
Abstract
The contribution of stem cells derived from adult tissues to the recovery of pancreatic islets from chemical injury is controversial. Analysis of nonhematopoietic differentiation of bone marrow-derived cells has yielded positive and negative results under different experimental conditions. Using the smallest subset of bone marrow cells lacking immuno-hematopoietic lineage markers, we have detected incorporation and conversion into insulin-producing cells. Donor cells identified by genomic markers silence green fluorescent protein (GFP) expression as a feature of differentiation, in parallel to expressing PDX-1 and proinsulin. Here we elaborate potential experimental difficulties that might result in false-negative results. The use of GFP as a reporter protein is suboptimal for differentiation experiments: (a) the bone marrow of GFP donors partially expresses the reporter protein, (b) differentiating bone marrow cells silence GFP expression, and (c) the endocrine pancreas is constitutively negative for GFP. In addition, design of the experiments, data analysis, and interpretation encounter numerous objective and subjective difficulties. Rigorous evaluation under optimized experimental conditions confirms the capacity of adult bone marrow-derived stem cells to adopt endocrine developmental traits, and demonstrates that GFP downregulation and silencing is a feature of differentiation.
Introduction
A
Assessment of the capacity of stem cells from the adult to differentiate, trans-differentiate, dedifferentiate, and convert to other cell types has been overloaded by extensive reports of negative results, largely considered to negate or at least question the validity of positive findings. Further, these data extend to the question whether pluripotent stem cells persist in the adult organism, or are largely restricted to tissue progenitors. Considering that the pancreas is a particularly difficult tissue to determine incorporation of cells tagged with green fluorescent protein (GFP) as a reporter protein, the burden of proof stands on the negative findings. Our analysis raised several possible causes that might impede the detection of grafted cell differentiation to produce insulin in the injured islets.
Materials and Methods
Animal preparation, diabetes, and transplantation
Mice used in this study were C57Bl/6J (B6, H2Kb, CD45.2), B6.SJL-Ptprca Pepcb/BoyJ (H2Kb, CD45.1), and C57BL/6-TgN(ACTbEGFP)1Osb (GFP, H2kb, CD45.2) purchased from Jackson Laboratories and housed in a barrier facility. The Institutional Animal Care Committee approved all procedures. Diabetes was induced in female mice aged 6–8 weeks by 5 daily (days −6 to −2) consecutive intraperitoneal injections of 60 μg/g Streptozotocin (STZ; Calbiochem-Merk). STZ was diluted in phosphate-buffered saline (PBS; Beit Haemek) and was used within 15 min of preparation. Mice displayed blood glucose levels above 250 mg/dL within 10–14 days at this STZ dosage and schedule of administration. Recipients were sublethally irradiated at 675 rad (total body irradiation) using an X-ray irradiator (RadSource 2000) at a rate of 106 rad/min (day −1). Cells suspended in 0.2 mL PBS were transplanted into the lateral tail vein.
Cell isolation
Whole bone marrow cells (BMC) from wild-type or GFP-positive male donors aged 6 weeks were harvested and elutriated as previously described [23]. Similar fractions of GFP expression (36%–58%) have been detected in the bone marrow of mice aged 4–12 weeks. Briefly, BMC from 8 to 10 donors were loaded into the chamber of a counterflow centrifuge (Beckman Instruments) operating at a constant speed of 3,000 rpm. Fractions were collected in 200 mL at elutriation flow rate of 25 mL/min to isolate the smallest subset of nucleated cells (Fr25), and the largest cells were collected in the rotor off position (R/O). Fr25 cells were lineage-depleted using rat-anti mouse monoclonal antibodies against CD5 (clone 53–7.3), GR-1 (clone RB6-8C5), Mac-1 (clone M1/70), and B220 (clone RA3-3A1/6.1) extracted from hybridoma cell lines (ATCC) and purified TER119 (eBioscience). Antibody-bound cells were incubated with goat-anti-rat secondary antibodies conjugated to magnetic beads at a ratio of 4–5 beads per cell (Dynal Biotech), and lineage-negative (lin−) cells were collected by exposure to a magnetic field.
Flow cytometry
Measurements were performed with a Vantage SE flow cytometer (Becton Dickinson) in samples that underwent red cell lysis, after blocking nonspecific binding with 1 μL mouse serum. Pan-hematopoietic marker was detected with antibodies against minor antigens CD45.1 (clone A20) and CD45.2 (clone 104; eBioscience).
Tissue preparation
Mice were sacrificed by CO2 asphyxiation. A blunt 20 G catheter was placed into the left ventricle and the inferior vena cava was sectioned. Exsanguination was performed by infusion of 30 mL of ice-cold PBS containing 1.5% fresh paraformaldehyde and 0.1% glutaraldehyde using a mini-perfusion pump (at a rate of 2.5 mL/min). Excised pancreata were placed in this medium for 2 h at 0°C–4°C for additional fixation, and immersed in 30% sucrose overnight. Tissue embedded in OCT (Sakura Finetek) was frozen in isopentane suspended in liquid nitrogen and sectioned (3–6 μm) with a Cryotome (Termo Shandon). Cryosections were stored at −80°C before analysis.
Immunofluorescence and fluorescence in situ hybridization
Donor cell analysis in host pancreata was performed in 2 steps to include immunofluorescence in stained frozen sections and fluorescence in situ hybridization. Sections were fixed in acetone for 10 min at −20°C and permeabilized by 10 min incubation with 0.2% Saponin, 1% bovine serum albumin, and 0.1% Triton-100, then stained with primary antibodies for 1 h, washed, and counterstained with respective secondary antibodies for 30 min at room temperature. Nuclei were labeled with Hoechst-33342 (1:1,000; Molecular Probes), and sections were mounted in antifade medium (Dako Glostrup) and air-dried. Serial cryosections from fixed pancreas were immunostained with primary antibodies: polyclonal mouse-anti-proinsulin (1:20; R&D Systems), goat-anti-PDX-1 (1:5,000, kindly provided by Shimon Efrat), rat anti-Pecam-1 (CD31, 1:100, CBL 1337; Chemicon), biotinylated anti-mouse CD45 (1:100; Biolegend), biotinylated donkey-anti-mouse (1:1,000), and rabbit-anti-GFP (1:100; Santa Cruz Biotechnology). These primary antibodies were counterstained with fluorescein isothyocyanate-labeled donkey-anti-rabbit (1:200; Jackson Immunoresearch) and goat-anti-rat monoclonal antibodies (1:200; Santa Cruz Biotechnology), Alexa Flour 568-conjugated donkey-anti-goat (1:500), and Cy3-conjugated rat-anti-goat (1:200; Molecular Probes). Biotinylated primary antibodies were counterstained with Cy3-conjugated Streptavidin (1:400) and Cy5-conjugated Streptavidin (1:500) both from Jackson Immunoresearch.
