
Abstract
The development of nonhuman primate (NHP) embryonic stem cell (ESC) models holds great promise for cell-mediated treatment of debilitating diseases and to address numerous unanswered questions regarding the therapeutic efficacy of ESCs while supplanting ethical considerations involved with human studies. Here we report successful establishment and characterization of 3 novel baboon (Papio cynocephalus) ESC lines from the inner cell mass of intracytoplasmic sperm injection-derived blastocysts. Embryos were cultured in an improved baboon embryo in vitro culture protocol. The inner cell mass of blastocyst was laser-dissected and plated on mouse embryonic fibroblast feeder cell monolayer in the NHP ESC culture medium. Three cell lines with characteristic ESC morphology have been cultured through an extended period (>14 months), with 2 male cell lines (UT-1 and −2) and 1 female cell line (UT-3) displaying normal baboon karyotypes. Reverse transcription-polymerase chain reaction analysis confirmed that all 3 lines express primate ESC pluripotency markers, including OCT-4, NANOG, SOX-2, TERT, TDGF, LEFTYA, and REX-1. All 3 lines demonstrated positive immunocytochemical staining for OCT-4, stage-specific embryonic antigen-3, stage-specific embryonic antigen-4, TRA-1-60, and TRA-1-81. Baboon ESCs injected into NOD/SCID mice formed teratomas with all 3 germ layers. In addition, embryoid body-like spherical structures were derived and initial outgrowth was observed when embedded into extracellular matrix Matrigel®. The ESC lines established in this NHP model have the potential to extend our knowledge in the fields of developmental biology, regenerative medicine, and future applications, including preclinical safety assessment of in vivo stem cell therapy.
Introduction
E
We selected yellow baboon (P. cynocephalus), an Old World NHP species, to derive ESCs for scientific and practical reasons. The baboon is a close relative to humans and the complex physiology of humans can be mimicked in this species [21], hence the importance as a relevant experimental model for studying human medicine. In addition, baboons are readily available from the large colony at Southwest Foundation of Biomedical Research in San Antonio, Texas. Historically, baboon in vitro fertilization (IVF) and embryo culture have poor yields of embryos reaching the blastocyst stage when compared to rhesus monkey and humans; the majority baboon preimplantation embryos stall at morula stage. We have tested multiple media and feeder cell combinations to optimize culture conditions with improved blastocyst rates [22,23]. On the basis of the improved embryo culture protocol, our objectives in this study were to derive baboon ESC from the ICM of intracytoplasmic sperm injection (ICSI)-derived baboon embryos, and characterize such baboon ESC lines. Successful derivation and characterization of 3 baboon ESC lines after laser-assisted dissection of the ICM are described in the present study.
Materials and Methods
Baboon ovarian stimulation, oocyte retrieval, IVF with ICSI, and embryo culture
Animal protocols conducted in this study were approved by the Institutional Animal Care and Use Committee at the University of Texas Health Science Center at San Antonio. Individually caged adult healthy fertile female baboons at day 1 or 2 postmenses received recombinant human gonadotropin r-FSH (75 IU/day; Serono) for 7–8 days and r-FSH/r-LH (75 IU each/day; Serono) for 3 days followed by a single injection of human chorionic gonadotropin (hCG, 2,500 IU; Serono), followed by laparoscopic surgery and follicular aspiration 35–37 h after hCG injection. Follicular fluid aspirates were examined under a stereomicroscope (Olympus) to identify cumulus-oocyte complexes, oocytes were denuded, and cumulus-free mature oocyte morphology was accessed. Those at metaphase II stage with the presence of first polar body were selected for IVF-ICSI. Male baboons were sedated with ketamine and semen specimens were collected by rectal probe electro-ejaculation. Semen was processed by swim-up, and sperm with normal morphology and motility were selected for ICSI. A single sperm was immobilized by gently squashing the sperm tail against the bottom of the Petri dish before ICSI. Injected oocytes were rinsed twice in the mHTF HEPES-buffered medium (Irvine Scientific), and then transferred into 4-well plates (Nunc) of the embryo culture medium (Sydney IVF; Cook Medical) and buffalo rat liver (BRL; ATCC) feeder cell coculture, supplemented with 10% fetal bovine serum and amino acids, covered by light paraffin oil, and placed in incubator with 37°C triple gas (5% CO2, 5% O2, 90% N2).
