
Abstract
Murine embryonic stem cells (mESCs) are pluripotent cells that can be propagated in an undifferentiated state in continuous culture on a feeder layer or without feeders in the presence of leukemia inhibitory factor (LIF). Although there has been a great advance since their establishment, ESC culture is still complex and expensive. Therefore, finding culture conditions that maintain the self-renewal of ESCs, preventing their differentiation and promoting their proliferation, is still an area of great interest. In this work, we studied the effects of the conditioned medium from a bovine granulosa cell line (BGC-CM) on the maintenance of self-renewal and pluripotency of mESCs. We found that this medium is able to maintain mESCs' self-renewal while preserving its critical properties without LIF addition. mESCs cultured in BGC-CM expressed the stem cell markers Oct4, Sox2, Nanog, SSEA-1, Klf4, Rex1, and ECAT1. Moreover, mESCs cultured in BGC-CM gave rise to embryoid bodies and teratomas that differentiated effectively to diverse cell populations from endoderm, mesoderm, and ectoderm. Further, we found that mESCs cultured in BGC-CM have an increased proliferation rate compared with cells grown in the mESC standard culture medium supplemented with LIF. These findings may provide a powerful tool to culture mESCs for long periods of time with high proliferation rate while preserving its basic characteristics, contributing to the application of these cells to assess potential tissue engineering and cellular therapy applications.
Introduction
M
mESCs grow as colonies and can be cultured in vitro for extended periods, maintaining their capability to contribute to all cell lineages when implanted into a blastocyst. Their pluripotency is demonstrated by their ability to generate chimeras and transgenic mice and to differentiate in culture into ectodermal, endodermal, and mesodermal derivatives [3].
mESCs can be propagated in an undifferentiated state in continuous culture on a feeder layer of mitotically inactivated mouse embryonic fibroblasts (MEFs) or without feeders in the presence of leukemia inhibitory factor (LIF) or related cytokines [4]. Upon withdrawal of this factor, mESCs give rise to dimensional complex structures called embryoid bodies (EBs). These cell aggregates can be further induced to differentiate into a variety of specific cell types [5].
Although there has been a great advance since their establishment, ESC culture is still difficult and expensive. Therefore, finding conditions that maintain the stable self-renewal of ESCs, preventing their differentiation and promoting their proliferation, is still an area of great interest.
Granulosa cells (GCs) are the primary cells in the ovary that provide the physical support and microenvironment required for the developing oocyte [6]. They are a major component of the ovarian follicles, surrounding the oocytes and playing a key role in follicle development [7]. Moreover, the GCs are an abundant source of numerous factors such as transforming growth factor β (TGF-β), insulin-like growth factors (IGF), IGF-binding proteins, activins, and inhibins [6,8].
Research performed by one of us (L.B.) led to the obtention of a bovine GC line (BGC-1) by spontaneous immortalization of primary cultures, which was originally selected for its maintenance of hormonally regulated fibronectin (FN) gene expression [9]. BGC-1 cells show some characteristics of primary cultures, such as the hormonal regulation of FN biosynthesis, similar responsiveness to growth factors, and a qualitatively similar protein secretion pattern [10,11]. In addition, conditioned medium from this cell line (BGC-CM) is mitogenic for both BGC-1 cells and primary cultures of BGCs [10]. The myriad of factors secreted by GCs and the mitogenic properties that BGC-CM showed led us to speculate that this cell line may provide a CM with valuable properties.
In the present work, we studied the effect of the CM from the BGC line, described above, on the maintenance of self-renewal and pluripotency of mESCs. We found that BGC-CM is able to maintain mESCs' self-renewal while preserving its critical properties without LIF addition. We have also found that mESCs cultured in this medium have an increased proliferation rate compared with cells grown in the mESC standard culture medium supplemented with LIF. These findings could provide a powerful tool to culture mESCs while preserving their basic properties and increasing their proliferation rate, both highly desirable features in an ESC culture medium.
