
Abstract
The potential use of neural stem cells (NSCs) in basic research, drug testing, and for the development of therapeutic strategies is dependent on their large scale in vitro amplification which, however, introduces considerable risks of genetic instability and transformation. NSCs have been derived from different sources, but the occurrence of chromosomal instability has been monitored only to a limited extent in relationship to the source of derivation, growth procedure, long-term culture, and genetic manipulation. Here we have systematically investigated the effect of these parameters on the chromosomal stability of pure populations of mouse NSCs obtained after neuralization from embryonic stem cells (ESCs) or directly from fetal or adult mouse brain. We found that the procedure of NSCs establishment is not accompanied by genetic instability and chromosomal aberration. On the contrary, we observed that a composite karyotype appears in NSCs above extensive passaging. This phenomenon is more evident in ESC- and adult sub-ventricular zone-derived NSCs and further deteriorates after genetic engineering of the cells. Fetal-derived NSCs showed the greatest euploidy state with negligible clonal structural aberrations, but persistent clonal numerical abnormalities. It was previously published that long-term passaged ESC- and adult sub-ventricular zone-derived NSCs did not show any defects in the cells' proliferative and differentiative capacity nor induced in vivo tumour formation, although we here report on the chromosomal abnormalities of these cells. Although chromosomal aberrations are known to occur less frequently in human cells, studies performed on murine stem cells provide an important complement to understand the biological events occurring in human lines.
Introduction
T
Mouse neural stem cells (NSCs) can be derived from different sources, and despite their wide use as model systems for mammalian central nervous system development and differentiation, their karyotypic stability has been poorly characterized. The few available studies have enlightened the occurrence of genomic variability in NSCs suspension cultures (ie, “neurospheres”)[19] with spontaneous transformation [20 –23]. It has been recently reported that after 4 weeks in culture, fetal brain-derived neurospheres show reduced telomere length with subsequent gain of chromosome 1 and random structural chromosomal aberrations [24]. These changes have consequences on the proliferative behavior and functions of neurosphere cells [24 –26], and it has been suggested that extensive in vitro culture might, in some studies, also generate malignant cells [27,28].
Here we focused our attention on adherently growing, self-renewing murine NSCs generated from ESCs and from primary fetal and adult central nervous system tissues [29 –31]. These cells, named NSCs, resemble neurogenic radial glia of the developing brain [32,33] and retain their multipotentiality while giving rise to up to 80% neurons after extensive in vitro expansion [29,30,33 –36]. Further, these cells maintained their capacity to generate a high percentage of MAP2 neurons with a GABAergic phenotype and firing action potentials after more than 130 passages [35]. Moreover, ES- and fetal-derived NSCs never induced any tumor formation when implanted in mice [29] (Brilli et al. unpublished data). Although these cells did not show any biologically relevant alterations, the systematic cytogenetic analysis here reported on a number of NSC lines derived from different sources revealed variable degree of chromosomal aberrations. This phenomenon did not occur during the ESCs neuralization but is a function of the derivation source, passages, and genetic modifications. Extensive chromatin fibers occasionally appeared during the neuralization process, possibly as a consequence of cell stress. NSCs started to acquire the first abnormalities at passage 5, and these became more evident after genetic engineering. Moreover, NSCs derived from fetal mouse brain were comparatively more stable (but not exempted from abnormalities) with regard to sub-ventricular zone (SVZ)-derived and ESC-derived NSCs. These results underscore the importance of performing routine cytogenetic analysis not only for studies dealing with lineage specification and maturation but also to test the potential effects of given chromosomal changes on the biochemical and physiological processes under investigation. A systematic analysis on murine stem cells is useful not only to regularly monitor their natural history but also to better understand stem cell biology.
Materials and Methods
Maintenance of mouse stem cell lines
NSCs have been derived and passaged as reported elsewhere [29,31]. Cells were routinely grown on uncoated 25 cm2 flasks (Iwaki, Barworld) in Euromed-N medium (Euroclone, Celbio) supplemented with 1% N2 (Gibco, Invitrogen) and EGF and FGF-2 (20 ng/mL each; PeproTech, Tebu-Bio) [29]. Cells were regularly passaged by Accutase (Sigma) dissociation (1–2 min at 37°C) and pelletted by centrifugation (3 min at 1.2k rpm). Medium was changed every 2–3 days.
L14 neurospheres were generated using a standard protocol [37] from E14 foetal striatum dissected from green fluorescent protein (GFP) reporter mice [38]. Neurospheres were grown in uncoated 25 cm2 flasks (Iwaki, Barworld) in the same medium used for NSCs expansion. Spheres were regularly pelletted by centrifugation (3 min at 1.2k rpm) and passaged 1:2 by mechanical dissociation every 4–5 days.
Engineered LC-1 eGFP cells were obtained through infection of LC-1 cells (passage 12) by means of lentiviral particles carrying the eGFP cDNA [29]. Bulk culture of lentivirally infected LC-1 eGFP cells is indicated as LC-1 12+# cells (# indicates the passage after the engineering procedure). eGFP labelled COR-1 cells (COR-1 eGFP) were generated by electroporation procedure at passage 6 (cultures are indicated as COR-1 6+#, where # indicates the passage after the engineering procedure).
LC-2 subclones were generated by seeding LC-2 cells (passage 3) at the density of 0.5 cell/well in a laminin coated 96-well plate. Wells containing 1 single cell were selected under the microscope and followed for colony formation. Clones were expanded and passaged as just indicated.
