
Abstract
Despite its prognostic value in in vitro fertilization, early embryo morphology is not reported on in the derivation of human embryonic stem cell (hESC) lines. Standard hESC derivation does rely on blastocyst development and its efficiency is highly correlated to inner cell mass (ICM) quality. Poor-quality embryos (PQEs) donated for hESC derivation may have a range of cleavage-stage abnormalities that are known to compromise further development. This study was implemented to determine whether specific PQEs traits influence the efficiency of good-quality ICMs to derive new hESC lines. We found that although the types of PQEs investigated were all able to make blastocysts with good-quality ICMs, the ICMs were unequal in their ability to derive hESCs. Good-quality ICMs from embryos with multiple poor-quality traits were unable to generate hESC lines, in contrast to good-quality ICMs from embryos with a single poor-quality trait. In addition, our data suggest a direct correlation between the number of ICM cells present in the blastocyst and its capacity to derive new hESC lines. This study is the first to demonstrate that ICM quality alone is an incomplete indicator of hESC derivation and that application of in vitro fertilization–based early embryo scoring can help predict hESC derivation efficiency. Experiments aiming to quantify, improve upon, or compare hESC derivation efficiency should thus take into consideration early embryo morphology scoring for the comparison of groups with equal developmental competence.
Introduction
D
As with other embryo sources, hESC derivation using PQEs is most efficient when they develop to the blastocyst stage and when good-quality inner cell masses (ICMs) are used [7,9]. Although describing blastocyst quality is common in hESC derivation reporting, information on early embryo development is rare [19]. In assisted reproduction technology (ART), early embryo morphology is the best predictor of developmental competence and individual traits are not equal in their predictive value of implantation [20 –23]. Although reaching the blastocyst stage is a developmental milestone in embryo development, inclusion of early embryo morphology still provides the best indicator of further viability when selecting blastocysts for clinical use [24]. Incorporation of early embryo morphological criteria with blastocyst quality could provide similar insight into successful hESC derivation.
The classical embryological indicators of decreased implantation potential used in ART laboratories include a high degree of fragmentation [25,26], multinucleated (MN) blastomeres [27 –30], and/or delayed development [31,32]. In addition to these inferior morphological traits, human zygotes that do not contain 2 pronuclei (PN) at fertilization check are mostly discarded from fertility treatment and form another type of PQE for hESC derivation [33]. As embryo selection criteria evolve, additional poor-quality parameters such as oocyte dysmorphisms, nucleoli misalignment in the PN, and blastomere asymmetry may also be included in the designation of PQEs [34 –36]. As there are no strict guidelines for designating an embryo as poor quality, the embryos provided to hESC derivation laboratories may have varying degrees of impairment within the PQE pool and between source IVF laboratories.
There have been at least 16 reports describing the feasibility of using PQEs as a source for hESC derivation, with efficiency ranging from 4.1% to 25% [1 –16]. The variations in hESC derivation efficiency between laboratories are commonly attributed to differences in derivation methodology and experience levels. Inconsistent reporting of blastocyst use and quality make comparing efficiencies between laboratories difficult [19]. Chen et al. [37], however, reported a 13%–50% range in derivation efficiency between identical experiments using good-quality embryos. These differences suggest an underlying genetic or embryological influence that may reflect the diversity of the embryos provided for research.
This study was implemented to determine if specific poor embryo traits influence the derivation efficiency of new hESC lines. Concomitantly, this study also provides a detailed assessment of our PQE pool, their competency to develop to the blastocyst stage, and their resulting ICM quality. Using only good-quality ICMs under identical conditions, a baseline efficiency of hESC derivation for embryos with high fragmentation, MN blastomeres, delayed development, abnormal fertilization, or a combination of these traits was established. We found that although our PQE types were all able to make good-quality ICMs, these ICMs were not equal in their ability to generate new hESC lines. Surprisingly, good-quality ICMs were only able to derive new hESCs if they originated from single-trait PQEs, but not from multiple-trait PQEs. Moreover, the percentage of novel hESC lines derived is similar to the percentage of ICMs with high cell number. We conclude that the type of PQE and their ability to make high-cell-number ICMs are key determinants for the efficiency of hESC derivation.