Materials and Methods
Cell culture
Ainv15 and R1 mESC lines were obtained from ATCC. The mESCs were first expanded on an MEF feeder layer previously irradiated. Then, subculturing was carried out according to specifications. Briefly, subconfluent cultures were trypsinized and replated (1:3–1:6 split) every 48–72 h onto irradiated MEFs or 0.1% bovine gelatin-coated tissue culture plates. Cells were propagated either in mESC standard medium consisting of knockout Dulbecco's minimal essential medium (high-glucose DMEM) (Invitrogen/Gibco 10829-018) supplemented with 15% ES-qualified fetal bovine serum (FBS) (Invitrogen/Gibco 16141-061), 100 μM MEM nonessential amino acids (Gibco), 0.5 mM beta-mercaptoethanol, 2 mM
Cell differentiation
mESCs were differentiated using the hanging drop method. Briefly, cells were trypsinized, centrifuged, and suspended in differentiation medium (standard medium containing 20% FSB, without LIF) to a concentration of 25,000 cells/mL. Afterward, 25-μL drops were placed on the lid of a bacterial plate. After 48 h, EBs formed in the bottom of the drops were collected and placed in new bacterial plates with fresh differentiation medium, allowing them to grow for 3 more days. On the fifth day, EBs were collected and replated onto 0.1% gelatin-coated dishes. From the sixth day onward, adherent EBs were used as described.
Conditioned media experiments
BGC-1 or HEK293T cells were grown in high-glucose DMEM supplemented with 10% FBS (Gibco), 1 mM sodium pyruvate (Sigma), and antibiotics to a 90% confluence. Then, the culture medium was replaced by fresh standard ES medium without FBS and beta-mercaptoethanol and incubated for 24 h. After incubation, CM was removed, centrifuged at 2,000 g for 10 min, and supplemented with 15% FBS and 0.5 mM beta-mercaptoethanol before use.
Reverse transcription–polymerase chain reaction
ESCs were grown in BGC-CM, standard medium plus LIF, standard medium without LIF, or HEK293T-CM for at least three passages or 7 days on 0.1% bovine gelatin-coated tissue culture plates. Total cellular RNA was isolated from subconfluent cultures according to Chomczinsky and Sacchi [12]. The yield and purity of RNA samples were assessed by the absorbance at 260 and 260/280 nm ratio, respectively. One microgram of total RNA was retrotranscribed using MMLV reverse transcriptase (Promega) and 30 ng/μL Random Primers (Invitrogen). Conventional polymerase chain reaction (PCR) amplification of DNA was carried out using Taq DNA Polymerase (Invitrogen) and 12.5 pM specific oligonucleotides (Table 1) in a Peltier Thermal Cycler DNA engine (BioRad). Analysis of the PCR products was performed by electrophoresis in a 2% agarose gel stained with ethidium bromide.
Immunofluorescence
ESCs were seeded on Permanox chamber slides (Nunc) and grown in the indicated medium. The cells grown in each condition for at least three passages or 7 days on 0.1% bovine gelatin-coated tissue culture plates were fixed in 4% paraformaldehyde for 20 min. Fixed cells were permeabilized in 0.2% Triton X-100 for 30 min, blocked with 1% phosphate-buffered saline (PBS)–0.01% Tween normal goat serum, and incubated overnight with polyclonal primary antibodies listed in Table 2. Primary antibodies were detected with the secondary antibodies mentioned in Table 2. Nuclei were stained with DAPI according to the manufacturer's instructions (Santa Cruz Biotechnology).
Teratoma formation and histological analysis
The ESCs that have been cultured in BGC-CM for at least three passages or 7 days on 0.1% bovine gelatin-coated tissue culture plates were trypsinized, centrifuged, and suspended at 1 × 107 cells/mL in DMEM in the absence of FBS. Cells were kept at 37°C until the injection. Then, 100 μL of the cell suspension (1 × 106 cells) were subcutaneously injected into the dorsal flank of 6–8-week-old male nude (nu/nu) mice. Four weeks after the injection, animals were anesthetized in a CO2 atmosphere chamber and sacrificed by cervical dislocation. Tumors were surgically dissected from the mice, fixed in PBS containing 4% formaldehyde, and embedded in paraffin. Sections (4 μm) were stained with hematoxylin and eosin or immunostained. For immunohistochemistry, sections were blocked with 1% goat serum in PBS, washed, and incubated with the indicated primary antibodies (Table 2) for 1 h at room temperature. After rinsing in PBS, slides were incubated for 30 min with horseradish peroxidase-conjugated secondary antibodies, washed, and revealed (ABC Kit; Vector; Cat. No. PK-6200). Then, the samples were stained with hematoxylin. Mouse procedures were conducted under local ethical guidelines.
