
Abstract
It is well known that terminally differentiated cells derived from mesenchymal stem cells (MSCs) will lose the immunomodulation capacity. NANOG is known to be a core transcription factor in the maintenance of stem cell specific features or stemness. To evaluate whether NANOG was involved in the immunomodulation effects of MSCs, MSCs' immunomodulation capacity on lymphocyte activation and proliferation before or after endogenous NANOG interference was investigated. We found that MSCs' inhibitory effects on lymphocyte activation and proliferation was significantly weakened after NANOG knockdown. In addition, NANOG RNAi and chromatin immunoprecipitation experiments showed that NANOG suppressed the expression and secretion of DKK-1, transforming growth factor-beta1 (TGF-β1), TGF-β2, and TGF-β3, which are all important factors mediating MSCs' immunomodulation capacity. Based on these data, we propose that NANOG plays an important role in maintaining the immunomodulation functions of MSCs by regulating the expression and secretion of TGF-β1, TGF-β2, TGF-β3, and DKK-1.
Introduction
M
Previously, we have identified a pluripotent MSC subpopulation, which has the potential to differentiate into endothelium, epithelium, hematopoietic cells, muscle cells, and so on [20 –24], as well as immunomodulatory capacity [6,25 –27]. We found that these MSCs express NANOG [28], which is in consistent to the results of studies by Greco et al. [29] and La Rocca et al. [30]. Based on our previous finding that NANOG, as a critical transcriptional factor, was involved in regulating Dkk-1 expression by MSCs to inhibit cancer cell proliferation [28], we hypothesized that NANOG may play a role when MSCs excert their immunosuppressive effects. Our results showed that MSCs' immunomodulatory ability was significantly weakened when NANOG expression was knocked down via iRNA interference, which was accompanied by decreased expression of Dkk-1 and TGF-β. Our research provides the important evidence for the first time that there is an association between the NANOG and immunomodulatory characteristic of MSCs.
Methods
Culture of adipose-derived MSCs
Human raw lipoaspirates from 6 female patients undergoing selective suction-assisted lipectomy were collected as previously described after obtaining informed consent from the patients according to procedures approved by the Ethics Committee at Chinese Academy of Medical Sciences and Peking Union Medical College [22]. The patients were 52, 32, 31, 29, 30, and 32 years old, respectively. Briefly, the raw liposuctioned aspirates were digested with 0.2% collagenase II (Sigma) at 37°C for 30 min and cells were plated in T-25 tissue culture dishes at a density of 2×106 cells/mL. The culture medium used was DF12 containing 40% MCDB medium (Sigma), 2% fetal calf serum (Gibco Life Technologies), 1× insulin transferrin selenium (Gibco Life Technologies), 10−9 M dexamethasone (Sigma), 10−4 M ascorbic acid 2-phosphate (Sigma), 10 ng/mL epidermal growth factor (R&D Systems), 10 ng/mL platelet-derived growth factor BB (R&D Systems), 100 U/mL penicillin, and 1,000 U/mL streptomycin (Gibco). Once adherent cells were >70% confluent, they were detached with 0.125% trypsin and 0.01% EDTA and replated at a 1:3 dilution under the same culture conditions.
Vector construction
MSCV-neo-pSilencer, an RNA interference vector based on MSCV-neo (Clontech), was modified by inserting an siRNA expression cassette into the MCS and deleting 3′ LTR enhancer region (U3). MSCV-neo-simock (RNAi nonsilencing control), MSCV-neo-siDKK1 (targeting DKK-1), MSCV-neo-siNANOG (targeting NANOG), MSCV-neo-siTGF-β1, MSCV-neo-siTGF-β2, and MSCV-neo-siTGF-β3 were constructed by inserting specific shRNA templates into the siRNA expression cassette, respectively. The shRNA templates specific to their target genes were designed, synthesized, and annealed. The sequences of each shRNA are shown in Table 1.