Cells isolated from male mice were used for the experiments in order to maintain uniformity. A detailed description of the main properties of the cells used in this work is reported in Table 1.
All cell lines maintain neurogenic stability in culture evaluated by expression of immature, neuronal and glial antigents evaluated by immunocytochemistry in self-renewal or neuronal differentiation condition.
ESCs, embryonic stem cells; NSCs, neural stem cells; NA, not available.
Chromosome preparation and staining
To obtain chromosome preparations, actively dividing cells from 70% to 80% confluent culture flasks were treated with 1 μg/mL Colcemid (Sigma) for 2 h at 37°C. The cells were then collected by centrifugation at 2,000 rpm for 6 min. A hypotonic solution consisting of 0.56% KCl w/v pre-warmed at 37°C was added to the cell pellet, prefixation with some drops of ice-cold fixative (methanol:glacial acetic acid, 3:1,v/v) was performed, and then cells were fixed with new cold fixative and stored at +4°C. About 3 drops of the cell suspension were dropped on cold, dry slides. Slides were stored at room temperature in a dry place and then stained for karyotype analysis.
Quinacrine banding
Q-bands by fluorescence and quinacrine stain (QFQ) banding was obtained by immersion of the prepared slides in a solution of quinacrine mustard (5 mg/mL; Sigma) for 1 min at room temperature, washed for 1 min, and wet-mounted in McIlvaine's Buffer under a coverslip. Metaphase chromosomes were analyzed with a Leica DMR fluorescent microscope, using a U.V. lamp, a 515 nm barrier filter, a H3 exciter filter (420–490 nm), and an immersion 100× objective.
Giemsa banding
GTG (G-banding by Trypsin and Giemsa) banding was obtained by trypsin digestion (1:250, Difco, 10% in phosphate buffer) of the prepared slides for about 2 min at room temperature followed by Giemsa stain (4% in phosphate buffer) for 12 min. Metaphase chromosomes were observed with a Leica DMR microscope set in direct-light modality.
Images from the well-spread metaphases were captured using a CCD-camera with a dedicated software (Tesi Imaging, Italy).
The karyotypes were here described using the indication of the International Committee on Standardized Genetic Nomenclature for Mice [39,40], and the definitive karyotype is given by the analysis of about at least 50 metaphases per culture passage. A chromosomal aberration was defined as clonal when at least 2 metaphases showed the same aberration. If the abnormality was a missing chromosome, the same change had to be present in at least 3 cells to be accepted as clonal. Structural aberration was defined as rearrangement of chromosome segments resulting in the formation of ring chromosome, tandem or inverted duplication, and translocation.
Results
Aneuploidy occurrence in cultured ESC-derived NSC lines
NSCs derived from different sources have been shown to retain persistent self-renewal capacity and high neurogenic competence after extensive long-term in vitro culturing [29,30,35,36]. Previous studies showed that mouse NSCs maintain diploid chromosome content on at least 10 passages in vitro [29], but no indications were available on their stability after long-term expansion and genetic engineering.
In order to investigate these aspects, we first performed a karyotypic analysis on the already available LC-1 (mESC-derived) NSCs at passage 45 (p45) and LC-1 eGFP cells (carrying the eGFP transgene) at increasing passages (p12+10; p12+18 and p12+40). We found that the percentage of euploidy in p45 LC-1 cells was very low, bringing clonal numeric and structural aberrations to 58% and 2%, respectively (Table 2 and Fig. 1a). Only one metaphase showed a Robertsonian translocation involving chromosome 5 and 16 (Rb(5.16)). These abnormalities were also further evident in the LC-1 eGFP cells, in particular at passages 12+18 and 12+40, which were completely devoid of metaphases with normal complement (Table 2 and Fig. 1a). An additional marked characteristic of LC-1 and LC-1 eGFP cells was their tendency to tetraploidy (2n ≥80). In particular, this was true at p12+10 (about 6%) and p12+40 (about 90%) for LC-1 eGFP, whereas in parental LC-1 cells, just few metaphases showed a tetraploid complement. At the different passages considered, metaphase reconstruction of LC-1 eGFP also showed, at each passage considered, a composite karyotype with different chromosomes involved generally in clonal aneuploidies that were all shared simultaneously by different metaphases. Random chromosomal loss was evident, mainly in p12+18 metaphases (33.4%). The presence of small fragments of extrachromosomal DNA was also recorded in LC-1 eGFP cells with a frequency increasing, during in vitro expansion, from 13.3% to 46.7% (Table 2), likely due to cell stress. These fragments are referred to as marker chromosome (mar) even if they showed different morphologies such as ring-shaped chromosome fragments with centromere and acentric fragments (Fig. 2).