Cell proliferation assay
ESCs were grown in the standard medium with LIF. Then, cells were trypsinized and replated in a flat-bottomed 96-well plate. For each experiment, 3,000 cells per well were seeded and allowed to grow in either BGC-CM or standard medium plus LIF prepared without phenol red. Cell proliferation was quantified by the sodium 2,3,-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)-carbonyl]-2H-tetrazolium inner salt (XTT) method (Sigma) according to manufacturer's instructions. Briefly, a volume of XTT solution equal to 20% of culture medium was added 3 h prior to measurement. Formation of colored product, the orange formazan derivative, was quantified by collecting the culture medium, reading its absorbance at 450 nm, and subtracting background contribution from the reactive sodium salt of XTT at 690 nm in a multiwell plate reader (Microplate Reader, BioRad model 680). An initial measurement was taken 3 h after seeding cells in the plate. Afterward, absorbance at 450 and 690 nm was measured once a day during at least 5 days. To assess cell proliferation rate, corrected absorbance vs. time was graphed and slopes for each culture medium were then compared.
Results
To determine if BGC-CM is capable to maintain mESCs' self-renewal, we cultured Ainv15 mESC line in the standard mESC medium that was previously conditioned by the BGC line, without LIF addition. We compared these mESCs with parallel cultures grown in the mESC standard medium supplemented with LIF or the standard medium without LIF. We evaluated the basic properties of stem cells cultured on gelatin-coated plates, such as colony morphology, expression of undifferentiated state-specific gene markers, and pluripotency.