Retrovirus preparation and transduction
Retroviral production and transduction of MSCs were essentially carried out as previously described [31]. Briefly, MSCV-neo-simock, MSCV-neo-siDKK1, MSCV-neo-siNANOG, MSCV-neo-siTGF-β1, MSCV-neo-siTGF-β2, and MSCV-neo-siTGF-β3 vectors were cotransfected with pGag-pol and pVSVG to 293T cells using Lipofectamine® 2000 (Invitrogen). At 48–72 h posttransfection, the supernatants were collected, concentrated, and added to the culture media of MSCs. The transducted MSCs were used in the following experiments after 48 h of culture.
RNA preparation and reverse transcription–polymerase chain reaction
Total RNAs were extracted using Trizol® reagent (Invitrogen), and genomic DNA from all samples was removed using the RNA-free DNase digestion kit (Takara) according to the manufacturer's protocol. One microgram of total RNA was reverse transcribed using the Reverse Transcriptase M-MLV (Takara). Polymerase chain reactions (PCRs) were carried out using Taq DNA polymerase (Takara) and the primer sequences used to identify gene expression are listed in Table 2. Real-time PCR was performed with an ABI Prism Sequence Detection System 7500 (Applied Biosystems) using QuantiTectTM SYBR® Green PCR Kit (Qiagen). Triplicate wells were averaged and GAPDH was used as an internal control to normalize for loading differences between samples.
Western blotting
Cells was lysed in ice-cold lysis buffer [1% Triton X-100, 20 mM Tris-HCl (pH 8.0), 137 mM NaCl, 10% glycerol (v/v), 2 mM EDTA, 1 mM phenylmethylsufonyl fluoride, 10 μg/mL aprotinin, 10 μg/mL leupeptin, 50 μg/mL trypsin inhibitor, 1 mM sodium orthovanadate]. Cell lysates were then clarified by centrifugation and aliquots of each were removed for protein determination by the BCATM Protein Assay Kit (Pierce). Standard western blotting procedure was used with the polyclonal anti-NANOG (Santa Cruz), polyclonal anti-DKK1 (Santa Cruz), polyclonal anti-TGF-β1, 2, 3 (R&D Systems), and anti-mouse/anti-rabbit/anti-goat horseradish peroxidase-conjugated secondary antibodies (Zhongshan Biotech Com). Antibody/antigen complexes were detected using ECL reagent (Millipore).
Effect of MSCs on lymphocyte cycle
MSCs were prepared as previously described [26]. Mononuclear cells were separated by a Ficoll-Paque gradient centrifugation (specific gravity: 1.077 g/mL); these cells were washed with D-Hanks for 2 times and used as lymphocyte. Lymphocytes were cultured alone or cocultured with either NANOG, DKK-1, TGF-β1, TGF-β2, or TGF-β3–silenced MSCs, respectively, in RPMI 1640 supplemented with 10% fetal bovine serum, stimulated with phytohemagglutinin (PHA) (50 μg/mL, final concentration) for 2 days, and then harvested and quantified. One million lymphocytes were fixed with 70% cold ethanol at 4°C for 30 min, washed with phosphate-buffered saline twice, and stained with 50 μg/mL propidium iodide (PI) (Sigma) at room temperature for 5 min. Lymphocyte cycle was analyzed with ModFIT software.
Effect of MSCs on lymphocyte activation
Lymphocytes were cultured alone or cocultured with either NANOG, DKK-1, TGF-β1, TGF-β2, or TGF-β3–silenced MSCs, respectively, and stimulated with PHA (50 μg/mL final concentration) as described earlier. The expression of CD25 (BD) and CD69 (BD) was evaluated by flow cytometry at 24 h (CD25) or 12 h (CD69).
Chromatin immunoprecipitation assay
We performed chromatin immunoprecipitation (ChIP) as previously described [32]. Antibodies used in this experiment were anti-NANOG goat polyclonal antibody (Santa Cruze) and normal goat IgG (Santa Cruze). The DNA isolated from the complex was subject to PCR amplification using primers flanking the NANOG binding site [33] in the DKK-1 gene promoter: 5′-GGA GGA GGG CAA CTG AAG GAC-3′ (forward) and 5′-CCT CTC CTT TAT GCC AAT ACT CGC-3′ (reverse). The PCR products were run on a 2% agarose gel and stained with ethidium bromide.