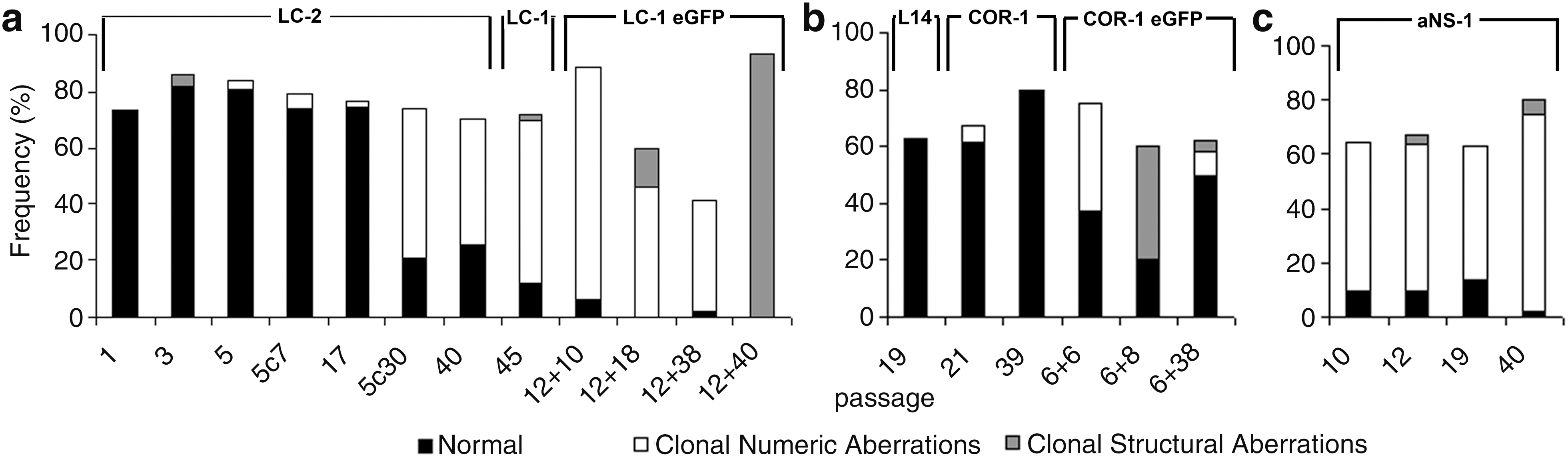
The frequency of euploidy, number of clonal/structural aberrations, and random loss/gain during the period of culture passage in:

Line LC-1 eGFP: examples of different mars (marker chromosomes; arrows) in a diploid (left) and tetraploid (right) metaphase.
Also 80,XXYY.
Not clonal; mar, marker chromosome.
Unable to define which chromosomes were involved in Robertsonian translocations; karyotype nomenclature follows the indication of the International Committee on Standardized Genetic Nomenclature for Mice (see text).
Chromosomal stability of LC-1 eGFP engineered cells was also assessed at different passages after the engineering procedure (gray rows).
On the whole, these results indicate that ES-derived NSCs lose their euploidy with long-term passaging and genetic engineering.
Analysis of the effects of neuralization process and passages on the karyotype of ES-derived NSCs
In order to systematically investigate the onset and progression of karyotipic abnormalities observed in long-term expanded and engineered LC-1 cells, we derived a new NSC line (named LC-2 cells) from mESCs. Analysis was performed on the starting ESC culture, the intermediate transient neural progenitors (NPs) [33], and the stabilized LC-2 cells at different in vitro passages before and after cryopreservation. In order to keep consistency with the previous LC-1 derivation, we employed the same ES46C mESC line [29]. Analysis of the starting ESC culture did not reveal abnormalities (Table 3), reinforcing the evidence of a broad genetic stability and integrity of the starting mESC culture. According to the NSC generation procedure [29], ES46C cells exposed for 7 days to N2B27 medium enter the neuralization phase and efficiently convert into a population of sox-1 positive NPs [41]. These represent a transient intermediate population that is transferred to EGF and FGF-2 supplemented serum-free medium (SFM) which ultimately convert them into stable, homogeneous, and long-term self-renewing NSCs [29,33]. Table 3 shows that NPs exhibited a normal chromosomal pattern with no manifest structural or numeric aberration, but a consistent fraction (about 50%) of NPs in culture exhibited the presence of extra-chromosomal chromatin fibers (Fig. 3). These structures consist of thin DNA threads that connect chromosomes [42] and resemble chromonemal fibers [43]. These fibers appear early in the NP generation phase and gradually disappear after complete NS conversion and subsequent in vitro culturing. We speculate that these fibers might be the result of a change of chromatinic status occurring during the transition from pluripotent ESCs to a somatic stem cell population or of a gradual adaptation of the cells to serum-free conditions they were exposed to during the neuralization process. When freshly generated LC-2 NSCs at p1 were analyzed, a stable chromosomal asset was found (Table 3). Nevertheless, passaging led to the appearance of minor structural (Rb19:19 at p3) and numeric aberrations (monosomy of chromosome 2 and 13 and trisomy of chromosome 1 and 12 at p5) although most of them were not clonal (Table 3). With in vitro passaging, LC-2 cells progressively showed signs of chromosomal instability, gaining clonal numeric aberrations of chromosomes 1 and/or 19 in about 50% of the metaphase analyzed (Table 3 and Fig. 1a), whereas no significative variation in random gain/loss was observed.