Materials and Methods
Ethical aspects
Institutional Review Board approval was obtained from the Ethical Committee, Ghent University Hospital and the Belgian Federal Ethical Committee on Embryo Research. All patients donating embryos signed informed research consents prior to their IVF cycle.
Embryo source
Following embryo transfer, embryos that did not meet the IVF laboratory's cryopreservation criteria due to high fragmentation (≥25%) on day 2 or 3 of development (day of oocyte retrieval being day 0), MN blastomeres on day 2, delayed development (<5 blastomeres on day 3 or a ≤1 blastomere increase from day 2 to 3), and/or abnormal fertilization (0, 1, or ≥3PN) on day 1 were available for use in this study. In addition, embryos must have contained at least 4 blastomeres on the afternoon of day 3 and could not have had >50% fragmentation for inclusion into the study.
Embryo culture
Fresh, day 3 PQEs cultured from the zygote stage in Cook Cleavage medium (Cook Ireland Ltd., Limerick, Ireland;
hESC derivation
Only day 6 blastocysts with grade A and B ICMs were used for derivation purposes and whole blastocysts were plated for all derivation attempts. The zona pellucida was removed by short exposure to warmed acidic tyrode's solution (Sigma, Bornem, Belgium;
All initial culture dishes were observed daily and hESC media were refreshed on alternating days. Any cellular organization of presumptive ICM organization/growth was mechanically dissected and moved to fresh MEFs at 4–7 days after initial culture. Transplanted clumps were observed daily for hESC-like outgrowth and expanding hESC colonies were mechanically passaged every 5 days into multiple organ culture dishes until they could be transferred safely to a tissue culture flask (Falcon T25, VWR). At this time, enzymatic passaging and expansion with 1 mg/mL collagenase (type IV; Invitrogen) was carried out every 5 days with daily media refreshment. As an internal quality control of culture conditions, a previously established hESC line was cultured in parallel with this experiment.
Karyotyping of hESC lines
Nearly confluent culture flasks of hESCs at early passages were induced to metaphase by colcemide (1:100 in salt solution; Karyomax; Invitrogen), harvested with trypsin–EDTA (Invitrogen), treated with a hypotonic solution of HCL (Sigma), and fixed in a 1:3 methanol/acetic acid solution (Sigma). The metaphase spreads were then prepared and analyzed as described previously using the G-banding technique [38].
Immunofluorescence of hESC lines
Undifferentiated hESC colonies grown on MEF-coated cover slips were fixed with 4% paraformaldehyde (Sigma) for 30 min at room temperature, washed repeatedly with phosphate-buffered saline (PBS; Sigma), and incubated with blocking solution [4% normal goat serum (Invitrogen) in PBS] for 1 h at room temperature. hESCs were then exposed to primary antibody (SSEA-1, SSEA-4, TRA-1-60, and TRA-1-81; ES-cell kit SCR002; Chemicon, Millipore, Brussels, Belgium;
Alkaline phosphatase assay
After 5 days of culture, undifferentiated hESCs were fixed with 4% paraformaldehyde for 2 min at room temperature. Alkaline phosphatase assay was performed per manufacturer's instructions (Alkaline Phosphatase Detection Kit; Chemicon, Millipore).
Embryoid body formation and analysis
Embryoid body (EB) formation was carried out as described previously [39]. Briefly, nearly confluent hESC colonies were detached from their supporting MEF layer with 1 mg/mL collagenase type IV (Invitrogen) in large clumps and transferred to ultra-low-attachment 6-well plates (Corning B.V. Life Sciences, Amsterdam, The Netherlands;
Differential staining of blastocysts
The differential staining method used was adapted from Thouas et al. [40]. Briefly, zona-intact blastocysts were placed in 500 μL of HEPES-buffered Human Tubal Fluid (Lonza, Verviers, Belgium;
Statistical analysis
Percentages were compared by contingency table analysis followed by chi-square or Fisher exact tests where appropriate. P values of ≤0.05 were considered to be significant.