A distinctive characteristic of mESCs is their defined colony morphology on culture. Changes in this morphology are highly notorious and clearly indicate whether the cells remain in an undifferentiated state or have undergone any differentiation process [3]. To have a broad view about the effect of BGC-CM on mESCs' self-renewal, we first examined the morphology of mESC colonies grown in the different experimental conditions for at least three passages by bright-field microscopy. As shown in Fig. 1, mESCs cultured in both the standard medium plus LIF and the BGC-CM presented a similar typical colony morphology, high refringence, and a low cytoplasm/nucleus ratio. On the other hand, both the negative control cultured in the standard medium without LIF and the mESCs cultured in a medium conditioned by a different cell line showed a clearly different aspect.
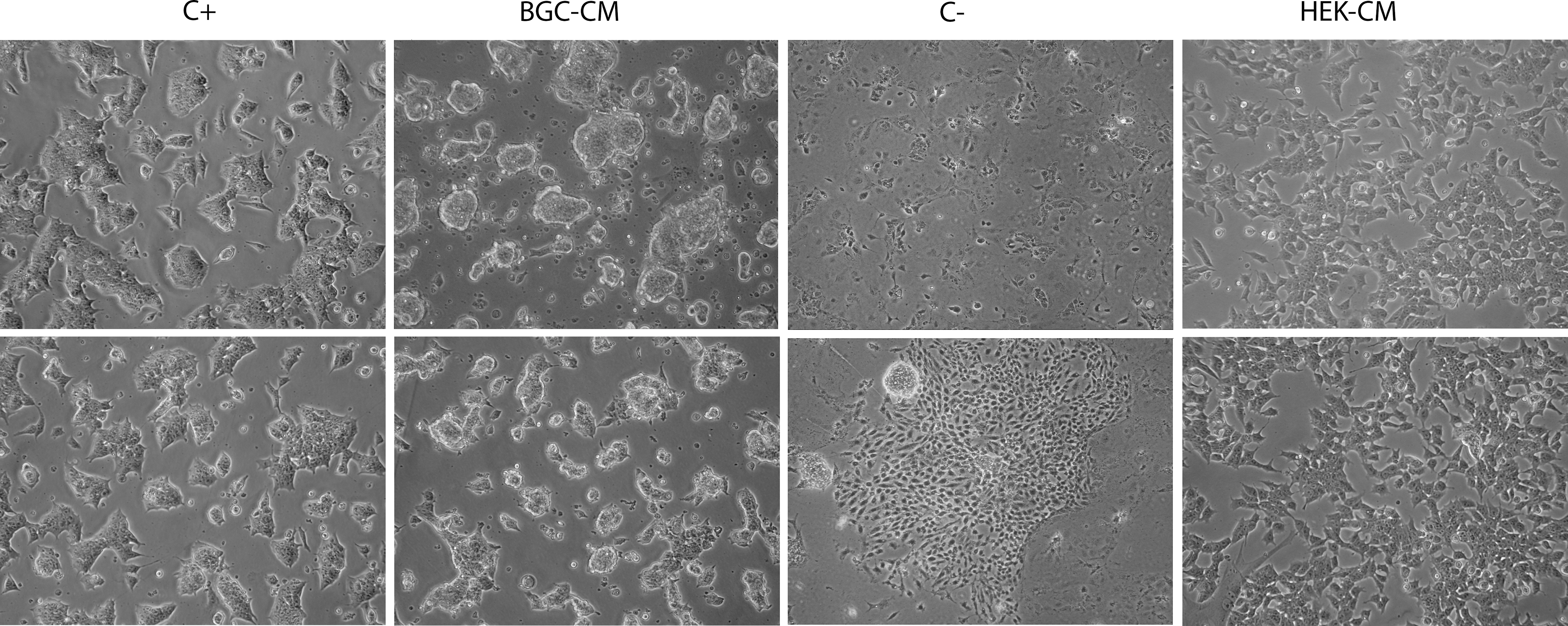
mESCs cultured in BGC-CM show typical undifferentiated colony morphology. Ainv15 mESCs were cultured in the indicated medium for at least three passages or 7 days. Representative phase-contrast images of colonies cultured in BGC-CM, standard medium plus LIF (C+), standard medium without LIF (C−), or HEK293T-conditioned medium (HEK-CM). mESCs, murine embryonic stem cells; BGC, bovine granulosa cell; CM, conditioned medium; LIF, leukemia inhibitory factor.
We next studied the expression of undifferentiated state-specific gene markers by immunofluorescence and RT-PCR. We first analyzed the expression of the stem cell markers Oct4, Sox2, Nanog, SSEA-1, and Klf4. Then, we also studied other genes expressed in mESCs, such as Rex1 and ECAT1 [13 –15].