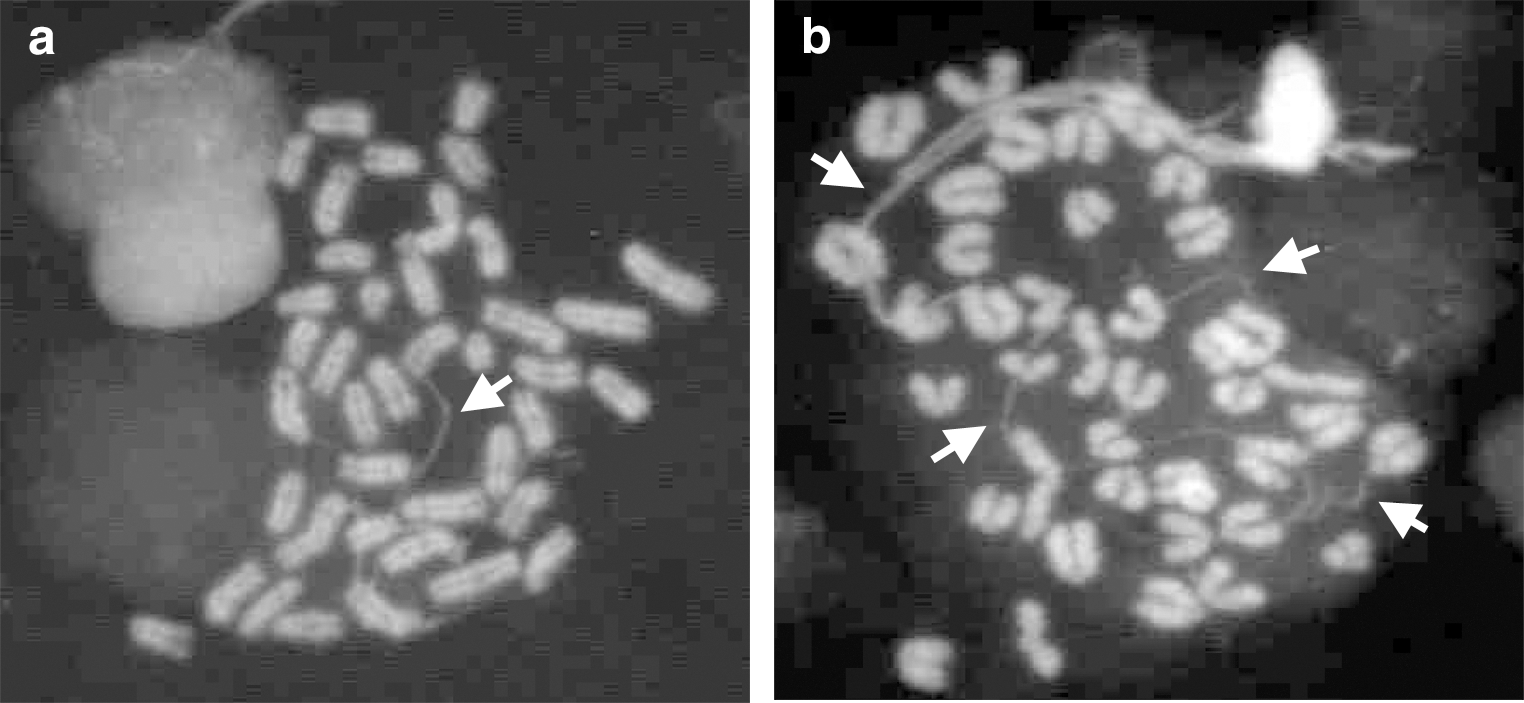
Examples of extra-chromosomal chromatin fibers (arrows) in neural progenitor (NP) cells
Not clonal; mar, marker chromosome; karyotype nomenclature follows the indication of the International Committee on Standardized Genetic Nomenclature for Mice (see text). Chromosomal stability of the LC-2 cells was also assessed after cryopreservation at passage 5 and expanded in vitro for further 7 and 30 passages (gray rows).
Karyotipic analysis of clonal populations of NSCs
In order to investigate the karyotipic stability of clonal populations, we analyzed the chromosomal pattern of different subclones generated from LC-2 (p3) bulk culture. We considered 5 different clones analyzed at p3 and p15 (Table 4). Among the 5 clones considered, 2 (LC-2_8 and LC-2_13) showed low instability and sporadic gain of chromosomes 11 and 19 and loss of 3, 13, and 14. Structural aberrations (presence of supernumerary chromosome “mar” and Robertsonian translocations) were observed only in single metaphases and they were lost with passages (Table 4). Although these events were casual and likely due to technical preparation of metaphase spreads, those occurring in LC-2_15, LC-2_20, and LC-2_22 were clonal being present in more than 3 metaphases. These cells acquired structural aberrations involving chromosomes 16 and 19 and gain of chromosomes 1, 5, and Y. These results indicate a certain heterogeneous intrinsic instability of this cell type and the tendency to acquire several aberrations that can be selectively clonally propagated in culture.
Not clonal; karyotype nomenclature follows the indication of the International Committee on Standardized Genetic Nomenclature for Mice (see text).
Analysis of the effects of cryopreservation procedure on the karyotype of ES-derived NSCs
In order to evaluate the effect of cryopreservation on possible occurrence of kariotypic instability, LC-2 cells were frozen at p5 in regular freezing medium containing 10% dimethyl sulfoxide, thawed, and cultured for another 7 and 30 passages (p5+7 and p5+30). The frozen/thawed LC-2 cells showed extra-chromosomal chromatin fibers in 12% of metaphases that were lost after 30 passages post-thawing; at this time point, the cells displayed clonal trisomy of chromosome 1 and 19 (Table 3, gray rows) as for the same cells continuously cultured without cryopreservation (Table 3, passage 40). These data indicate that when clonal aneuploidy occurs, the same chromosomes are maintained in the cells independently of the cryopreservation steps.