Statistics
Statistical analysis was performed with the statistical SPSS 13.0 software. The paired-sample t-test was used to test the probability of significant differences between samples. Statistical significance was defined as P<0.05.
Results
NANOG is involved in immunomodulatory effects of MSCs
MSCs suppressed the activation and proliferation of T lymphocytes in vitro. To figure out whether NANOG played a role in this process, we downregulated NANOG with siRNA interference. After siRNA transfection, NANOG expression was decreased by nearly 80%, as shown by reverse transcription polymerase chain reaction (RT-PCR) and western blot (Fig. 1A, B). We then compared the immunomodulatory capacity of MSCs carrying NANOG siRNA (SiN) or MSCs with mock vectors (Mock). When lymphocytes were cultured alone (None group), the percentage of CD69+ cells were 42.24%±4.5% at 12 h after activation and the percentage of CD25+ cells was 31.21%±9.63% at 24 h after activation, respectively. When lymphocytes were cocultured with Mock MSCs (Mock group), the percentages of CD69+ and CD25+ cells decreased to 23.66%±9.4% and 9.25%±5.8%, respectively (P<0.05, vs. None group). However, when lymphocytes were cocultured with SiN-MSCs (SiN group), percentages of CD69+ and CD25+ cells restored to 31.77%±10.27% and 18.4%±6.27%, respectively (P<0.05, vs. Mock group) (Fig. 2A–D). We next examined the cell cycle status of activated lymphocytes under similar conditions. When lymphocytes were cultured alone, 72.27%±6.29% cells were in G0/G1. In Mock-MSCs group, the G0/G1 rate increased to 90.62%±7.76% (P<0.05, vs. None group). In