Results
The number of poor-quality traits influences blastocyst development and ICM quality
There were 1,221 PQEs included in the hESC derivation portion of this study and 31.2% of these developed to the blastocyst stage. Figure 1 illustrates the type, number, and blastocyst development of embryos comprising our PQE pool. Because abnormally fertilized embryos were not scored for day 2 MN, there is no overlap of these 2 traits in the Venn diagram. Embryos with a single poor-quality trait (high fragmentation, MN blastomeres, delayed development, or abnormal fertilization) comprised 65.3% of the PQE pool and had significantly better blastocyst development when compared with embryos with overlapping poor-quality traits (34.7% vs. 24.8%; P < 0.001, chi square). Of the embryos with single poor-quality traits, those with high fragmentation represented a significantly higher proportion of the PQE pool (38.8% vs. 16.2%, 23.2%, 21.8%; P < 0.001, chi square, for MN, delayed development, and abnormal fertilization, respectively). There were no differences in blastocyst development between the PQE groups having different single poor-quality traits.

The type, number, and blastocyst development (%) of poor-quality embryos available for hESC research in our center.
Overall, good-quality ICMs were formed in 40.3% of blastocysts produced or 12.6% of embryos cultured. The ICM quality for blastocysts generated by each poor quality trait is represented in Fig. 2. Only 25.7% of the blastocysts originating from embryos with multiple poor-quality traits had good-quality ICMs, which was significantly lower than those originating from highly fragmented, MN, delayed, and abnormally fertilized embryos (25.7% vs. 43.8%, 50.0%, 44.4%, and 49.0%, respectively; P < 0.05, chi square).
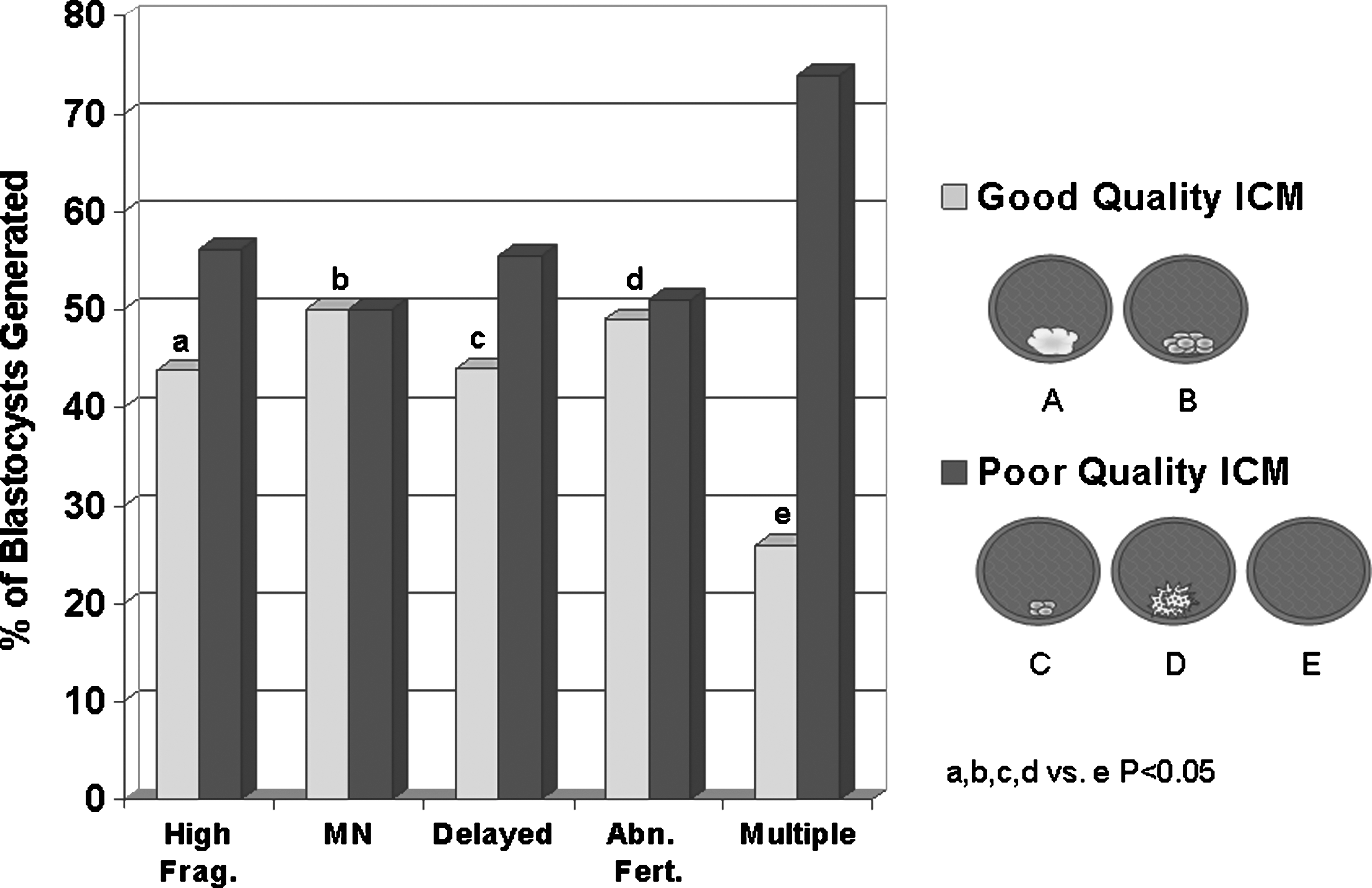
The ICM quality of blastocysts produced from embryos with different poor-quality traits. ICM, inner cell mass.
The number of poor-quality traits in the PQEs is crucial for hESC derivation