X and Y chromosomes were painted in fresh sections or immunostained slides after treatment with 0.025% pepsin for 30 min at 37°C. Slides were rinsed 1 min in distilled water and 5 min in 2 × SSC; dehydrated by sequential immersion for 2 min 70%, 85%, and 100% ethanol at room temperature; and air-dried. A second fixation in 70% formamide at 72°C for 2 min was followed by exposure to cold 70% ethanol and then dehydration by immersion in graded ethanol solutions. Nuclear probes (Applied Spectral Imaging; Migdal Haemek) were denatured at 74°C for 7 min and at 37°C for 30 min. Probe hybridization was performed at 37°C overnight, and then slides were sequentially washed in 0.4 × SSC stringency solution at 74°C and 2 × SSC solution supplemented with 0.1% NP4O detergent at room temperature. Slides were washed and mounted with antifade containing diamidino-2-phenylindole (DAPI). In our hands, Y chromosome was detected using a painting probe in 77% ± 14% of islet cells in male controls (∼400 islet cells).
Images were acquired with an Axioplan 2 (C. Zeiss) fluorescence microscope equipped with an Apotome and with a C. Zeiss confocal laser scanning microscope using AxioVision 4.5 and LSM 510 software, respectively. Images were pseudocolored and RGB reconstructed using Adobe Photoshop software.
Results
Positive results
Our analysis focuses on the capacity of cells derived from the adult bone marrow to adopt pancreatic endocrine differentiation traits in vivo. The smallest subset of nucleated BMC is endowed with durable multilineage hematopoietic potential [23 –26], differentiates into hepatocytes [27], and contributes to recovery of multiple epithelial tissues [28]. Multiple doses of STZ were used to induce chemical diabetes in B6. Boy female mice (CD45.1, days −6 to −2), and sublethal irradiation at 675 rad (day −1) was administered as additional injury and considering the possibility that cells might migrate to the bone marrow before incorporation in the pancreas (Fig. 1A). STZ administration was adjusted to yield partial islet destruction and allow survival for extended periods of time with hyperglycemia (415 ± 82 mg/dL at day 100), considering that residual tissue is necessary for assessment of cell incorporation. Diabetic mice were grafted with the smallest nucleated fraction of counterflow centrifugal elutriation from male GFP+ donors (CD45.2, day 0), providing triple mismatch markers for cell identification (male CD45.2+GFP+ → female CD45.1+GFP−). Transplantation of female R/O hematopoietic progenitors from wild-type mice (GFP−) reduced blood glucose levels, and additional recovery of glucose homeostasis was observed after transplantation of the smallest nucleated fraction depleted of cells expressing lineage markers (Fr25lin−) (Fig. 1B). Plasma insulin levels were elevated in recipients of Fr25lin− cells that displayed near-normoglycemic blood glucose levels at the experimental end point (P < 0.01 vs. mice without cell transplant). Male (Y chromosome) cells were detected in the pancreas, expressing PDX-1 and proinsulin (Fig. 1C). Pancreato-duodenal hoemobox-1 (PDX-1) is an essential transcription factor in developing and functional β cells [29], and proinsulin is a better index of hormone production within the cells, removing the doubt of insulin uptake from the surrounding injured islets [30]. Our fluorescence in situ hybridization analysis detects 68% ± 9% of male cells with low nonspecific false-positive staining (Fig. 1D).
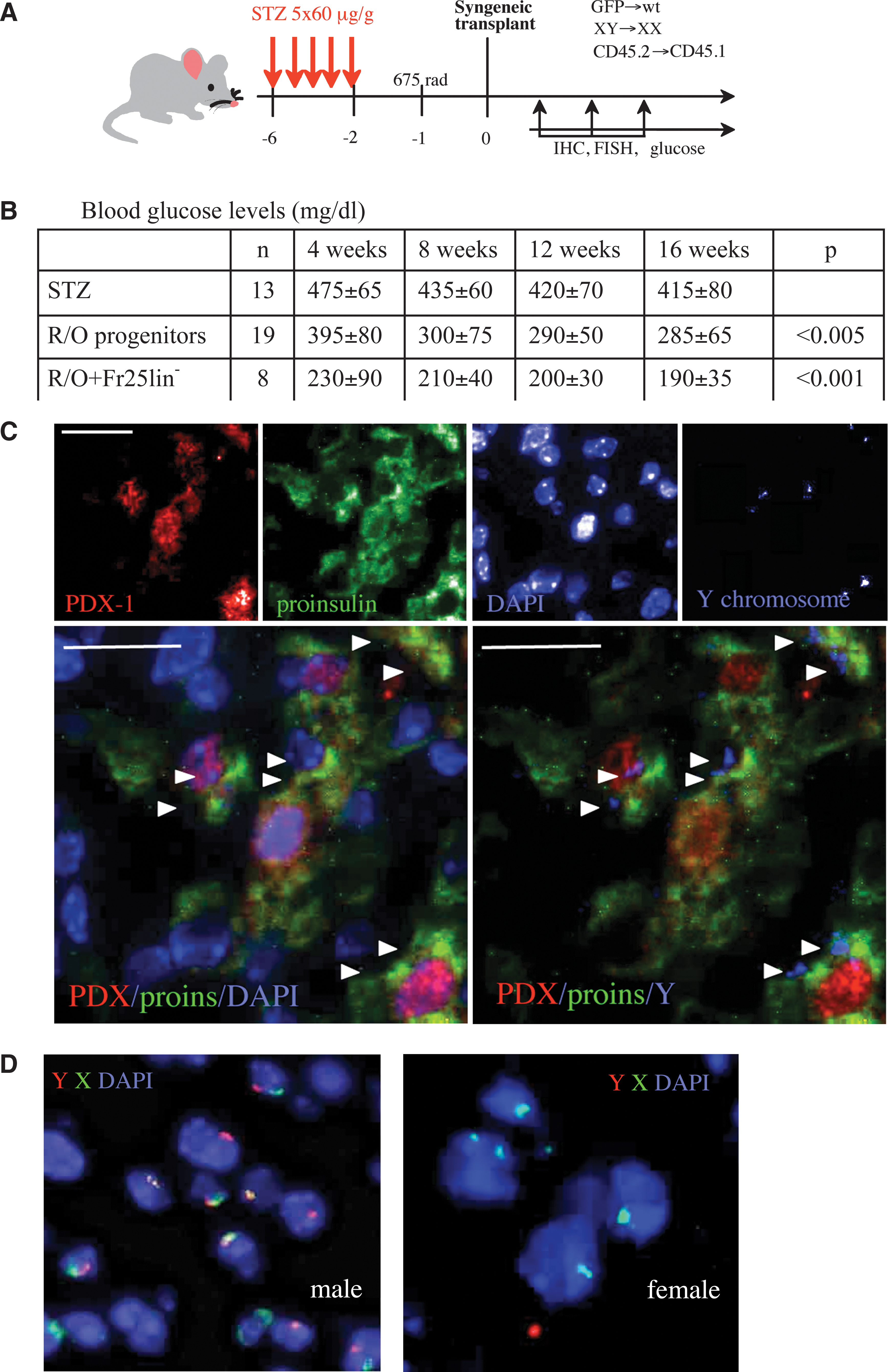
Donor cell identification in injured islets.