SiN-MSCs group, the G0/G1 rate restored to 79.79%±6.89% (P<0.05, vs. Mock group) (Fig. 2E, F). These results showed that suppression of NANOG expression significantly reduced MSCs' ability to either suppress lymphocyte activation or arrest lymphocytes in G0/G1 phase.

RNAi efficiency of NANOG in mesenchymal stem cells (MSCs). RNAi efficiency of NANOG in MSCs was assayed by
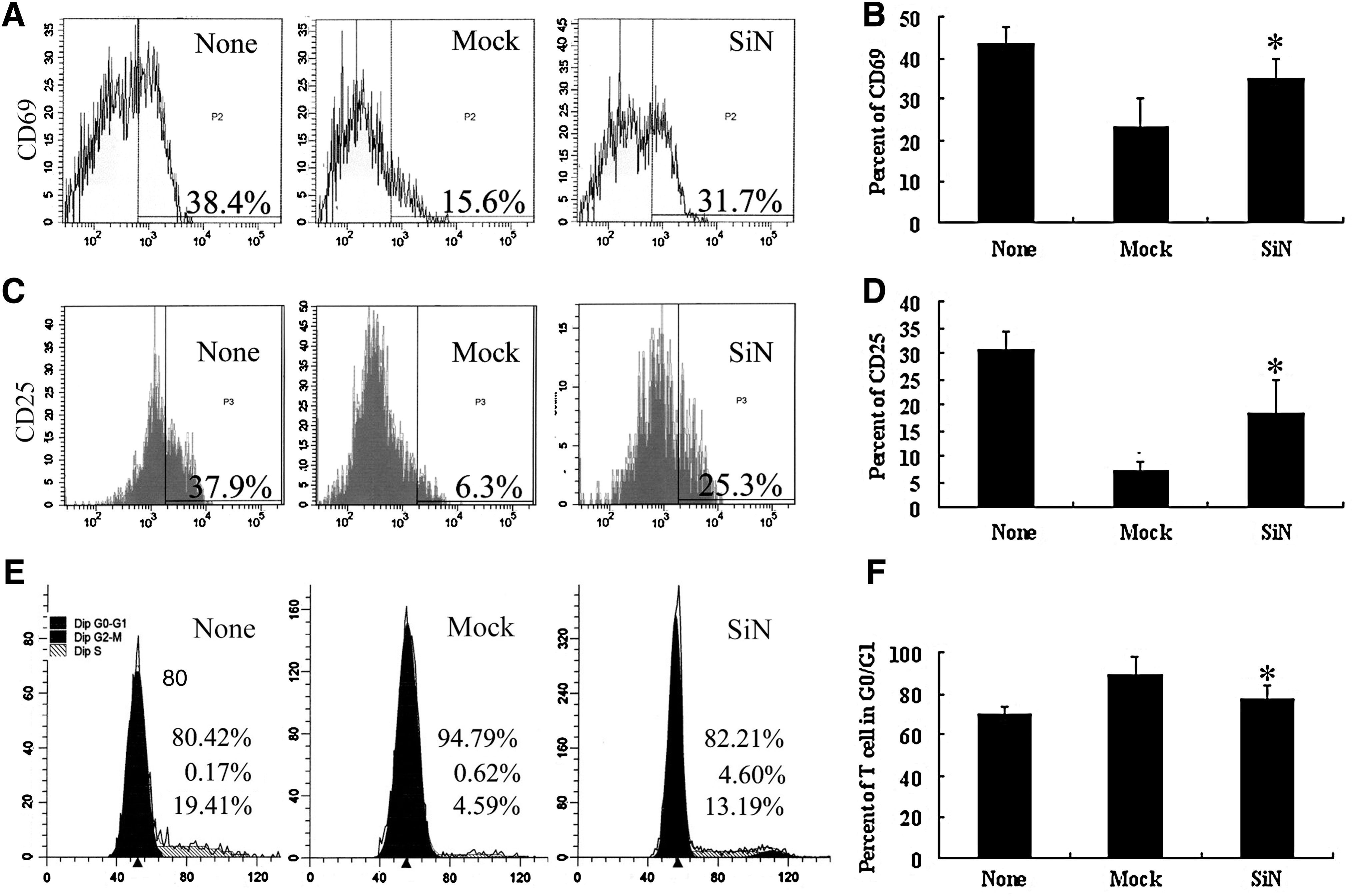
Expression of Nanog in MSCs influences lymphocyte proliferation and activation. The inhibitory effect of MSCs on lymphocyte proliferation and activation is attenuated when Nanog in MSCs is knocked down by RNAi. CD69
NANOG knockdown decreases secretion of TGF-β1, 2, and 3 in MSCs
As a nuclear transcriptional factor, NANOG maintains the characteristic features of stem cells by regulating expression of a variety of target genes. Previous studies have reported that MSCs exert their immunomodulatory function partly by secreting a lot of immunosuppressive factors. We next investigated whether NANOG could regulate the expression of these factors. We focused on TGF-β1, TGF-β2, TGF-β3, DKK-1, HGF, and IL-10 as the possible candidates of NANOG-specific target genes and detected the expression levels of these genes after NANOG knockdown.
We performed RT-PCR to measure the expression levels of TGF-β1, β2, β3, DKK-1, HGF, and IL-10 in SiN-MSCs and Mock-MSCs (Fig. 3A). The mRNA expression levels of TGF-β1, 2, and 3 and DKK-1 decreased significantly, whereas IL-10 expression was upregulated in SiN-MSCs. Expression of HGF was not affected. These results were confirmed by western blot (Fig. 3B).