The efficiency of PQEs to generate new hESC lines using blastocysts with good-quality ICMs is presented in Table 1. Notably, embryos with multiple poor-quality traits were not able to generate new hESC lines, whereas each single-trait group was able to establish hESCs with a combined efficiency of 14.9% per attached blastocyst or 2.0% per embryo cultured. The inclusion of embryos with multiple poor-quality traits in the PQE pool lowers the overall derivation efficiency to 12.1% per attached blastocyst and 1.3% per embryo cultured. High fragmentation was the single-trait group that showed the highest derivation rate (17.0% per attached blastocyst or 2.6% per embryo cultured). The MN group had the lowest derivation efficiency of the single traits and was the only single-trait group to not significantly differ from embryos with multiple traits for derivation efficiency per embryo cultured. Of the abnormally fertilized embryos, 2 lines originated from 1PN embryos and 1 from a 0PN embryo. In addition, there were no differences found between PQE groups or between plated blastocysts that did or did not make an hESC line for trophectoderm quality or level of blastocoel expansion.
All statistics were calculated using Fisher exact test: a vs. c, P < 0.05; b vs. c, P < 0.05; d vs. h, P < 0.005; e vs. h, P < 0.05; f vs. h, P < 0.05; g vs. h, P < 0.005.
hESC, human embryonic stem cell.
Characterization of new hESC lines originating from PQEs
Each of the 16 new lines showed high levels of alkaline phosphatase activity, stained positively for pluripotency markers including SSEA-4, TRA-1-60, and TRA-1-81, and were negative for SSEA-1 (Fig. 3). All lines were able to form EBs and ongoing histological assessment indicates differentiation into the 3 germ layers for those fully analyzed (Fig. 3). Karyotyping resulted in normal karyograms for all lines established and showed 10 of the lines were female and 6 were male. In addition to the normal hESC lines established, there were 7 additional hESC-like outgrowths that failed to generate stable hESC lines. Of these, a 1PN abnormal outgrowth was determined to be tetraploid and a 3PN outgrowth was triploid. The other initial hESC-like outgrowths (2 from fragmented embryos, 1 from an MN embryo, 1 from an additional 3PN, and 1 from an embryo with multiple traits) had increased differentiation and/or abnormally slow growth and were not able to be propagated enough for karyotype analysis or detailed characterization.

Examples of hESC characterization criteria for newly derived cell lines.
