 compared with all of the other groups (Fig. 4B-a, e). In addition, the hEBs in the BMP2-containing hydrogel construct and the hESC-CECs in the mixed hydrogel construct displayed a similar pattern of weak Cbfa1 and COL1 staining compared with hESC-CECs in the BMP2-containing hydrogel construct (Fig. 4B-b, c, f, and g). We found significant differences in the expression levels of COL1 and Cbfa1 between the hEB and hESC-CEC groups in the presence or absence of BMP2 (Fig. 4B-i, j).
Discussion
Various stem cell types may be used as alternative cell sources during therapeutic osteogenesis for the treatment of large bone defects that cannot be healed naturally [44]. The most obvious advantage of using hESCs instead of MSCs for osteoregeneration is that hESCs are immortal and could potentially provide an unlimited supply of differentiated osteoblast-like cells [44]. Although some reports have shown the derivation of osteoblast-like cells from hESCs in vitro [45], the capacity to direct their osteoblastic differentiation through genetic modulation has not yet been demonstrated. Here, we demonstrate the first successful regeneration of bone tissues using directly differentiated osteogenic cells derived from hESCs through lentiviral transduction of the osteogenic transcription factor Cbfa1 into hESCs. In our previous study, the EF-1a promoter was shown to drive gene expression in hESCs more efficiently than the CMV or CA promoter when a lentiviral vector system was used [29]. Thus, to enrich the transduction efficiency of the Cbfa1 gene, we initially created hESCs carrying the Cbfa1 gene under the control of the EF-1α promoter. Because Cbfa1 is a transcription factor that cannot be detected using a surface antibody, Cbfa1-positive cells cannot be purified by FACS. However, our lentiviral Cbfa1 gene transduction system was able to overcome these problems, and we obtained over 18% Venus-positive hESC-CECs by FACS (Fig. 1). As shown in Fig. 2, a homogeneous population was prepared by sorting Venus-positive cells, and these cells demonstrated high expression levels of osteoblast-related genes such as parathyroid hormone receptor, osteopontin, BSP, COL1, and OCN. Above all, mineral deposition was detected in most of the sorted cells. The results obtained for the in vitro characterization of hESC-CECs suggest that the osteoblastic differentiation of hESCs using this gene transduction system produces highly purified osteogenic cells with a high yield.
With respect to bone formation, the effects of BMP2 on stem cells have been previously reported to depend on the context in which the factor is provided [39]. We have also previously demonstrated the accumulation of an osteogenic matrix released from stem cells mixed with BMP2 in a hydrogel construct [42]. In the present study, we examined the bioactivity of BMP2 mixed with hESC-CECs in hydrogel constructs. Interestingly, the hydrogel constructs with hESC-CECs and BMP2 grew without additional osteogenic supplements. These findings suggest that BMP2 influences and supports the osteoblastic differentiation of implanted cells in a manner that promotes the expression of the osteogenic phenotype and potentially eliminates the need for external supplements. During bone regeneration, a distinct bone-specific morphology and identifiable physiological characteristics, such as calcium deposition and the appearance of COL1 and BSP expression, are normally observed [46]. The potential to produce in vivo an explantable material that mimics natural bone tissue and could be utilized to repair damaged tissues or to reconstruct bone defects is appealing [47]. Figure 4B shows the expression levels of the proteins COL1 and Cbfa1 in bone tissues from the hydrogel constructs (with and without BMP2), as evaluated by immunostaining with COL1 and Cbfa1 antibodies. COL1 was produced at higher levels by hESC-CECs cultivated in the BMP2-containing hydrogel construct than by the other cell groups. Interestingly, the hESC-CECs were still capable of undergoing osteoblastic differentiation when BMP2 was mixed into the hydrogel construct, and the expression of bone-specific proteins was still induced by the growth factor BMP2. Based on these results, hESC-CECs and BMP2 mixed in a hydrogel construct could offer a suitable environment for retention of the osteoblast phenotype while allowing the synthesis of functional bone-related materials, such as the extracellular matrix. This new system may be suitable for further research and for the development of novel biomedical therapies.
Footnotes
Acknowledgment
This research was supported by a grant (SC3110) from the Stem cell Research Center of the 21st Century Frontier Research Program funded by the Ministry of Education, Science, and Technology, Republic of Korea.
Author Disclosure Statement
The authors declare no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.