Expression of DKK-1 and transforming growth factor-beta1 (TGF-β1), 2, and 3 in MSCs is regulated by NANOG. Dkk-1, TGF-β1, TGF-β2, TGF-β3, hepatocyte growth factor (HGF), and interleukin-10 (IL-10) expression levels in MSCs with Nanog knockdown (SiN) were assayed by real-time PCR
To investigate the mechanism how NANOG regulates the expression of these factors, we scanned upstream promoter region of DKK-1 gene and found NANOG-specific binding sequence. CHIP assay confirmed that NANOG could bind the upstream promoter region of DKK-1 gene (Fig. 4). Unexpectedly, we did not find any NANOG binding sequence in the upstream promoter region of either TGF-β1, β2, or β3, indicating that NANOG might indirectly regulate TGF-β1, β2, and β3.
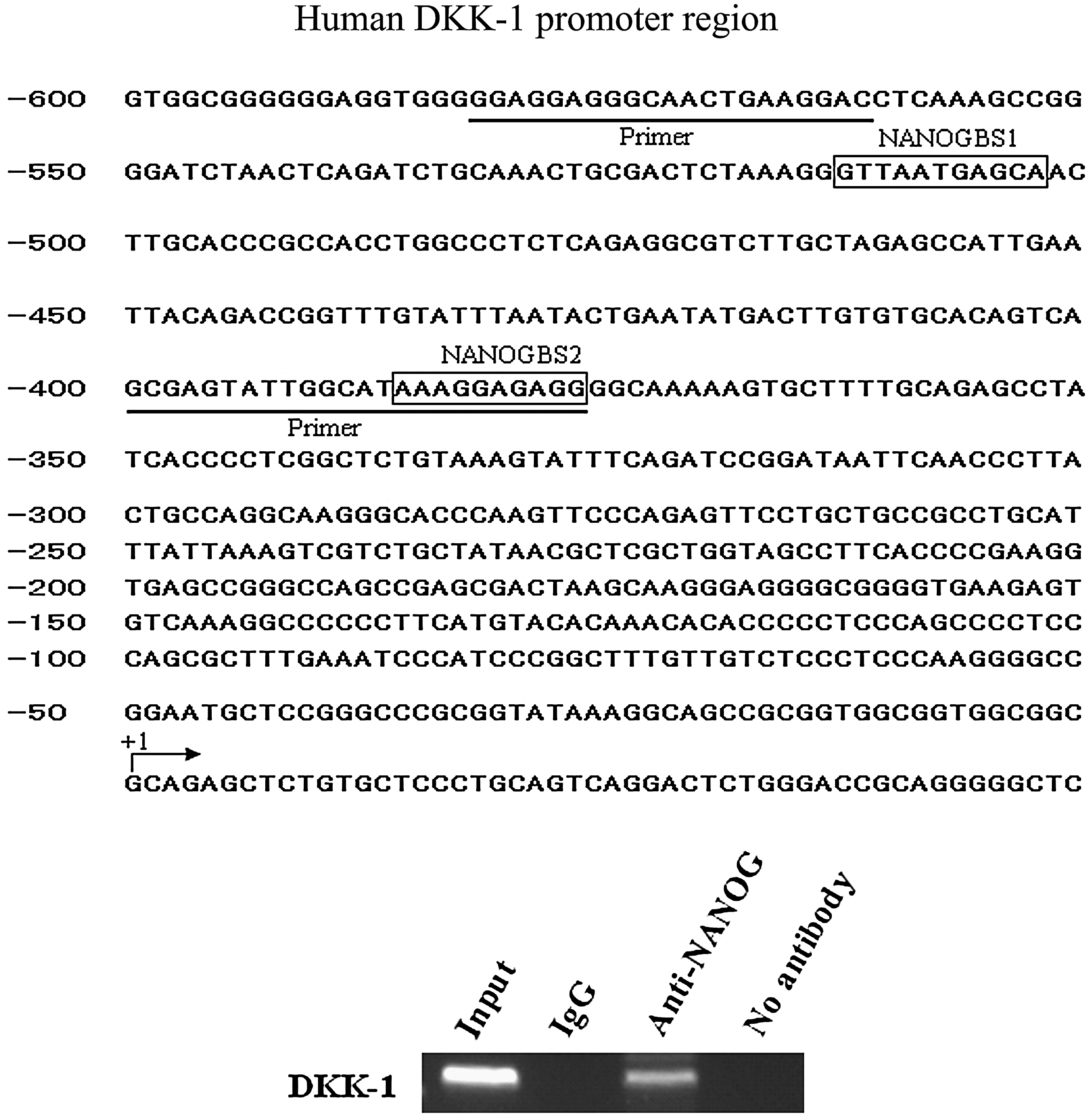
NANOG binding upstream promoter region of DKK-1 gene. The primers were designed to amplify a 206-bp fragment (−582/−377) of DKK1 promoter region spanning these 2 NANOG binding sites in chromatin immunoprecipitation samples. Background signals were tested amplicons in negative control samples immunoprecipitated without antibody.