NSCs derived from the adult and fetal brain show different susceptibility to accumulate chromosomal aberrations with long-term in vitro culturing
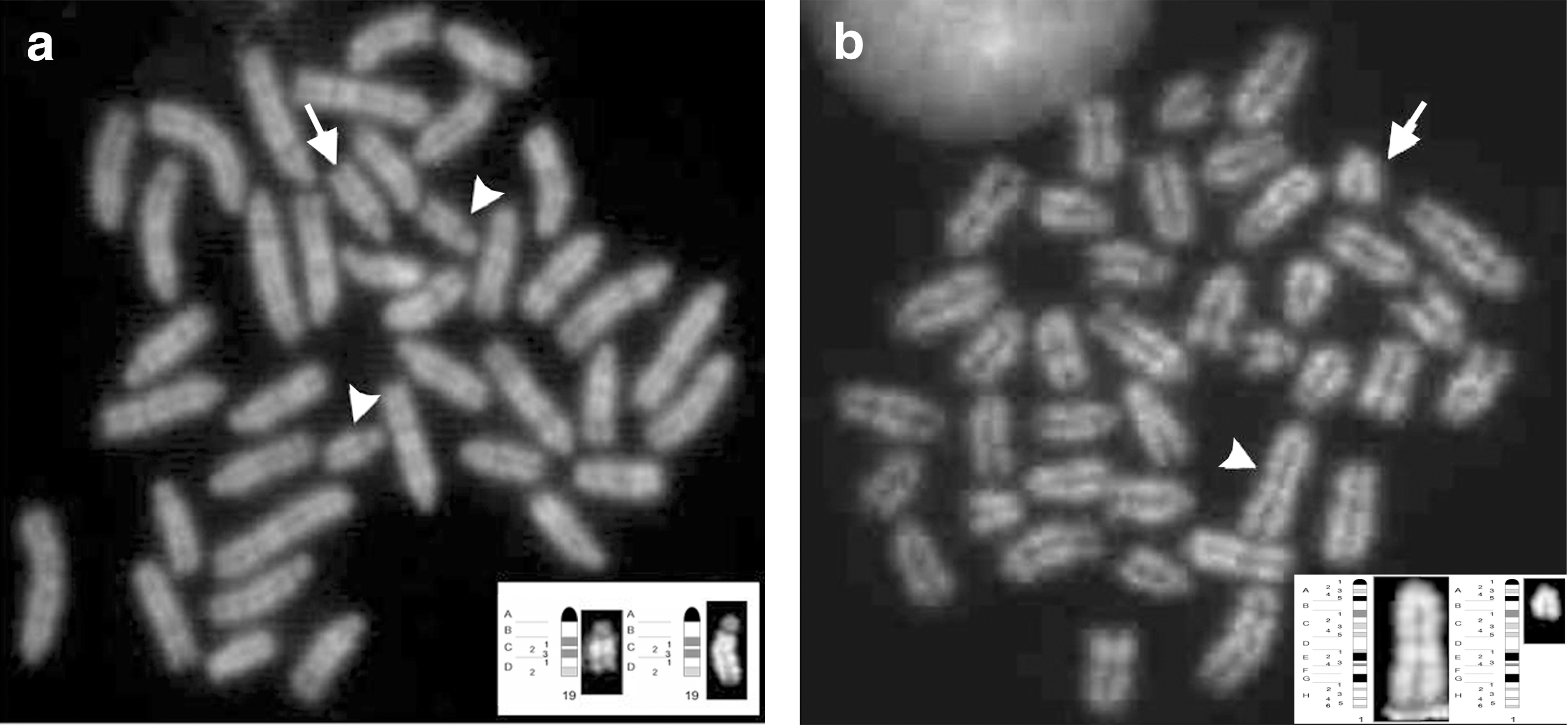
We next analyzed the chromosomal organization of NSCs previously derived from the fetal and adult mouse brain [29 –31]. In particular, adult SVZ-derived aNS-1 cells were cytogenetically examined at different passages (p10, p19, and p40), and euploid frequencies are reported in Table 5. A greater frequency of clonal aneuploidies than structural aberrations occurred in these cells (Table 5 and Fig. 1c), trisomy of chromosome 1 being the most frequent numeric aberration ranging from 29% (passage 10) to 62% (passage 40). Much less frequent were trisomies of chromosomes 18 and 19, whereas at p10 the presence of a supernumerary chromosome 19 was recorded as showing duplication of the q arm in C band (Dp(19C); Fig. 4a).
Not clonal; karyotype nomenclature follows the indication of the International Committee on Standardized Genetic Nomenclature for Mice (see text).
Chromosomal stability of the engineered cells was also assessed (gray rows).
COR-1 (fetal brain derived) NSCs were cytogenetically analyzed at passages 21 and 39. Although we observed euploidy in about 61.2% and 80% of cells at passages 21 and 39, respectively (Table 5 and Fig. 1b), there were no evident clonal structural aberrations, but only a low percentage of clonal monosomy of chromosome 17 (6.1% at passage 21). This alteration is lost with further passaging, and the random gain/loss observed in both passages was likely due to metaphase preparation.
On the whole, these results indicate that NSCs derived from fetal brain also carry chromosomal abnormalities but are less subjected to accumulate karyotipic aberrations than adult SVZ-derived NSC lines.