As shown in Fig. 2, we detected the expression of all the mESC markers evaluated in mESC cultures maintained in BGC-CM. Moreover, we were able to culture mESCs up to 20 passages in this medium, and we did not detect changes in the pattern of gene expression or the colony morphology (data not shown). As expected, in the negative control, the expression levels of the stem cell markers evaluated were clearly lower than in the other experimental conditions (Fig. 2A and data not shown).
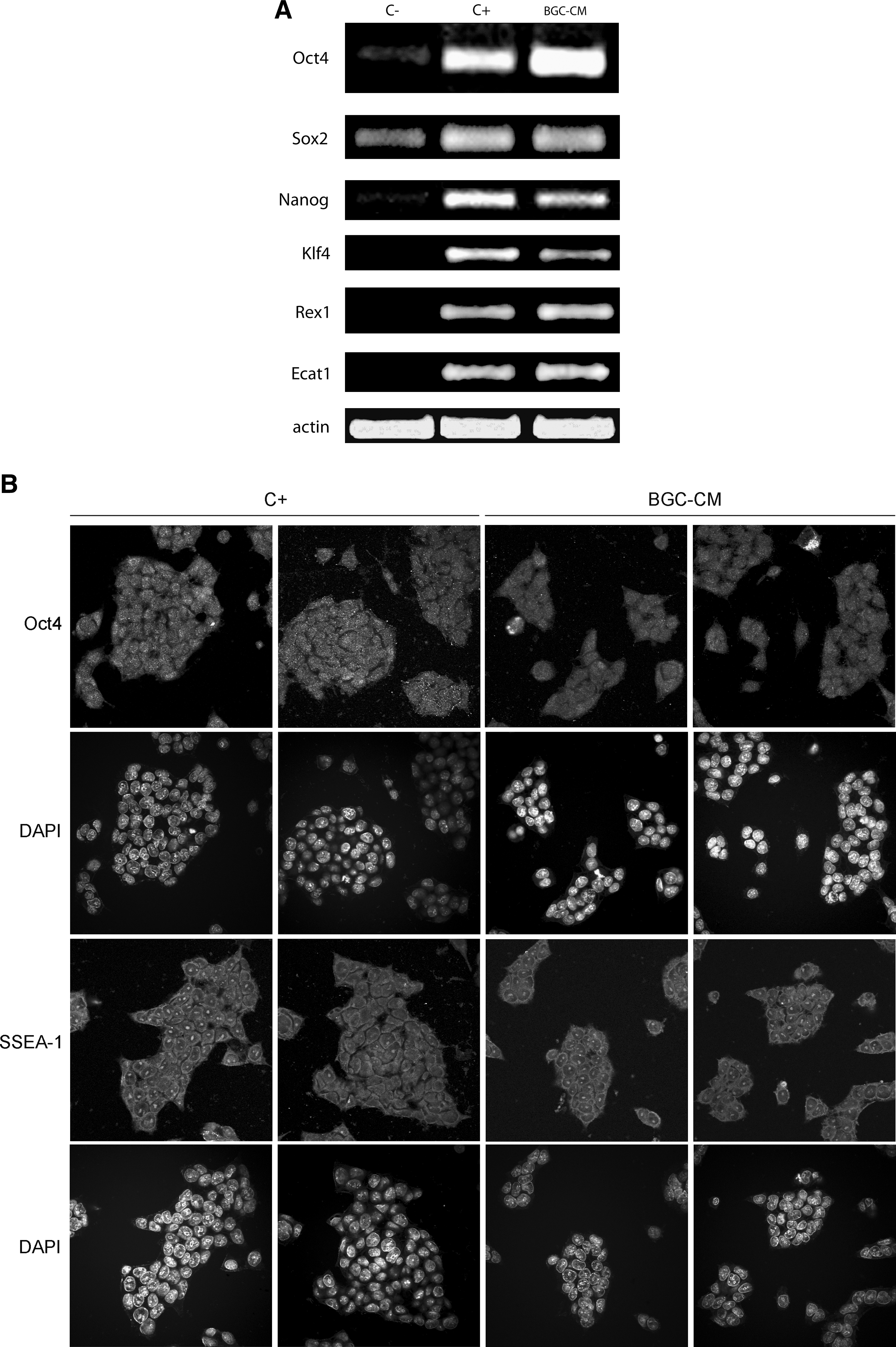
mESCs cultured in BGC-conditioned medium express undifferentiated state marker genes. mESCs were cultured in BGC-CM, standard medium plus LIF (C+), or standard medium without LIF (C−) as indicated, for at least three passages or 7 days.
Further, we have also studied BGC-CM effect on different mESC lines, such as CGR8, R1, and NKx2.5eGFP ESCs. In all cases, we were able to propagate them in BGC-CM for several passages while preserving their morphology and the expression of stem cell marker genes (data not shown).
Having established that mESCs cultured in BGC-CM preserve their undifferentiated state, we wondered whether the cells that have been cultured in this medium remain pluripotent, maintaining their capability to originate in vitro, cells from endoderm, mesoderm, and ectoderm. Following the previously described hanging drop protocol [16], we attempted to differentiate them. Both mESCs maintained in BGC-CM or standard proliferation medium gave rise to EBs (Fig. 3A). When plated on gelatin-coated Petri dishes, these EBs differentiated effectively to diverse cell populations (Fig. 3B). By immunofluorescence or RT-PCR, we analyzed the expression of alpha-fetoprotein, as an endoderm gene marker; Brachyury or alpha smooth muscle actin, as mesoderm gene markers; and βIII tubulin, as an ectoderm marker [13,14]. We found that EBs derived from ESCs cultured in BGC-CM expressed gene markers from cell types derived from the three embryonic layers (Fig. 3C, D).