Mechanism of immunomodulation of MSCs
To demonstrate the exact role of TGF-β subtypes and DKK-1 secreted by MSCs in lymphocyte activation and proliferation suppression, we knocked down the expression of DKK-1, TGF-β1, TGF-β2, and TGF-β3 in MSCs, respectively, by iRNA and then evaluated their ability on suppressing lymphocyte activation. RT-PCR and western bolt showed that the expression of these cytokines decreased by 70% after siRNA transfection (Fig. 5A, B). The percentage of CD69+ activated lymphocytes was 42.24%±4.5% (None group), which decreased to 23.66%±9.4% when cocultured with Mock-MSCs (vs. None group, P<0.05). The CD69 expressing rate restored to 34.5%±6.11% when lymphocytes were cocultured with MSCs transfected with TGF-β3 interfering vectors (vs. Mock-group, P<0.05), but not with MSCs transfected with DKK-1, TGF-β1, or TGF-β2 interfering vectors (Fig. 6A).

RNAi efficiency of DKK-1, TGF-β1, TGF-β2, and TGF-β3 in MSCs. RNAi efficiency of TGF-β1, TGF-β2, TGF-β3, and Dkk-1 in MSCs was assayed by real-time PCR
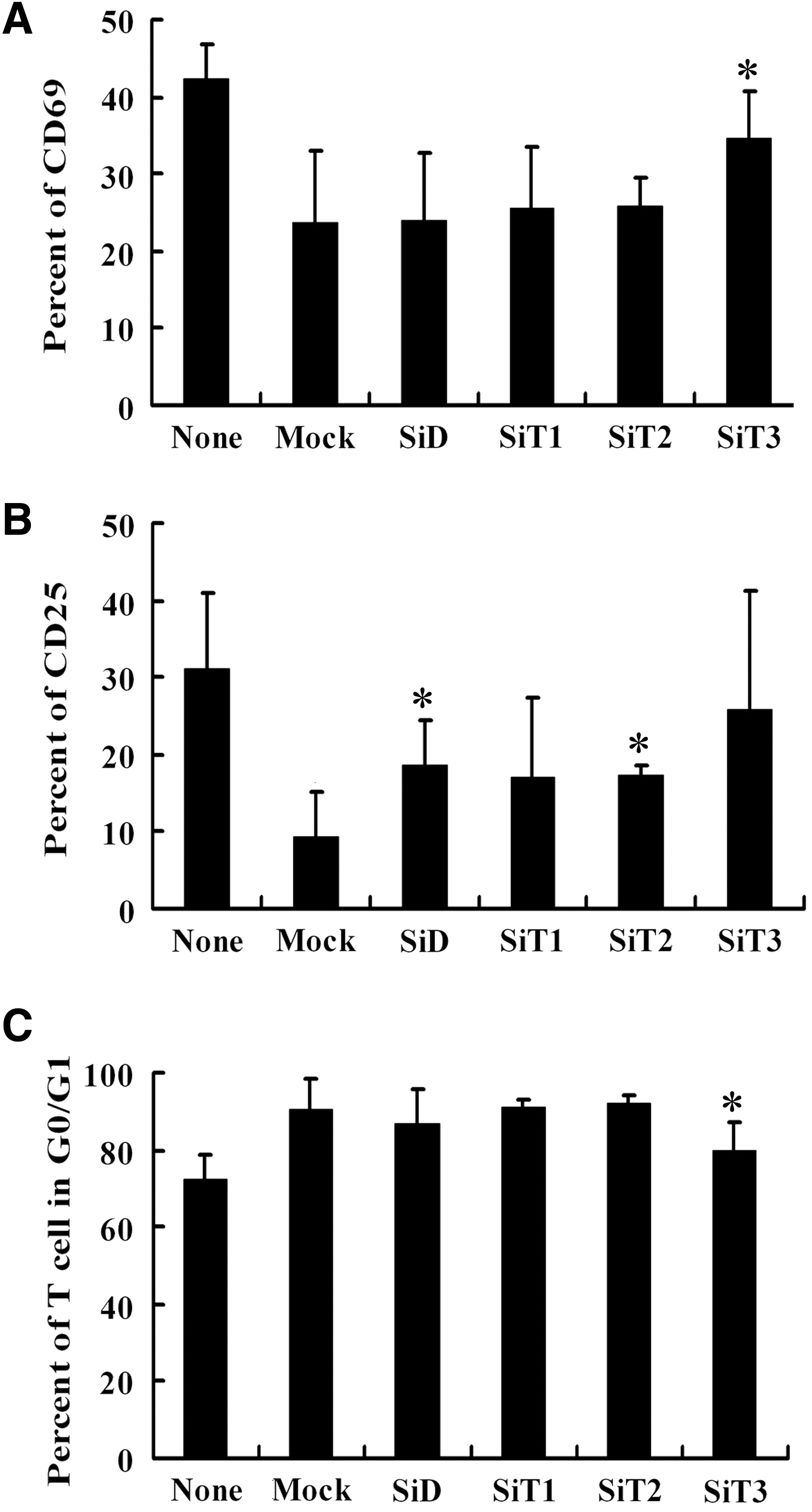
The inhibitory effect of MSCs with silenced DKK-1, TGF-β1, TGF-β2, and TGF-β3 on lymphocyte proliferation and activation. The inhibitory effects of MSCs with knocked-down DKK-1, TGF-β1, TGF-β2, and TGF-β3 on lymphocyte cycle and activation were compared with that of MSCs carrying Mock:
Twenty-four hours after PHA stimulation, the expression rate of CD25 was 31.21%±9.63% when lymphocytes were cultured alone (None group). It decreased to 9.25%±5.8% when lymphocytes were cocultured with Mock-MSCs (vs. None group, P<0.05), but restored to 18.5%±6.0% and 17.17%±1.5% when DKK-1 and TGF-β2 were knocked down by iRAN, respectively (P<0.05, vs. Mock group). CD25 expressing rate of lymphocytes cocultured with TGF-β1 and TGF-β3 interfered MSCs were 16.9%±10.57% (SiT1 group) and 25.7%±15.5% (SiT3 group), respectively, which have no significant difference with MSCs carrying Mock group (Fig. 6B).
Cell cycle analysis showed that 72.27%±6.29% of lymphocytes were in G0/G1 phase after PHA stimulation in the absence of MSCs (None group). When they were cocultured with Mock-MSCs (Mock group), the rate of G0/G1 phase increased to 90.62%±7.76% (P<0.05, vs. None group). It decreased to 79.72%±7.52% when TGF-β3 was knocked down (P<0.05, vs. Mock group). But knockdown of DKK-1, TGF-β1, or TGF-β2 did not reduce MSCs' ability to arrest lymphocyte to G0/G1 cycle (Fig. 6C). These results showed that TGF-β and DKK-1 cooperated in MSCs' modulatory effect.
Discussion
MSCs possess immunomodulatory capacity both in vitro and in vivo, which depicts a broad prospect for treating disorders of immune system. Unlike conventional immunosuppressants, MSCs are living cells, which secrete a variety of cytokines to regulate the interactive network of immune reactions. The underlying mechanisms are, therefore, fairly complicated and still largely unknown, although lots of investigations have been done.