Negative results
Numerous prior studies [31 –36] have pointed out the capacity of grafted GFP+ cells to home to the injured pancreas, with preferential affinity to the pancreatic ducts, blood vessels, and islet perimeter (Fig. 2A). While observer's attention is attracted by the GFPbright cells surrounding the islets (Fig. 2B), exclusion of these cells with intense fluorescent signal from the field of analysis (Fig. 2C) allows the observation of GFPdim cells in central regions of the injured islets (Fig. 2D). It is plausible that differentiation will take place within the inner regions of the islets, where the injury signals and possibly inductive factors are at maximal intensity.
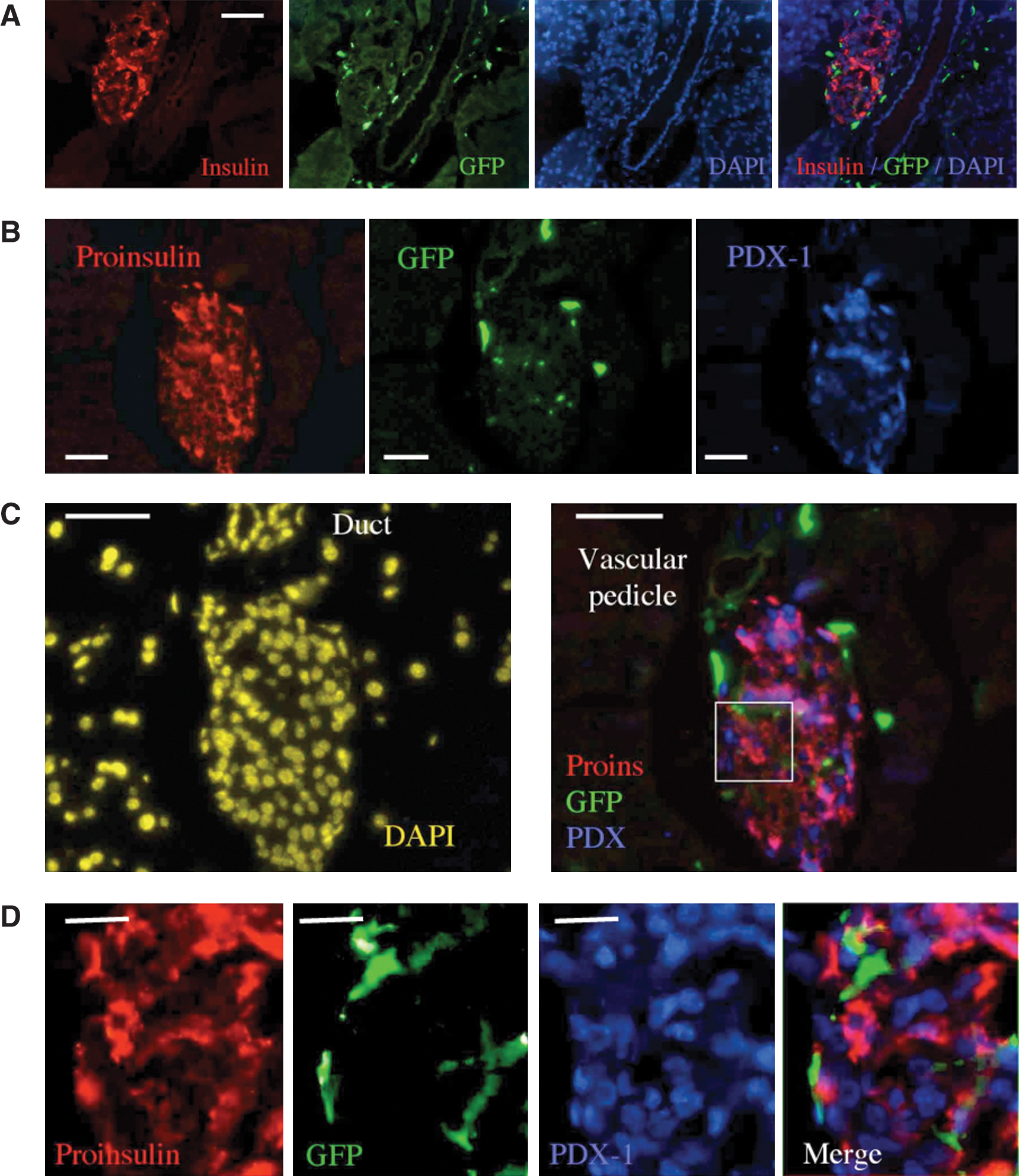
Analysis of GFP expression in islets of mice with chemical diabetes after irradiation and transplantation of the smallest bone marrow subset from GFP donors.
High-resolution analysis and color conversion
Positive identification of GFPdim cells within in the inner regions of the islets rich in PDX-1 and proinsulin allows further assessment of intracellular co-localization by confocal microscopy. Such analysis demonstrates that proinsulin co-localizes with dim GFP signals, whereas the hormone precursor is not found in GFPbright cells (Fig. 3A). The extensive use of pseudocolored layers for image reconstruction and analysis has drawn attention to a change in color as evidence of co-localization of 2 molecules. As such, both GFP and proinsulin reside in the cytosol and pseudocolored red and green layers are expected to convert to a neutral yellow (Fig. 3B). Such color mixtures are often observed; however, juxtaposition of labeled molecules may not result in color conversion, because of either dominant signal of one layer or sequestration of the markers in different regions of the cell.