Fertilization was assessed 16–18 h post ICSI, with the normal fertilization determined by the observation of 2 pronuclei and protrusion of the second polar body. Zygotes with normal fertilization were selected and returned to the plate with the culture medium and feeder cells. Starting at 48 h post ICSI, embryos were transferred into a new plate of the blastocyst culture medium (Sydney IVF; Cook Medical) with BRL cell coculture every 2 days through the blastocyst stage in incubator at 37°C supplemented with triple gas (5% CO2, 5% O2, 90% N2).
Laser-assisted dissection of ICM and derivation of baboon ESC
Blastocysts from combinations of sequential and single media with or without BRL cell coculture were dissected to isolate ICM from trophectoderm (TE) using Zilos-tk laser module (Hamilton Thorne). Isolated ICM were plated on 4-well plates, precoated with a monolayer of mitomycin-C inactivated mouse embryonic fibroblast (MEF; Millipore) feeder cells, in the conditioned ESC culture medium consisting of knockout Dulbecco's modified Eagle's medium (Invitrogen) supplemented with 20% knockout serum replacement (Invitrogen), 2 mM
After 10–14 days in 4-well plates, cell colonies with ESC morphology were mechanically passaged to 6-well plates (Corning) precoated with MEF and cultured in the MEF-conditioned ESC culture medium changed daily. The culture medium used in this study was conditioned with MEF in T75 flasks (Corning) for 24 h before the medium change, and bFGF and LIF were added before the medium change. Areas of differentiated morphology were removed and only those with ESC morphology were passaged to mitomycin-C-inactivated MEF in 6-well plates 10–14 days after the initial plating, and then subsequent large cell colonies were mechanically split and transferred onto new feeder layer for propagation every 5–8 days depending on the confluence of ESC and condition of MEF. Under daily monitoring, differentiated non-ESC morphology areas found during the cell colony expansion were mechanically trimmed by 25 gauge needles to facilitate the continuous growth of undifferentiated ESC colonies. Cells were cultured in incubator at 37°C with triple gas.
Characterization of baboon ESC by karyotyping
Baboon ESC karyotyping was performed by on actively dividing ESC treated with ethidium bromide, Colcemid, Hank's balanced salt solution, and Carnoy's fixative (3:1 methanol: acetic acid) in separate steps. Cell suspensions were placed on slides for G-Banding by 2.5% trypsin and Wright stain and viewed to identify metaphase cells. Suitable metaphase cells were captured and karyotyped, with 10 cells counted and 2 analyzed.
Real-time reverse transcription-polymerase chain reaction analysis with pluripotency marker gene primers
RNA was isolated from baboon embryo-derived cell colonies with ESC morphology and cultured using RNeasy Micro kit (Qiagen) with a genomic DNA removal step as per manufacturer's protocol. Reverse transcription (RT) was carried out using the Ready-To-Go T-Primed First-Strand Kit (GE Healthcare). Baboon fetal skin fibroblast cells and RNA-free water were used as controls. Real-time RT-polymerase chain reaction (PCR) and subsequent analyses were carried out using Smartmix PCR beads (Cepheid) with 0.25 μL SybrGreen in the Cepheid SmartCycler using primer sets for common NHP ESC pluripotency markers OCT-4 (POU5F1), NANOG, SOX-2, LEFTYA (LEFTY2), TDGF, TERT [24], REX-1 [25], and housekeeping gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH) transcripts [26]: OCT-4 (POU5F1) forward TGGAGAAGGAGAAGCTGGAGCAAA, reverse GAAGCTAAGCTGCAGAGCCTCAAA; NANOG forward CAGAACTGTGTTCTCTTCCACCCA, reverse CGCTGATTAGGCTCCAACCATACT; SOX-2 forward CCCCCGGCGGCAATAGCA, reverse TCGGCGCCGGGGAGATACAT; LEFTYA (LEFTY2) forward CCTGTGTCCTTCCATTTCCTGTCT, reverse AAGCCCTTCATCCTTCCTCTTAGC; TDGF forward CATATTGGCCAGTCTGGTCTCGAA, reverse ATTCCTTCAGCACTCTGGTTCCTC; TERT forward AGCTATGCCCGGACCTCCAT, reverse GCCTGCAGCAGGAGGATCTT; REX-1 forward GCGTACGCAAATTAAAGTCCAGA, reverse primer CAGCATCCTAAACAGCTCGCAGAAT; GAPDH forward ACCACAGTCCATGCCATCAC, reverse TCCACCACC CTGTTGCTGTA.