ICM cell number analysis shows a correlation with hESC derivation efficiency
As differences in hESC derivation efficiency were observed despite similar ICM morphology, we also wanted to determine if ICM morphology is an accurate indicator of ICM cell number and if PQEs produce ICMs with comparable cell numbers. An additional 345 PQEs were cultured and 121 blastocysts (35.1%) were produced. Good-quality ICMs were formed in 43.0% of blastocysts produced or 15.1% of embryos cultured, which was similar to the hESC derivation portion of this study. Eighty-eight blastocysts were successfully differentially stained to determine cell allocation patterns to the ICM. Good-quality (grade A and B) ICMs had significantly more ICM cells than poor-quality (grade C–E) ICMs (18.6 ± 16.9 vs. 9.1 ± 7.7; P < 0.001, chi square). Figure 4A shows the cell numbers of the 33 good-quality ICMs that were differentially stained. Although the mean cell number of these good-quality ICMs does not significantly differ by trait, 12.1% of the ICMs have >3 times the mean cell number of the good-quality ICMs, and surprisingly, this is the percentage of hESC derivation in this study (12.1%). These outliers are found in the high fragmentation, delayed development, and abnormal fertilization groups. Figure 4B and 4C show 2 differentially stained blastocysts with similar morphological ICM grades but incongruent ICM cell numbers.

Discussion
The deselection of embryos from clinical use is based on morphological observations during preimplantation development that are known to negatively correlate with successful implantation [24]. There are numerous poor-quality traits and each confers varying degrees of developmental impairment and risk for chromosomal aberrations, both singly and in combination with other traits [41]. As a potential source for the derivation of new hESC lines, this heterogeneous cohort of embryos has gained newfound significance in regenerative medicine. The first step in maximizing the value of this resource is to better understand the different types of embryos that comprise the PQE pool and the ability of each type to generate hESCs.
This study is the first to fully examine the influence of early embryo morphology on hESC derivation efficiency. By subdividing the PQE pool into groups based on traditional markers of poor implantation, we were able to determine that early embryo traits can influence the ability of good-quality ICMs to generate hESCs. The overall efficiency of hESC derivation in this study was 12.1% per attached blastocyst, but when stratified by poor embryo trait, efficiency ranged from 0% to 17%. This range of derivation efficiency indicates that PQEs are not a uniform population and that ICM quality alone is an incomplete predictor of hESC potential. Most notably, embryos with >1 poor-quality trait during early development were unable to generate new hESCs compared with those with a single poor trait (14.9% per attached blastocyst). These blastocysts were morphologically indistinguishable from the blastocysts originating from single-trait PQEs, were cultured in identical conditions, and yet were not responsible for establishing a single new hESC line.
The culture of embryos to the blastocyst stage is thought to provide a mechanism of self-selection for viable embryos [42]. Our overall blastocyst rate of 31.2% using fresh PQEs is similar to IVF-based reports such as those of Shoukir et al. [43] (37.8% for all 2PN PQEs), Hardy et al. [44] (39% for all PQEs including abnormally fertilized), and Balakier and Cadesky [30] (43% for MN embryos). In our study, embryos with multiple poor-quality traits had a significantly lower blastocyst formation rate and these blastocysts had significantly fewer good-quality ICMs. Because this group of embryos was unable to generate new hESCs lines, we suggest that good-quality ICM development does not overcome the developmental defects inherent in these embryos. These results show that blastocyst development and ICM quality alone are not reliable indicators of further viability.
Our findings are highly pertinent to the field of hESC derivation, where optimization of techniques and culture conditions are published regularly. In ART research, advances in embryo manipulation, culture, and selection have led to increased pregnancy rates and a reduced incidence of multiple births [45]. The validity of these studies is based upon groupwise comparison of embryos with equivalent developmental competence. Embryo quality, typically defined by multiple factors throughout preimplantation development, is one of the main determinants of randomization in ART experimental design. Despite this, the prognostic value of embryo grading used in IVF has not been applied to hESC derivation research [46]. Our results indicate that the currently used morphological indicators of embryo implantation can be successfully applied to hESC derivation. As the different types of PQEs in our study were unequal in their ability to derive hESCs, proper randomization based on early embryo traits may be necessary when planning experiments with hESC derivation efficiency as an endpoint. This may be especially helpful in maintaining reproducibility of results between laboratories using embryos of disparate quality. Whether variations in embryo quality account for the wide range in derivation efficiency reported between laboratories needs further examination.