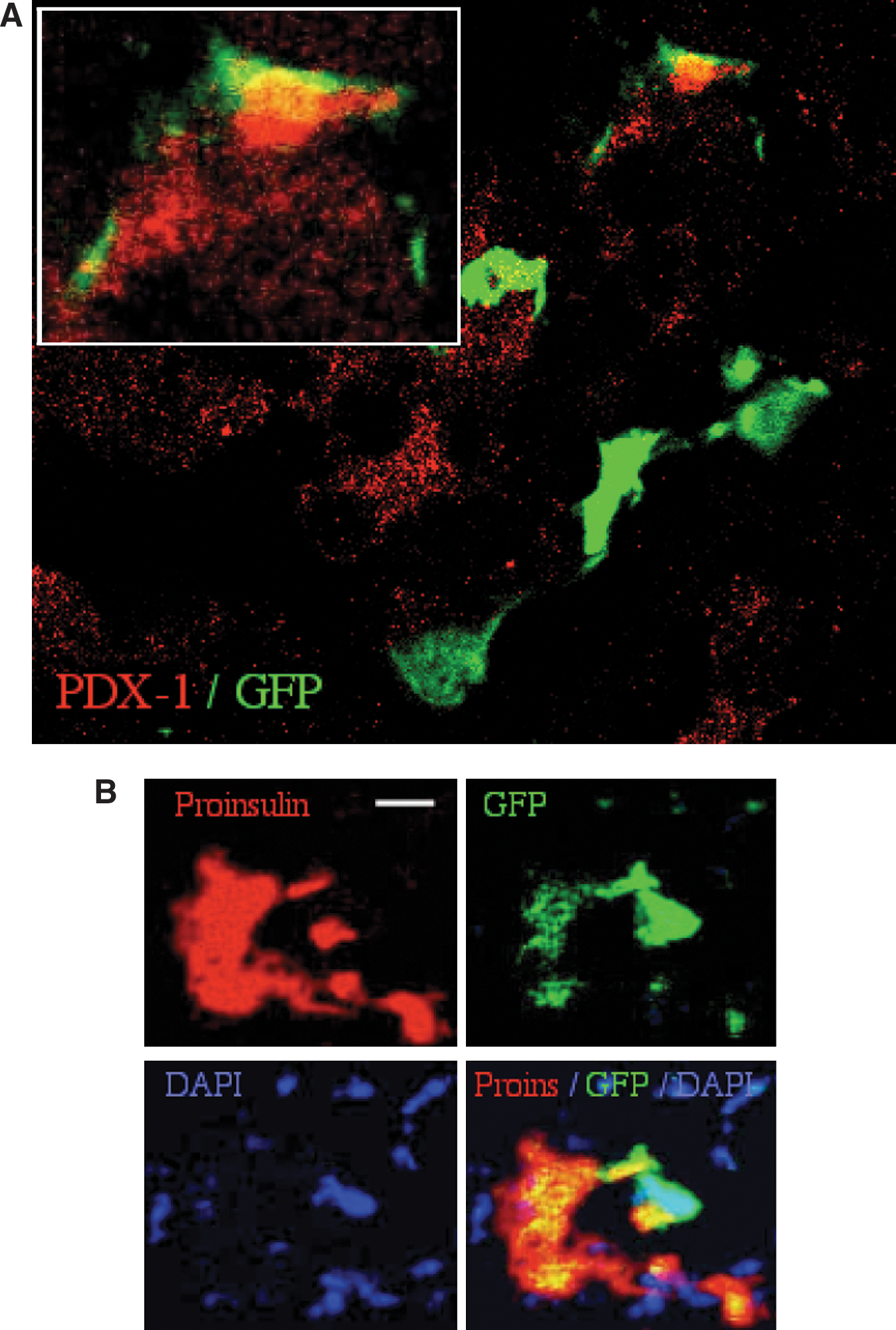
Qualitative analysis of GFP+ donor cells in injured pancreatic islets.
Positive identification of donor cells
Incidental incorporation of GFP+ donor cells in injured islets (Fig. 4A) has been repeatedly reported under various experimental protocols [31,32,37,38]. However, downregulation of GFP in differentiating cells [39 –41] calls for rigorous analysis of tissue samples using a reliable marker, such as Y chromosome in sex-mismatched transplants [42,43]. While focusing on 4 GFPbright islet-infiltrating cells (Fig. 4A), unequivocal detection of donor cells using a genomic marker (male to female transplants) reveals GFP silencing (Fig. 4B). The insets demonstrate 7 GFPbright Y+ cells, 9 GFP−Y+ cells, 1 GFPdimY+ (inset 2), and 1 GFPbrightY− cell, consistent with GFP silencing and downregulation in 59% of donor male cells incorporated in the islets. Further, clear distinction can be made between 7 GFPbrightY+PDX-1− cells located mostly at the islet perimeter and 4 GFPdim/−Y+PDX-1+ cells in central islet regions (insets 1 and 2), which represent 17%–22% of islet infiltrating donor cells identified by GFP, Y chromosome and both. This is the higher end of incorporation of donor cells in the injured islets, the lower end being complete absence of donor cells. This analysis is representative of a general characteristic of GFP downregulation and/or silencing in islet infiltrating cells that express endocrine pancreatic markers, which might be used a as a criterion of differentiation.
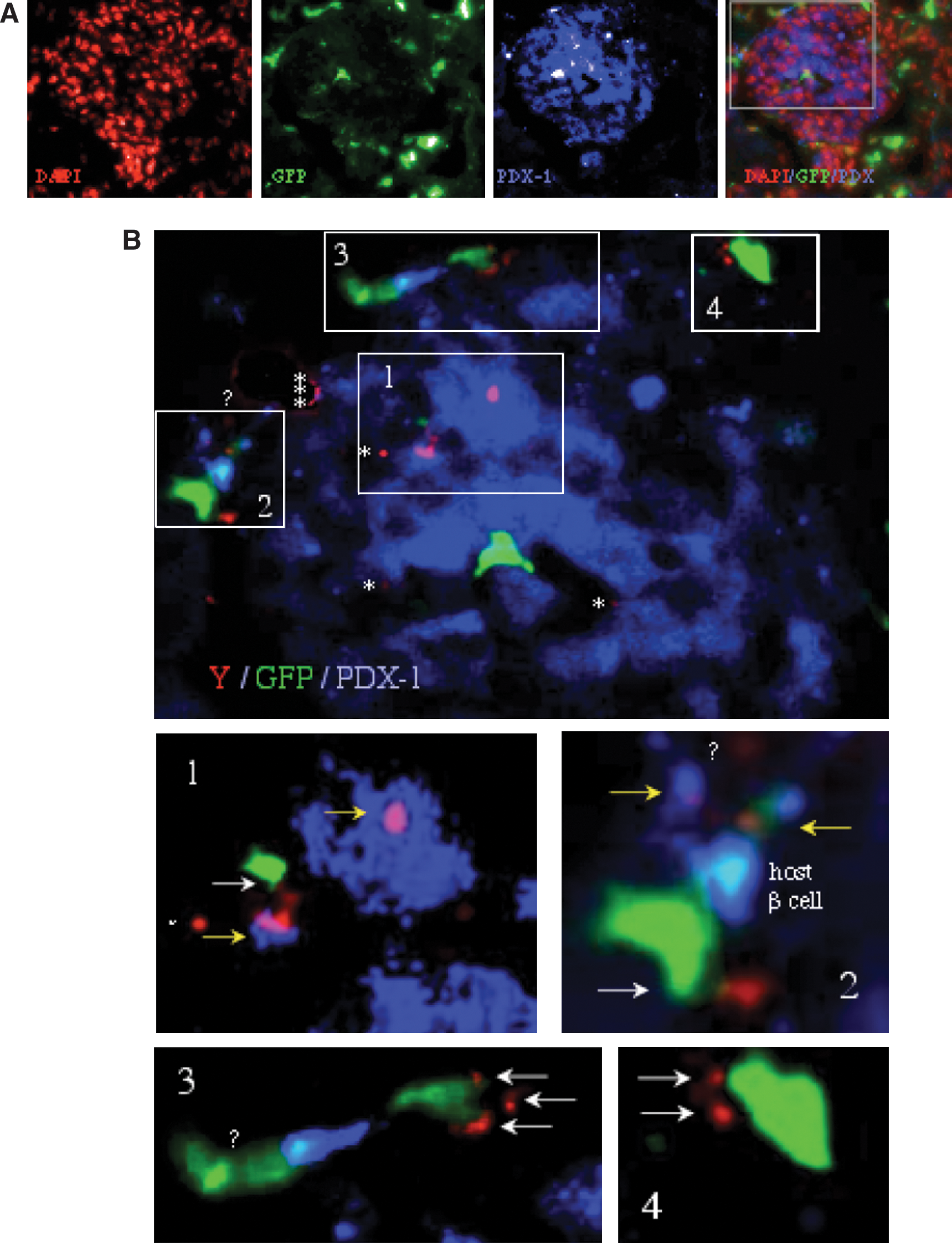
GFP is silenced in differentiating cells.