Monolayer of NSCs versus suspension neurosphere growth of fetal brain cells
In order to assess whether monolayer NSC culturing procedures can differently impact on chromosomal instability with regard to the suspension neurosphere culturing, we analyzed L14 neurospheres at passage 19. L14 were derived according to standardized procedures from the E14 mouse fetal brain [19]. Of the 54 metaphases analyzed in this cell line, 63% showed normal karyotype with no clonal aneuploidy (Table 5 and Fig. 1b). Random chromosome loss and gain was 24% and 13%, respectively, but this might be associated with technical artefacts (Table 5). Only 3 metaphases showed a complex karyotype: (i) 40,XY,−8,+Dp(19C) showing loss of chromosome 8 and a supernumerary chromosome 19 with a duplication of q arm in C band; (ii) 39,XXY, −9, Rb(3.5) resulting from a Robertsonian translocation between chromosome 3 and 5 (Fig. 5a); this metaphase also showed a duplication of the X chromosome and loss of chromosome 9; (iii) 40,XY,Rb(4.4) in which chromosome 4 was present in 3 copies consisting of normal 4 and a Robertsonian translocation between two chromosomes 4 linked together at the centromere (Fig. 5b).
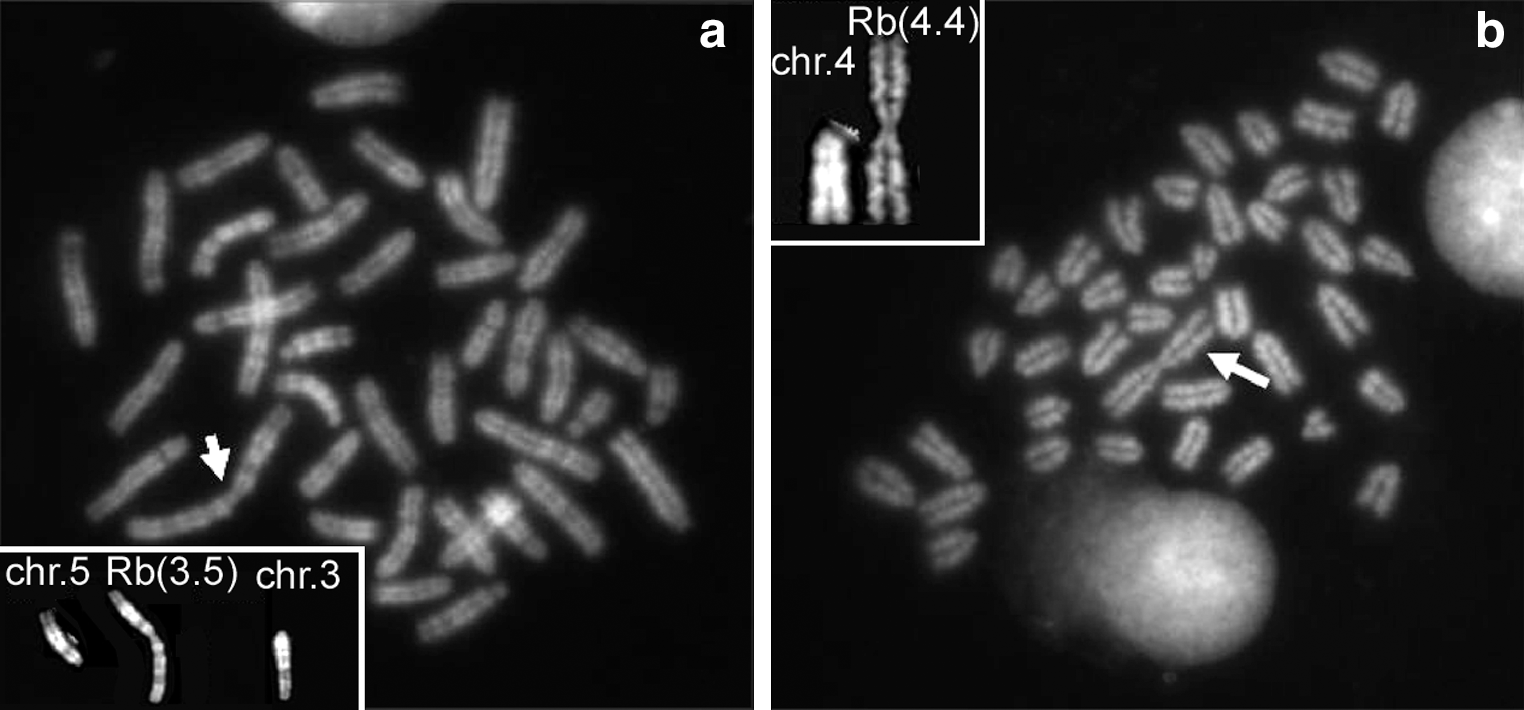
Line L14: examples of Robertsonian translocations.
On the whole, these results indicate that neurosphere cultures are comparable to monolayer of fetal NSCs in term of chromosomal instability.