NANOG is a transcriptional factor in charge of stem cell renewal and maintaining multipotency, first discovered in embryonic stem cells (ESCs) and later in cord blood stem cells and MSCs [29,34 –36]. So far as we know, NANOG is specifically expressed in stem cells and shares little homogeneity with other genes. Currently, there is no investigation addressing the role of NANOG in the immunomodulatory ability of MSCs. In this experiment, we, for the first time, demonstrated that NANOG was involved in MSCs' ability to suppress lymphocyte activation and proliferation. We found that suppression of Nanog by iRNA significantly reduced MSCs' ability to suppress lymphocyte cycle and activation. As PHA is a T-cell mitogen, it can be hypothesized that T cells are a predominant target of the immunosuppressive effect of MSCs in our culture system. NANOG is a nuclear protein rather than a secreted soluble factor and thereby cannot influence lymphocyte directly. We proposed that NANOG regulated the expression of other specific target genes to realize those functions. In the present study, we focused on the relation between NANOG and soluble mediators. We found that both RNA and protein expression levels of TGF-β1, TGF-β2, TGF-β3, and DKK-1 decrease in MSCs when NANOG was knocked down by iRNA. Further study revealed that there is a NANOG binding domain in the upstream promoter region of DKK-1, which was confirmed by CHIP assay. However, no such NANOG binding domain could be identified within the upstream promoter region of either TGF-β1, β2, or β3. Presently, how NANOG regulates TGF-β1, β2, and β3 expression is unclear. It has been reported that Nanog can interact with Smad1 in mouse ESCs, interfere with the recruitment of coactivators to active Smad transcriptional complexes, and repress the expression of bone morphogenetic protein-responsive genes [37]. Xu et al. further reported that SMADs can bind the NANOG promoter and directly regulate NANOG expression [38].
TGF-β was found to act as a potent growth inhibitor of a variety of cell lines [39]. TGF-β has 3 isoforms (type β1, type β2, and type β3) with 70%–80% sequence homology, which use the same receptors and exhibit indistinguishable effects on immune cells in vitro. TGF-β1 is produced by many cell types and TGF-β3 is mainly produced by mesenchyma [40,41]. We knocked down different subtypes of TGF-β in MSCs and showed that TGF-β3 was the key molecule responsible for the suppressive effects of MSCs on lymphocyte activation and proliferation. This is in consistent with our previous observation that MSCs from myelodysplastic syndrome-refractory anemia (MDS-RA) patients secreted less TGF-β1 and β3 and had deficient immunomodulatory function [26].
DKK-1 is a blocker of Wnt signaling pathway [42,43] and is involved in the antiproliferation effect of MSCs [44]. We recently also demonstrated that MSC inhibits tumor cell proliferation by DKK-1 [28]. However, whether DKK-1 was also involved in MSCs' inhibition of lymphocyte proliferation and activation has not been reported. In this study, the results from RNA interference showed that DKK-1 played a synergetic role in the immunomodulation of MSCs on lymphocyte activation, but had no influence on lymphocyte cycle arrest. Although it has been reported that DKK-1 played a role in suppressing tumor growth [45 –47] indicating that the mechanism of MSCs' immunomodulation is very complicated.
Nevertheless, these soluble factors were not the only means by which MSCs realized the effects of suppressing lymphocyte cycle and activation. Previous studies demonstrated that dozens of soluble factors were involved in the immunomodulation of MSCs. We believe that various cytokines work together to carry out the immunomodulation. The mechanism of immunomodulation of MSCs in vivo would be more complicated, as MSCs are living cells, which can react differently to various microenvironments of immune.
Footnotes
Acknowledgments
This work was supported by grants from the “863 Projects” of Ministry of Science and Technology of People's Republic of China (no. 2006AA02A109, 2006AA02A115), National Natural Science Foundation of China (no. 30830052), Beijing Ministry of Science and Technology (no. D07050701350701), and Cheung Kong Scholars Program.
Author Disclosure Statement
No competing financial interests exist.