Objective difficulties in pancreatic islet analysis
Chemical injury to the islets is a prevalent model in experimental attempts to reconstitute β cell mass. Injury caused by STZ results in variable changes in islet infrastructure and function, accompanied by marked heterogeneity in expression of the specific markers (Fig. 5), as also observed with alloxan-induced injury (not shown). Islet structure is distorted by cell loss, with some islets displaying prominent downregulation of proinsulin (in ∼44% of nucleated cells) and insulin (in ∼40% of nucleated cells) production, and PDX-1 expression (∼23% of nucleated cells). It is unclear whether the higher incidence of cytosolic PDX-1 reflects difficulties in processing of the fragile samples from STZ-treated mice or intrinsic cellular reactions to injury. Under these conditions it is expected that changes in islet architecture will impose adverse influences on the behavior and differentiation of candidate stem cells according to their location. Newer models of toxin receptor expression for targeted β cell depletion [4] and disrupted proinsulin processing [44] are elegant alternative models to induction of hyperglycemia with a minor inflammatory component.
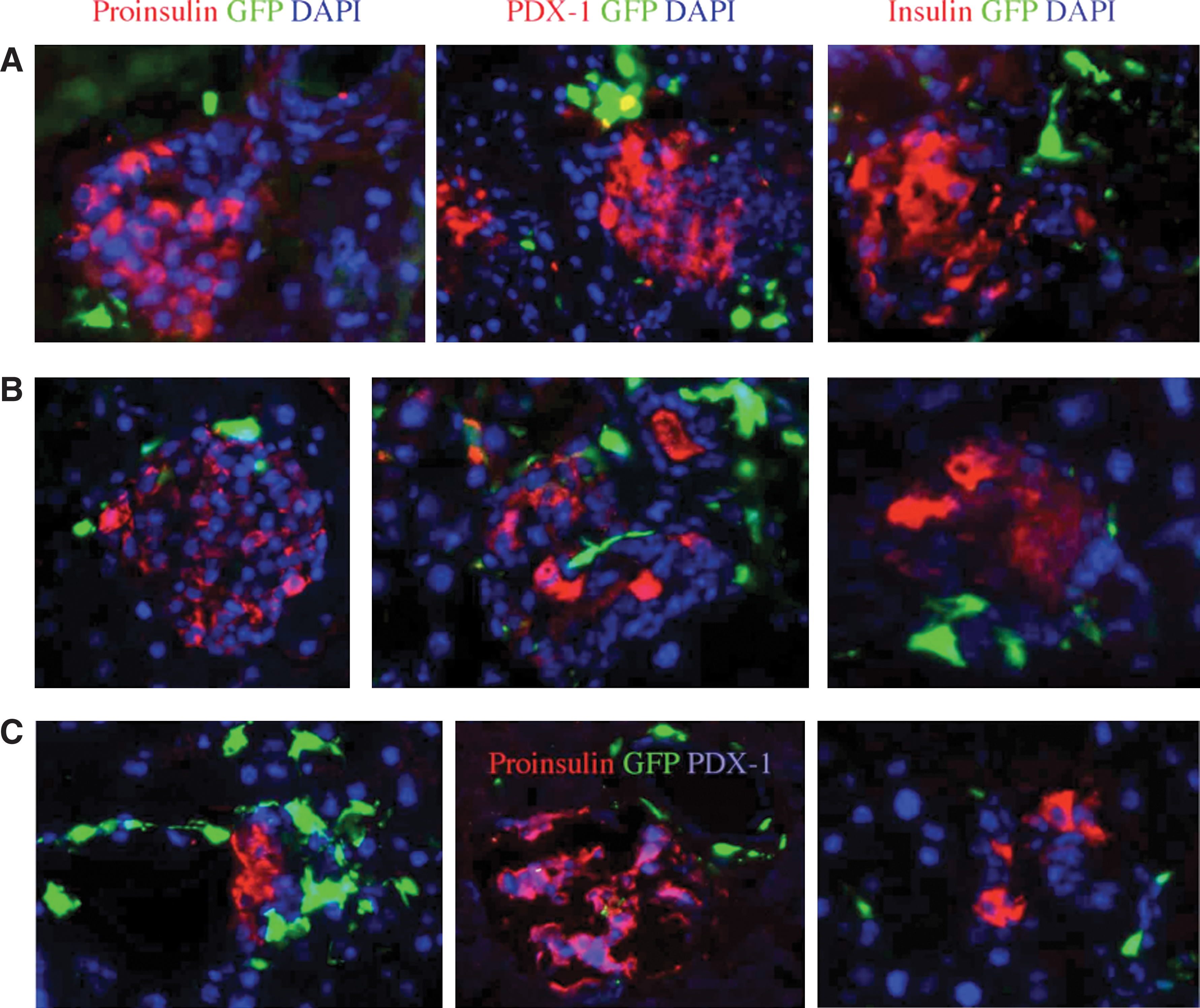
The injury factor. Representative RGB reconstructed images of islets stained for insulin, proinsulin, PDX-1, and diamidino-2-phenylindole (DAPI) to demonstrate
Subjective difficulties in pancreatic islet analysis
The human factor has a major impact on tissue processing, observation, and data interpretation. Prior studies [31 –36] show preferential homing of GFPbright cells to pancreatic ducts, blood vessels, and the islet perimeter. This is likely a result of focused attention of the observer on GFPbright cells that surround the islets (Fig. 6A). BMC are frequently detected in the walls of pancreatic ducts and contribute to revascularization after injury (Fig. 6B). As much as rigorous interpretation rules are applied and standardized, the final step relies on subjective interpretation of the data, which is not always easy (Fig. 6C).