The effect of genetic engineering on fetal-derived NSCs
Finally, aiming at assessing whether genetic engineering might worsen the chromosomal pattern of fetal-derived NSCs, as previously observed for ESC-derived NSCs (Table 2), we analyzed cultures of engineered COR-1 eGFP at p6+6 and p6+38 (Table 5, gray rows). Although the mitotic index for these cells was low (few metaphases per passage) except for passage p6+38, euploidy ranged from 50% at p6+38 to 37.5% at p6+6, suggesting that only cells with a normal complement succeed in completing cell division and are positively selected. In these cells, clonal aberrations both numeric and structural were reported (Table 5 and Fig. 1b). Notably, we observed deletion of chromosome 1 from C band of q arm to Tel1q (del(1C)) with a frequency of 4% in p6+38 (Fig. 4b). Clonal monosomy for chromosomes 13 and 19 were observed at passage 6+6 with a frequency of 37.5% and of 8% at passage 6+38, respectively (Table 5).
These results indicate that engineering procedures applied to the established mouse NS lines exacerbate the accumulation of the chromosomal defects independently of the source of origin of the NSCs.
Discussion
Several evidences have described the onset of chromosomal abnormalities in cells grown in vitro [20,44,45]. This has also been reported to occur occasionally during mESC derivation and culturing [46]. Similar abnormalities have been found in neurons and glia in the normal brain, likely due to mitotic segregation defects occurring in neural progenitor cells [47]. In particular, a number of studies now indicate that within the adult human and mouse SVZ and other brain regions, there are a considerable number of neural stem/progenitor cells showing loss of heterozygosity and an abnormal karyotype resulting in chromosomal mosaicism and cellular diversity [48]. Structural variation in the neural genome is likely to be one important mechanism for neuronal diversity, as functioning neurons with aneuploid genomes form genetically mosaic neural circuitries as part of the normal organization of the mammalian brain [49]. Accordingly, these chromosomal changes may be brought in vitro [20,47,50]. The presence of chromosomal abnormalities in cultured neural stem/progenitor cells may, thus, reflect the brain area from which the cells originate [21,51] in addition to the way in which NSCs are isolated or processed, their long-term cultivation [52] and, possibly, cryopreservation conditions.
Here we performed a systematic cytogenetic analysis of mouse adult, fetal, and ES-derived NSC lines at different culturing passages summarized in Fig. 1 and Table 6. We found various chromosomal abnormalities including polyploidy, mono and trisomy, translocations and duplications in all the cell lines considered. As previously reported, chromosomes gain (either total or partial acquisition) predominate over loss in all cell lines analyzed as a function of culturing passages [15]. In general, cells in culture acquire DNA damage and likely undergo random mutations at a rate of about 1 nucleotide per 109 cells [2]. These phenomena may confer gradual adaptation to the in vitro synthetic environment. Besides the numerous and diverse nature of the alterations, we have observed the presence of chromatin fibers in a number of metaphases that represent unfolding of entire or partial chromosomes. Extensive chromatin fiber conformation influences DNA tension [53], linker histone H1 binding to NAP1 [54], and in vivo and in vitro DNA methylation-dependent chromatin fiber compaction [55]. It has been observed that during the development of transient organs in animals and plants, sister chromatids usually do not become separated after replication rounds and retain cohesion at highly reiterated sites as centromeres. This might trigger the formation of hyperploid cells, a condition widely observed in stem cell lines and malignant tumors that arises initially from cells arrested in the G2/M or spindle checkpoint, unable to segregate chromosomes by mitosis [56]. We mainly detected the presence of these fibers in some cells during the neuralization phase, when cultures are exposed to serum-free media and, therefore, subjected to considerable stress adaptation. Their presence gradually decreased as cells proceeded toward the NSC stage differentiation, whereas numerical and structural chromosomal aberrations increased with the number of passages. Serum starvation may represent a source of chromosomal instability, as it can induce structural aberrations and aneuploidy [57]. It has been reported that human fetal fibroblasts exposed to serum deprivation display double centrosomeres, translocation, deletions, and high telomerase activity, which, in turn, might cause cellular transformation. It may be that the de-condensation of the chromatin, either totally or partially, may set the stage for further chromosomal breakage and dysfunctions. Further studies are required in order to elucidate the exact biological role of these structures in cultured cells.
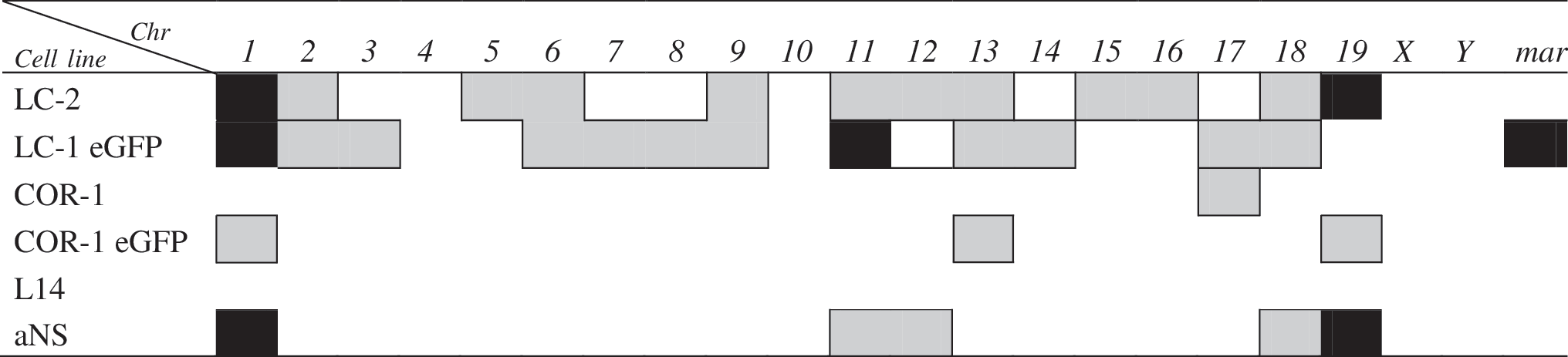
In black, the clonal chromosomal aberrations (either numeric or structural) mantained in further passages; in gray, clonal chromosomal aberrations observed only at a specific passage.