It is estimated that over half of all IVF embryos produced are chromosomally abnormal and the incidence of these abnormalities increases with inferior embryo quality [5,47,48]. This should not, however, discourage the use of PQEs for hESC derivation, because PQEs form the most abundant source of embryos for research. Up to 42% of blastocysts originating from PQEs have been shown to be chromosomally normal [49]. In addition, successful derivation of euploid hESC lines from aneuploid blastocysts has been reported by several groups [50 –53]. These reports corroborate preimplantation genetic screening data showing that a high proportion of aneuploid embryos are actually mosaic mixtures of normal and chromosomally abnormal cells [54]. Hardarson et al. [49] found that if embryos with poor-quality traits during development result in abnormal mosaic blastocysts, there are an increased number of affected cells when compared with mosaic blastocysts from good-quality embryos. Nevertheless, euploid hESCs derived from aneuploid embryos point to an in vitro selection of normal cells for the establishment of hESCs. In addition, analysis of hESC lines derived from PQEs have shown to have no adverse affect on phenotype or differentiation ability after long-term culture [10,14]. We found 7 initial hESC-like outgrowths difficult to propagate to stable hESCs and 2 of these were confirmed to have abnormal karyotypes. In comparison, the 16 euploid hESC lines derived in this study followed typical hESC self-renewal and remained undifferentiated in culture for extended periods. Despite their correlation with increased aneuploidy during IVF, our results further validate the use of PQEs for the derivation of karyotypically normal hESCs.
In this study, 12.1% (4/33) of the differentially stained blastocysts with good-quality ICMs had 3 times the average ICM cell number. This outlier group may have a better chance of establishing hESCs than the average ICM, possibly even if they were mosaic for aneuploidy. Interestingly, the occurrence of these high-cell-number ICMs was the same as our overall derivation efficiency (12.1%) and they originated from the 3 PQE groups most competent at establishing hESCs. Whether there is a threshold number of euploid ICM cells needed for the generation of a new hESC line is unknown. We were, however, able to confirm that good-quality ICMs have twice the number of cells when compared with poor-quality ICMs, possibly explaining their increased efficiency at deriving hESCs. With this in mind, a promising option for increasing the efficiency of the hESC derivation process may be to enhance ICM cell numbers by adaptation of culture conditions. Human embryos, for example, had an increase in ICM cell number by 35% and 60% when cultured to the blastocyst stage with media augmented with granulocyte–macrophage colony-stimulating factor or insulin-like growth factor-1, respectively [55,56]. Our study has demonstrated the feasibility of using the abundant supply of PQEs for experiments aiming to improve ICM cell number and increasing hESC derivation efficiency.
In conclusion, PQEs are an abundant and viable source for establishing new hESC lines and can be used for improving hESC derivation efficiency. However, this study has shown that PQEs are a heterogeneous population with varying levels of developmental impairment. Our results indicate that the morphological indicators of embryo grading as used in ART can be successfully applied to hESC derivation. We found that embryos with multiple cleavage-stage abnormalities were not able to establish new hESCs despite their ability to develop to blastocysts with good-quality ICMs. In addition, a correlation between ICM cell number and hESC derivation suggests that increasing ICM cell number could increase derivation efficiency. Further attempts to optimize hESC derivation culture, techniques, or efficiency need to incorporate early embryo morphology in addition to ICM quality for the comparison of groups with equal developmental competence.
Footnotes
Acknowledgments
T.O. is supported by a research grant from the Bijzonder Onderzoeksfonds of the Ghent University, Belgium (grant no. BOF O1J15707). S.C.d.S.L. is supported by The Netherlands Organization for Scientific Research (grant No.: VENI 916.76.015). P.D.S. is a holder of a fundamental clinical research mandate by the Flemish Foundation for Scientific Research (FWO-Vlaanderen).
Author Disclosure Statement
All authors have no actual or potential conflicts of interest to disclose.