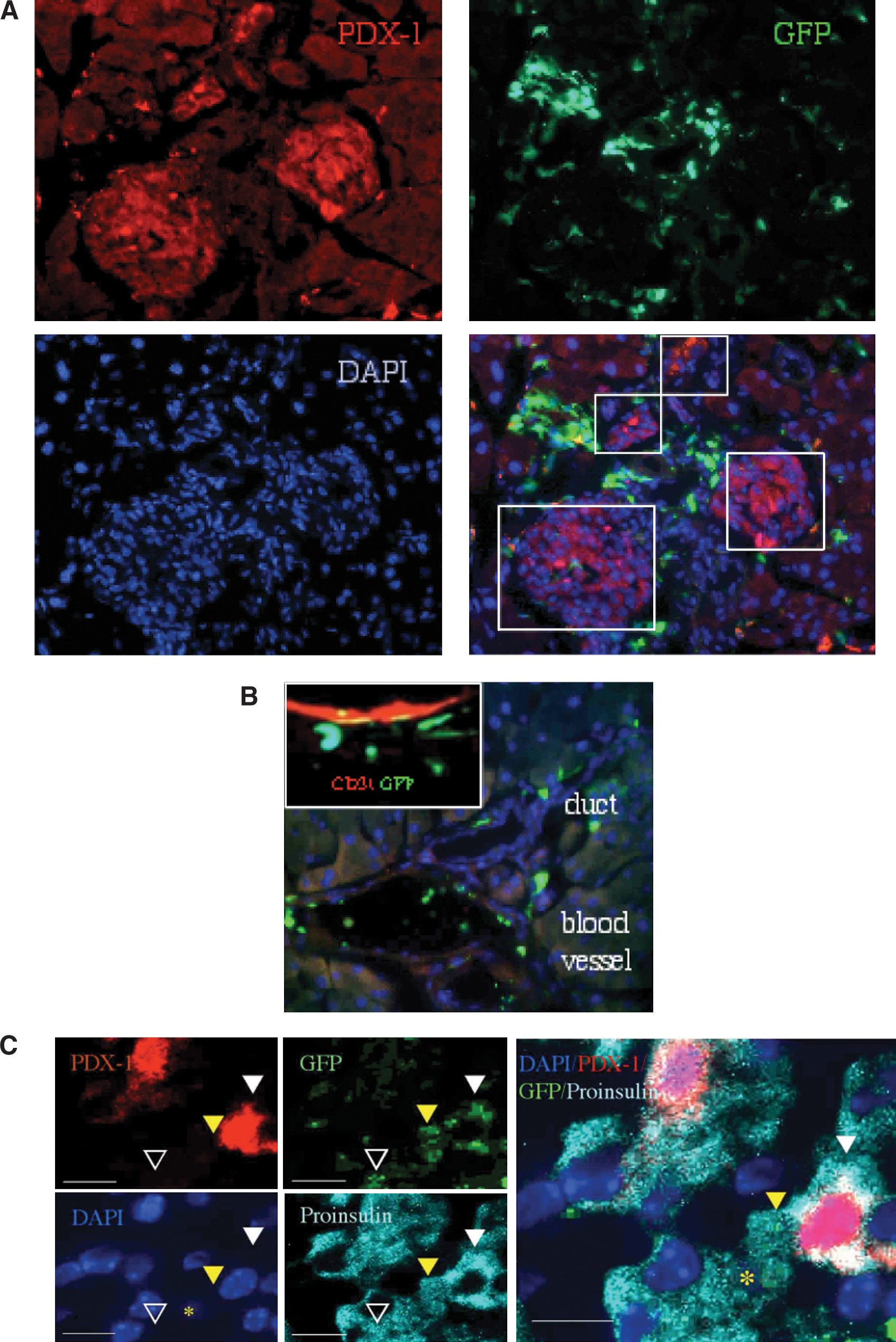
Objective and subjective factors in data analysis.
Fusion
Stem cells have been shown to fuse with parenchymal cells and adopt a tissue-like phenotype without undergoing differentiation under various experimental conditions [45 –47]. Using bone marrow-derived cells, the possible fusion partners of β cells could be either stem cells or their myeloid progeny [48]. CD45 is largely absent in the central regions of the islets under our experimental conditions; however, immunohematopoietic cells are detected in the islet perimeter. On rare occasions we observed formation of syncitia composed of several CD45+ cells at an extra-islet location, suggestive of participation of immune cells in clearance of tissue debris after injury (Fig. 7). Further, the pattern of scattered particles containing proinsulin is suggestive of degradation and phagocytosis of the degrading cells. We could not detect fusion products resulting in cells containing host–donor or donor–donor multiple sex chromosomes. It is unlikely that such rare events will be detected in analysis of tissue samples, nor could these account alone for tissue regeneration mediated by stem and progenitor cells [49].