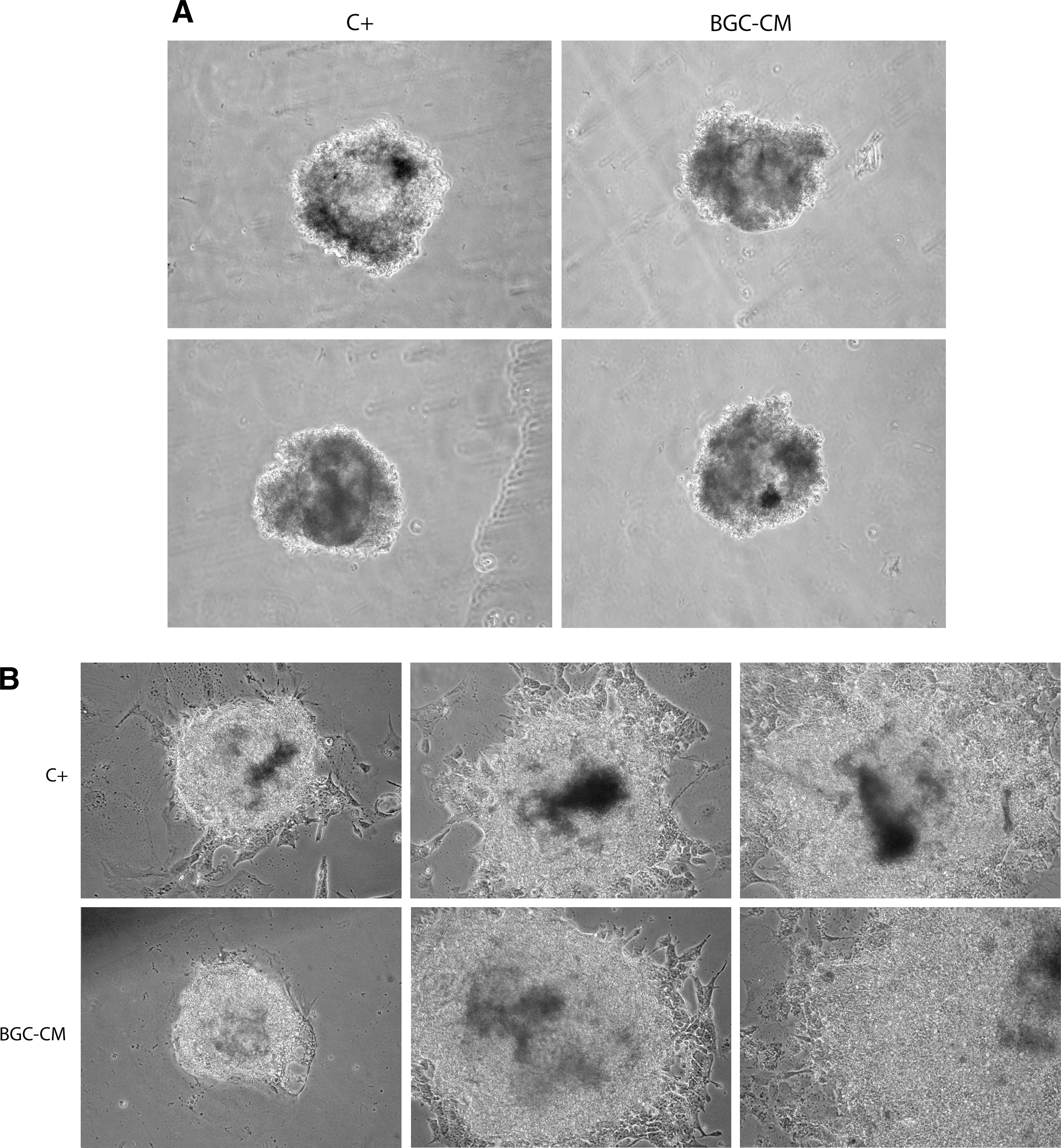
mESCs propagated in BGC-CM remain pluripotent. Ainv15 mESCs were cultured in BGC-CM or standard medium plus LIF (C+) for at least three passages or 7 days. Then, the cells propagated in the indicated medium were subjected to the hanging drop protocol and gave rise to EBs. Representative phase-contrast images of EBs in suspension

Up to this point, we have found out that mESCs maintained in BGC-CM preserve their basic properties, so we sought to evaluate whether these cells were able to differentiate to a cell type with possible therapeutic applications. The regeneration of cardiac tissue being an important target of stem cell-based therapy, we evaluated the expression of cardiomyogenic mesoderm gene markers Nkx2.5 and Miocardin [17,18]. We detected expression of these markers in Ainv15 (Fig. 2D) and in other mESC lines (data not shown), suggesting that mESCs cultured in BGC-CM preserved their ability to differentiate to cardiomyocytes.
We next examined the pluripotency of mESCs cultured in BGC-CM by teratoma formation. As shown in Fig. 4, mESCs that have been propagated in BGC-CM for at least three passages or 7 days remain pluripotent as they were able to differentiate in vivo into tissues from all the three germ layers. We confirmed differentiation into endoderm, mesoderm, and ectoderm by immunostaining with antibodies against GATA4, alpha smooth muscle actin, and βIII tubulin, respectively (Fig. 4H–J). As a whole, these data demonstrate that mESCs cultured in BGC-CM preserve its critical properties: self-renewal and pluripotency.

mESCs propagated in BGC-CM gave rise to teratomas. Four-micrometer sections from teratoma tissue were stained with hematoxylin and eosin or immunostained.