However, ESCs subjected to serum starvation for 7 days during neuralization procedure did not show any chromosomal aberration, which were instead appearing only after NSC lines were established. The most affected were the ESC derived LC-1 and LC-1 eGFP cells, the latter showing tetraploidy (2n>80) in about 90% of the metaphases (passage12+40). LC-1 eGFP cells (derived from LC-1 cell line on lentiviral infection carrying the cDNA for the GFP protein) exhibited further severe clonal numeric and structural aberrations with regard to the parental LC-1 cells. Insertion of foreign genetic material via retroviral vectors is known to trigger genomic instability, especially when the number and sites of insertion are arbitrary [58 –60]. Nevertheless, this technique is normally used to trace cells after transplantation to follow migration/integration in the host tissue and lineage specifications [61].
In the literature, there are divergent data on chromosomal stability of non-immortalized NSC lines and some data indicate that, at least for adult mouse SVZ-derived NSCs, chromosomal abnormalities are actually expected. Morshead and colleagues [27] reported that neurospheres after 15–20 passages undergo transformation as indicated by decreased doubling time, loss of growth factor dependence, or differentiative potential. Similar observations have been recently reported for mouse fetal-derived neurospheres [24]. On the contrary, one other study [62] reported that non-clonal neurosphere cultures derived from mouse adult (2 month old) SVZ are karyotypically normal until passage 40 (when they generate ∼15% of ß3-tubulin immunoreactive cells) but later at passage 70 begin to exhibit chromosomal aberrations in 10% of cells although no variation(s) on the behavior was reported. In our hands, fetal-derived neurosphere culture did not show any clonal aberrations at passage 19 although sporadic alteration of chromosomes 3, 5, and 19 were found as also described by other authors [24]. Also, no significant differences were reported when fetal NSCs were cultured in adherence, suggesting that none of these culture methods protects NSCs from gaining clonal aberrations.
We conclude that NSCs from different sources can manifest a variable tendency to acquire karyotypic aberrations, especially after extensive in vitro expansion. Fetal NSCs seem to acquire less chromosomal anomalies in culture. This might be due to programmed cell death that naturally occurs in vivo in more than 20% of fetal brain cells coincidental with early developmental processes of proliferation, migration, and differentiation [63]. The clearance of aneuploid cells in the developing brain could be one of the main functions of programmed cell death [48]. On the contrary, recent studies have hypothesized that mitotic-spindle checkpoint, which helps maintain chromosomal integrity during cell division, functions in hESCs and mESCs but does not initiate apoptosis as it does in somatic cells [64]. This allows an unusual tolerance to polyploidy resulting from failed mitosis. Thus, the spindle checkpoint is “uncoupled” from apoptosis in stem cells and is a likely source of karyotypic abnormalities. Importantly, in spite of the chromosomal anomalies observed in ES-, fetal- and adult-derived NSCs, we never observed important modifications in the biological behaviour of NSCs, also after prolonged growth in culture, that might be indicators of cellular transformation [29,30,35]. Aneuploidy and chromosomal aberrations also frequently occur in cultured cells in the absence of any predisposition to transformation [49,62]. Similarly, the chromosomal abnormalities observed in the cell lines herein tested did not lead to any alteration in their differentiation capacities as previously demonstrated [30,35,36], and none of the NSC lines tested in this manuscript has ever led to tumor formation when implanted in mice, even after more than 70 in vitro passages [29] (Brilli et al., unpublished data).
Conclusions
Given the huge potential of NSCs as model systems to study molecular events in normal and pathological processes, it is important to monitor genomic changes in cultured cells and to eventually determine the impact of such changes. Rapid loss of euploidy and accumulation of chromosome aberrations as a function of passage are probably inherent to most of the in vitro expansion procedures, at least for mouse cells, and may be even part of a physiological process in vivo.
Our results indicate that chromosomal aberrations accumulate in NSCs with passages in vitro in conjunction with loss of euploidy. It is still unclear what triggers the numerous and diverse chromosome abnormalities observed in NSCs, especially the ones derived from ESCs and adult SVZ. This may be related to the culturing conditions, the methods of dissociation, and the percentage of O2.
Our results suggest that mouse NSCs in culture have a predisposition to genetic instability although they do not apparently exhibit visible variations in their growth and differentiation properties. These results also open to the need to define new culture conditions to decrease chromosomal alterations in long-term cultures and should encourage researchers to perform regular high-resolution molecular and cytogenetic studies to monitor the chromosome integrity of in vitro expanded stem cell cultures.
Footnotes
Acknowledgments
Research activities are supported by NEUROscreen consortium (European Community's Sixth Framework Programme grant agreement LSHB-CT-2007-037766) to P.D. and L.C. and by NeuroStemcell (European Community's Seventh Framework Programme grant agreement nr. HEALTH-2008-B-222943) to I.B. and E.C.
Author Disclosure Statement
No competing financial interests exist.