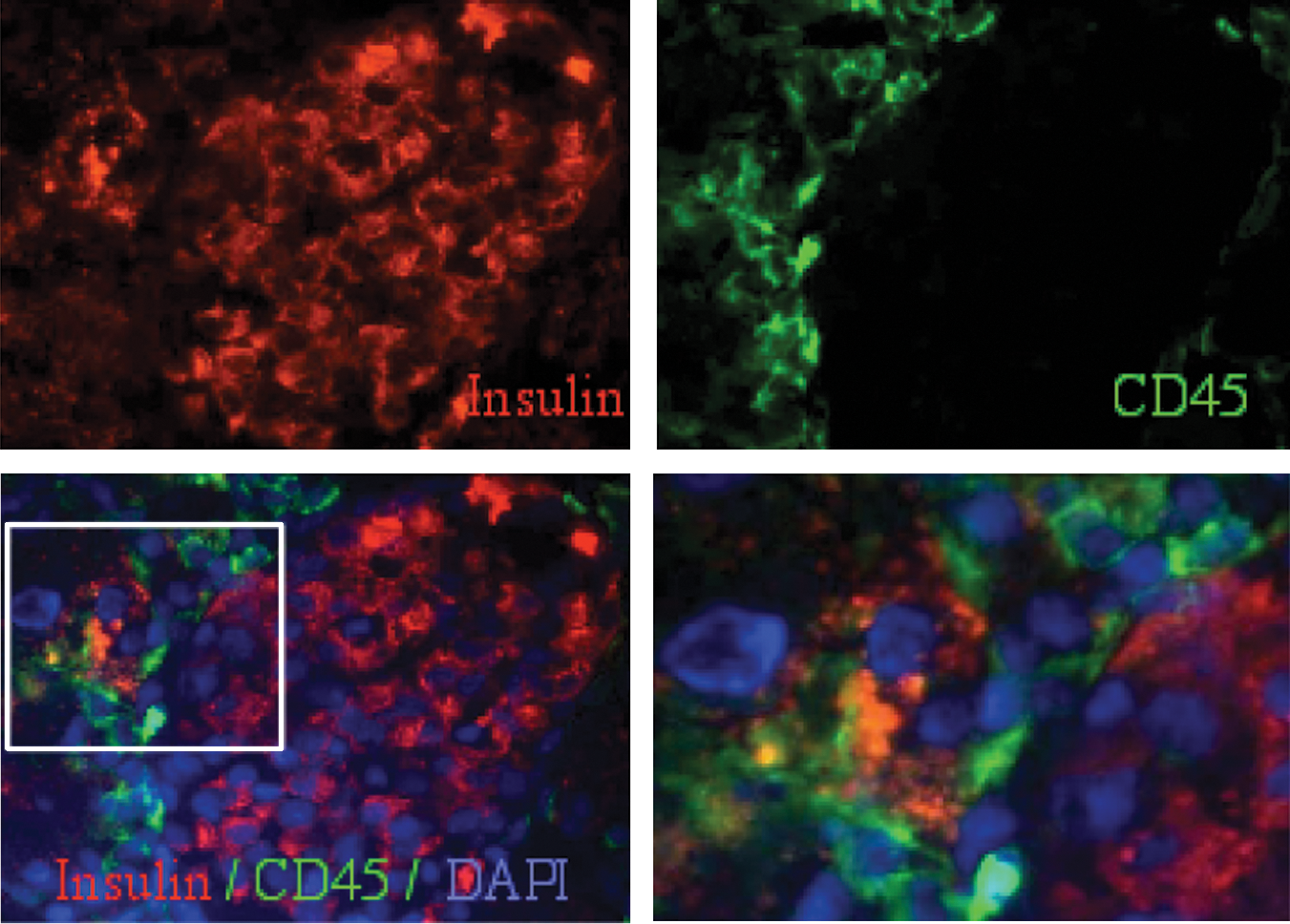
Insulin-producing donor cells lack hematopoietic markers. Morphology of debris clearance is demonstrated by a rare observation of a number of CD45-positive cells engulfing particles staining positive for insulin at the islet perimeter (scale bar 20 μm). Enlarged inset (right lower panel): the scattered appearance of insulin at the extra-islet location is characteristic of cell degradation.
Discussion
GFP is a useful and valuable marker with wide implementation in tracing experiments. For the particular purpose of proof of concept of stem cell behavior, this might be a liability rather than a useful tool. The type of experiments addressed here present difficulties in assessment of the functional incorporation of adult BMC into pancreatic islets. First, mice expressing GFP under control of the chicken β-actin promoter coupled with a cytomegalovirus enhancer display variable tissue-specific fluorescence [40,50]. GFP expression in the bone marrow encompasses 40%–70% of the nucleated cells [50], a distribution sustained in the fraction of cells isolated by elutriation (Fig. 8A). Second, lineage tracing in hematopoietic differentiation reveals partial expression of the reporter protein in primitive stem cells and developing progenitors [40], with similar variable distribution in the liver [39] and brain [41]. Incorporation of artificial chromosomes showed that promoter silencing is independent of location, and activation of the telomerase reverse transcriptase promoter requires histone deacetylation [51]. Silencing of transgene expression remains unsolved [52] even if the tissue is properly processed [53] and inspected [54], GFP is immunostained [55], the fluorescent signals are amplified [41], and lack of fluorescence is overcome by protein analysis [56], single-nucleotide DNA polymorphism [57], and detection of mRNA encoding GFP by amplified in situ hybridization [58]. Downregulation of GFP expression is similar to suppression of β-galactosidase in cells from ROSA26 mice [42,59], which has also provided negative results of islet reconstitution [60], and may be caused at the epigenetic level by methylation [61]. New approaches have achieved stable transgene expression by flanking the sequence with tandem copies of DNA insulators to protect from influences of neighboring enhancers [62]. Third, islets of GFP mice are dim to negative in fluorescence (Fig. 8C), evidence of specific physiological silencing of the reporter protein in this particular lineage [50]. The suboptimal experimental approach evolving from these considerations is the use of unevenly labeled cells to determine a differentiation process that often downregulates the transgene in a tissue that always silences the reporter protein. This phenomenon is not species specific, as cells from GFP rats equally silence expression [54]. Gene silencing in the particular transition of BMC to adopt various differentiation traits is the likely consequence of epigenetic factors that modulate the chromatin [63], such as methylation patterns [61]. Under these conditions, the absence of GFP might be rather considered as evidence of cell differentiation to adopt a pancreatic endocrine phenotype. A possible solution is the use of cells from transgenic mice expressing GFP under control of the insulin promoter [44,64]. Although the islets of these mice fluoresce, the use of GFP as an identifier of insulin expression in bone marrow-derived cells awaits confirmation.