As mentioned earlier, one of the properties of BGC-CM is its mitogenic activity on both GC primary cultures and BGC-1 cell line. Based on this evidence and on the fact that the colonies of mESCs cultured in BGC-CM looked bigger than those cultured in standard conditions, we considered important to study whether this CM had a mitogenic effect on mESC lines. To test our hypothesis, we measured culture proliferation rates by XTT assay for 5 days. As shown in Fig. 5, the proliferation rate of mESCs cultured in BGC-CM was clearly higher than the rate of the cells cultured in standard conditions. This feature of BGC-CM together with its ability to preserve mESCs' basic properties makes this medium a suitable and inexpensive alternative to propagate mESCs in culture for long periods of time.
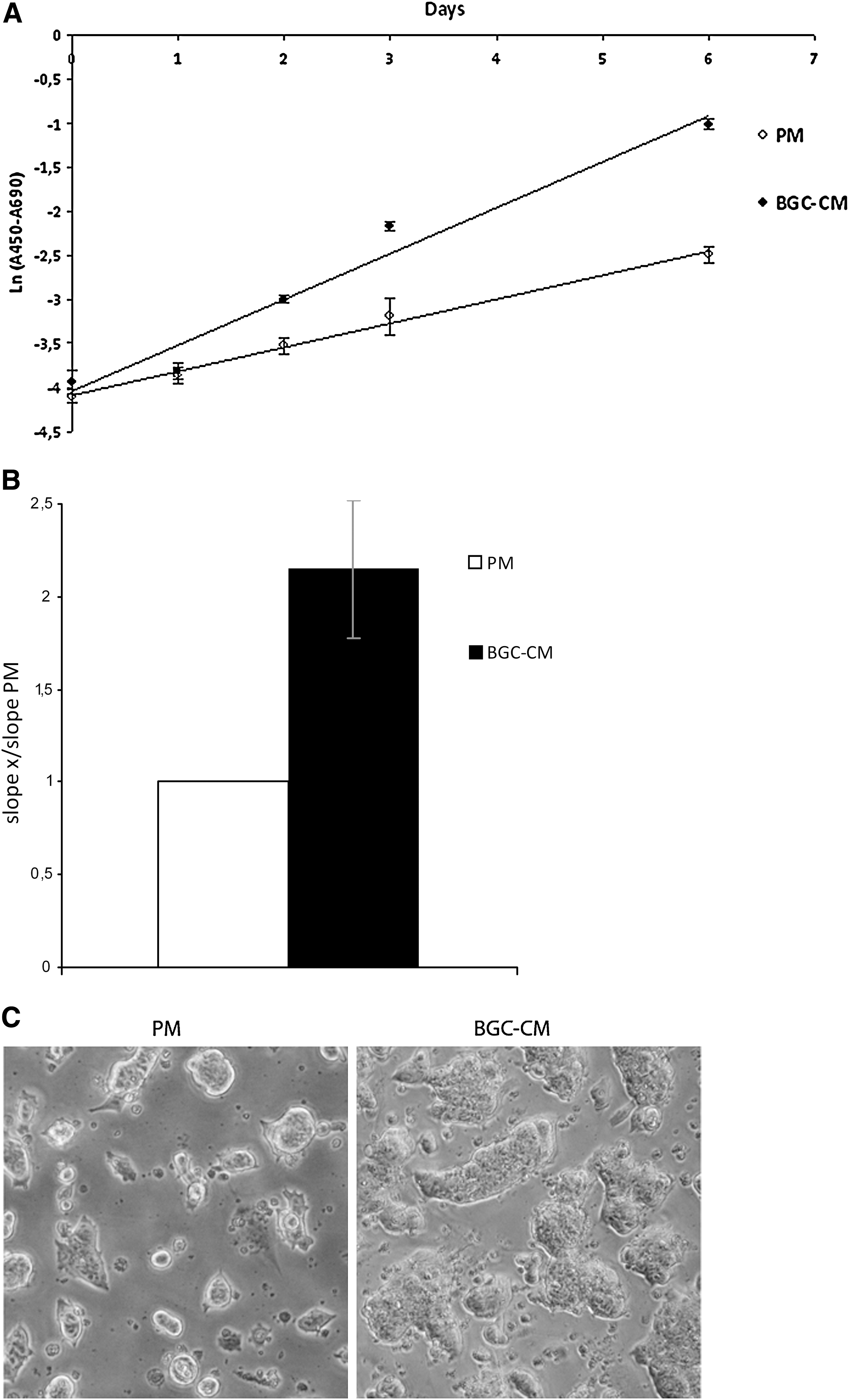
BGC-CM increases the proliferation rate of mESCs. Ainv15 mESCs were cultured in BGC-CM or the standard medium plus LIF (PM) for at least 5 days. The proliferation was evaluated once a day in the indicated medium by XTT assay.