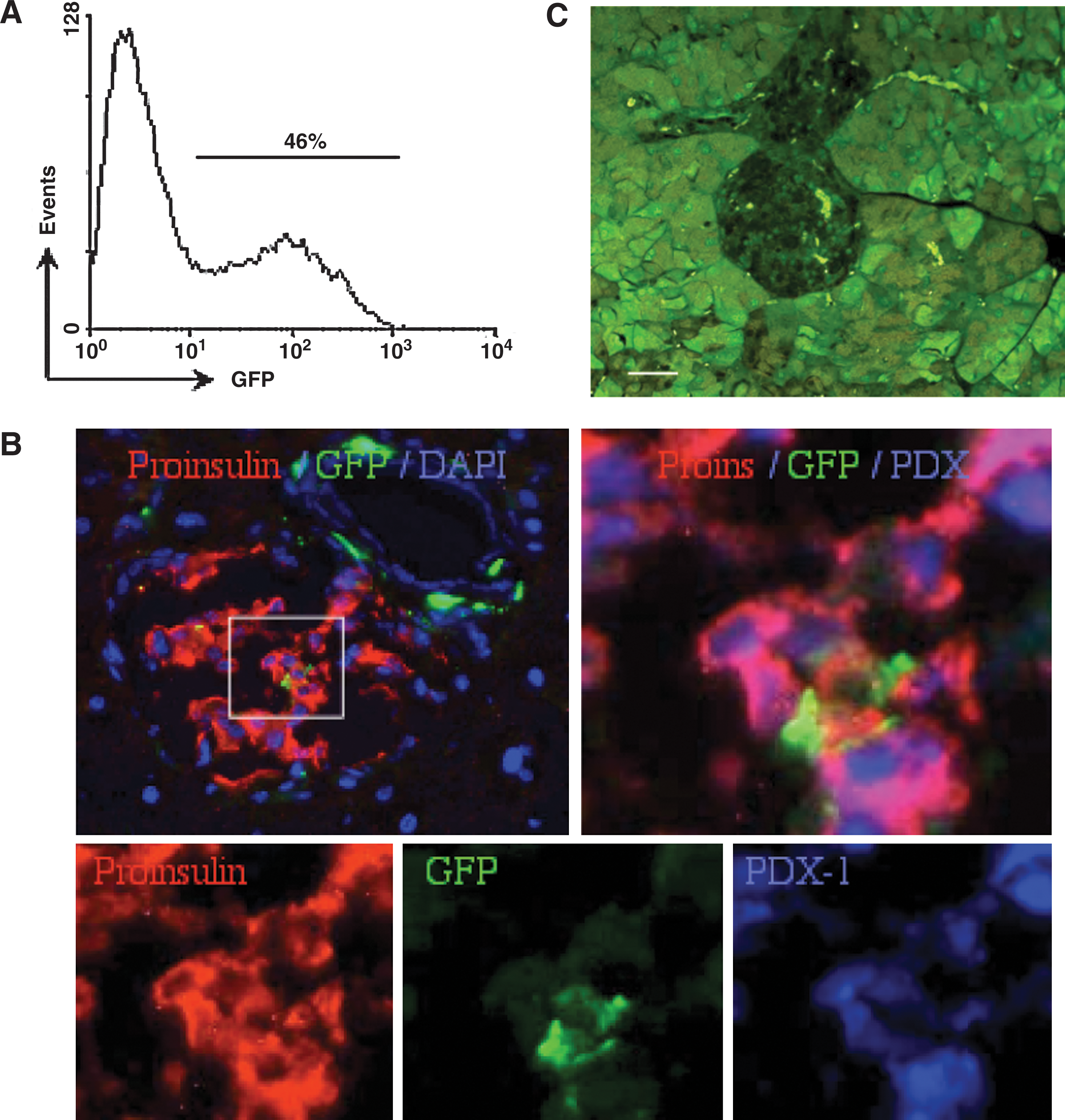
A suboptimal experimental setting is composed of the use of cells with incomplete reporter gene expression, to observe differentiation that silences the reporter gene expression in a tissue negative for the reporter protein.
Additional experimental difficulties are to be solved for differentiation experiments, including the temporal sequence of injury and cell transplantation, the timing of analysis, and the nature of cells used to reconstitute the endocrine pancreas. Such reconstitution experiments often require severe injury in the experimental setting [4,65], whereas in its absence there is no need for such incorporation [66]. Periods longer than 4–6 weeks are often required to observe differentiation of BMC in the pancreas [38,43]. Most important, the experimental outcome is determined by the nature of potent cells, which might be rare [38,67] and lack identifiable markers [68,69]. Negative selection by depletion of known progenitor markers [70] might well improve the efficiency of detection by removing the activity of progenitors.
Negative and positive results are equally important in qualitative analysis of the likelihood of biological processes to occur. One argument remains to be established, whether the rigorous analysis used in positive reports of an observed process have been adopted by the “failure to” and “little evidence of” reports. Experimentalists are often required to modulate the conditions to allow biological phenomena to be observed and analyzed. Since most experiments are being performed under nonphysiological conditions to reveal and highlight a particular phenomenon, we may have conceded to manipulation rather than observation. In the regular experimental practice, failure to observe a biological phenomenon suggests that it indeed does not occur, or our experimentation is not planned, performed, or analyzed at the desired accuracy and with proper controls. This is particularly relevant to repopulation of injured islets, which might be attained by multiple processes occurring in different cell populations [2 –22]. After the initial observation of functional incorporation of CD45-negative cells originating from the spleen in the pancreas under the immunomodulatory effect of Freund's Complete Adjuvant [71], a series of reports have challenged the significance of cell incorporation as a mechanism relevant to islet recovery [56,57,64]. The subsequent stimulating discussion and challenging experimentation have yielded significant insights into the process of islet remodeling and regeneration [42,43,59], including the question of adult stem cell plasticity. While some bone marrow-derived cells are endowed with differentiation capacity outside (and including) the hematopoietic compartment [27,28,33] prospectively selected cells bearing hematopoietic stem and progenitor markers lack this capacity under certain experimental conditions [32,34,36,70]. Using GFP as the only reporter protein, the experimental considerations delineated above would make incorporation of donor cells in the pancreas an unlikely outcome. The only study that used a genomic marker detected no donor cells in the islets after chemical injury and partial pancreatectomy [34], whereas a Cre/Lox system revealed wide incorporation of BMC in the islets in the absence of specific injury [33]. It would take much more effort to generalize such negative reports to the plastic developmental capacity of adult cells, in particular because the requirement for stringent analysis to avoid possible bias [72] has been largely met to present positive results [73]. Analysis of the incorporation of elutriated fractions derived from the adult bone marrow revealed significant contribution to islet recovery and restoration of glucose homeostasis [Iskovich S, unpublished data].
In summary, analysis and interpretation of stem cell behavior should account also for negative results that are important to set the limits of the potential of various subsets to replenish injured tissues. However, in the particular case of the pancreas, attention should be directed primarily to the positive results that succeed to overcome the experimental obstacles. In the case of conversion of bone marrow-derived cells into endocrine lineages, the myriad of negative results appears to evolve in part because suboptimal experimental design: use of cells with partial expression of the reporter protein to detect a differentiation process that is generally associated with downregulation of the reporter protein in a tissue that physiologically silences the reporter gene.
Footnotes
Acknowledgments
We thank Dr. Saul Sharkis and Dr. Michael Collector for the outstanding support, discussion, and conceptual contribution to this study. Funding was provided by a generous grant from the Leah and Edward M. Frankel Trust for bone marrow transplantation and a generous donation in memory of Papi and Mordechai Steigman.
Author Disclosure Statement
No competing financial interests exist.