Discussion
In this work, we have explored the possibility of culturing mESCs in a novel CM. mESCs propagated in this context maintained its critical properties and showed an increased proliferation rate.
The medium was conditioned by a BGC line previously established by one of us (L.B.) [9] and selected by its mitogenic properties on the same cell line and on granulosa primary cultures [10].
GCs express a myriad of secreted factors that could be involved in both the maintenance of mESCs' essential properties and the increase of mESC proliferation rate, such as fibroblast growth factors (FGFs), IGF, TGF-β family members, for example, Activin A [8,10,19], and LIF [20]. Moreover, Activin A [21] and LIF expression has been detected (unpublished results) in this BGC line. Originally, the mESC requirement of a feeder layer was substituted with the addition of LIF to the culture medium [22,23] and fetal serum was replaced by a TGF-β family member, the bone morphogenetic protein 4 [24]. In the last few years, multiple defined conditions that enable to culture of ESCs free of feeder layers and serum had been established. These conditions include combinations of inhibitors of pathways involved in differentiation, such as the FGF/Mek/Erk, and of glycogen synthase kinase 3 [25,26]. Although the precise components of BGC-CM have not been addressed in this work, the expression of LIF in GCs led us to speculate that this cytokine could be the main factor responsible for the maintenance of mESC features. Taking into account the multiple factors that are present in the CM, we cannot be sure that LIF is the sole factor responsible for this property. Therefore, we believe that the balance of the multiple signals in it preserves ESCs' self-renewal and pluripotency.
On the other hand, several candidates could be responsible for the increased proliferation rate of mESCs cultured in this medium. Although Activin A was reported to be a negative regulator of proliferation in some cell types, such as pancreatic acinar cells [27] and renal tubular cells [28], this TGF-β family member has mitogenic effects on multiple cell types [29], such as placenta [30], granulosa [21], and mESCs [31,32]. In contrast, although LIF induces proliferation in diverse cell types [33 –35], it has been reported that it is not related to proliferation in mESCs [36,37]. Conversely, it was reported that an isoform of FN present in the mentioned CM could be responsible for its mitogenic effect observed on GCs [11]. Moreover, it has been suggested that FN expression induced in mESCs by high glucose concentrations was responsible for an augmented proliferation of these cells [38]. Based on these evidences, we are studying whether Activin A and/or FN expressed by BGC line would be responsible for the mitogenic effect on mESCs. Moreover, we are evaluating whether there is an induction of FN expression on mESCs cultured in BGC-CM that could also be involved in this effect. Currently, we are analyzing the signaling pathways that are activated in mESCs cultured in the different conditions, followed by blocking the different candidate pathways to unravel which of them and how are they involved. Last but not least, as it is well known that bFGF is involved in human ESC pluripotency maintenance [39], and BGC line expresses this factor (unpublished results), we are evaluating whether BGC-CM preserves human ESCs' self-renewal and pluripotency.
Hereby, we propose that BGC-CM offers an inexpensive way for culturing these cells for long periods of time with a high proliferation rate, while preserving its basic characteristics. Moreover, we think that this medium would be a powerful research tool, because it provides a novel culture context to study the involved molecular mechanisms in self-renewal and pluripotency maintenance. We are confident that, in the near future, novel factors identified from this kind of culture contexts will contribute to produce improved therapy-suitable defined ESC culture medium.
Footnotes
Acknowledgments
The authors express thanks to Dr. Michel Puceat, Dr. Sean Wu, and Dr. Niels Geijsen for providing some of the mESC lines utilized in this work, to Marcelo Schultz for teratoma processing, and to Francisco Guaimas for his help with confocal microscopy. This work was supported by grants (to A.S.G.) from the University of Buenos Aires (X849), Agencia Nacional de Promoción Científica y Tecnológica (ANPCyT) (PID 115-PAE 37075), and Biosidus S.A. N.L. is supported by a fellowship grant from University of Buenos Aires; C.L., C.S., and J.B. are fellows from CONICET.
Author Disclosure Statement
No competing financial interests exist.