The thermal cycling conditions included an initial denaturation step of 2 min at 95°C, and 40 cycles of 10 s/95°C–15 s/60°C–30 s/72°C. For quantification, the cycle threshold number (Ct), which exhibits the maximum curve growth rate, was determined by using Cepheid SmartCycler software version 3.0. The relative gene expression of each sample, which was normalized with respect to GAPDH, was calculated by using the formula: 2Ct (GAPDH) – Ct(gene). Melt curve analysis was performed after each real-time PCR to ascertain PCR product specificity. Real-time PCR assays were performed in duplicates. PCR amplified products were electrophoresed in 1.2% agarose and stained with 0.5 μg/mL ethidium bromide and photographed with the Bio-Rad image system using Quantity One 4.4.0 software (Bio-Rad Laboratories).
Immunocytochemical staining with pluripotency markers
Expression of NHP ESC markers OCT-4, stage-specific embryonic antigen (SSEA)-3, SSEA-4, TRA-1-60, and TRA-1-81 was demonstrated by fluorescent immunocytochemical (ICC) staining, with 4′,6-diamidino-2-phenylindole (D9542; Sigma) used as the control. Cells were fixed with 4% paraformaldehyde in phosphate-buffered saline for 20 min at room temperature for OCT-4, TRA-1-60, and TRA-1-81, or absolute methanol 10 min at −20°C for SSEA-3 and SSEA-4. For OCT-4 staining, cells were permeablized in 0.2% Triton X-100 and 0.1% Tween-20 at room temperature for 40 min. Cells were blocked with 0.05% Tween-20 with 2% normal goat serum or 2% normal donkey serum followed by incubation with primary antibodies for 40 min at room temperature and secondary antibodies for 40 min at room temperature in the dark. All primary antibodies were from Santa Cruz Biotechnology, and all secondary antibodies from Jackson Immuno Research Labs. The dilution used was 1:100 for all except 1:50 for OCT-4 primary antibody: OCT-4, mouse monoclonal IgG (SC-5279)/Cy3 conjugated goat anti-mouse IgG (115-165-062); SSEA-3, rat monoclonal IgM (SC-21703)/Cy3 conjugated goat anti-rat IgM (112-165-075); SSEA-4, mouse monoclonal IgG (SC-21704)/Cy3 conjugated goat anti-mouse IgG (115-165-062); TRA-1-60, mouse monoclonal IgM (SC-21705)/Cy3 conjugated donkey anti-mouse IgM (715-165-140); TRA-1-81, mouse monoclonal IgM (SC-21706)/Cy3 conjugated donkey anti-mouse IgM (715-165-140).
Baboon ESC-derived teratoma formation
Baboon ESCs were excised from colonies and centrifuged. Pellets containing ∼1–1.5 × 105 (∼15–20 μL) cells were loaded into a 1 mL syringe attached to a 30 gauge needle and injected into the kidney capsule, testis, or intramuscular (i.m.) into the hind legs of individual nonobese diabetic severe combined immunodeficiency (NOD/SCID; Harlan) mice at 7–8 weeks of age. Teratoma formation was assessed by regular palpation starting 8–10 weeks postinjection, and dissection was performed on weeks 12–17 when a skin bulge outside of teratoma injection sites was obviously visible. SCID mice were euthanized and teratomas were dissected, processed with 10% buffered formalin, and then embedded in paraffin blocks to examine in vivo differentiation of ESC. Paraffin-embedded teratoma sections were stained with hematoxylin and eosin (H&E). H&E-stained slides were examined using an inverted microscope at 100× magnification.
In vitro differentiation of embryoid body formation
ESC colonies were mechanically excised from the culture plate and plated on low-adhesive 6-well plates (Corning). Spherical formations of embryoid bodies (EB) were embedded in 6-well plates precoated with extracellular matrix (Matrigel®; BD-Biosciences). ESC-derived EB formation and potential outgrowth in Matrigel was continuously monitored.
Results
Establishment of baboon ESC lines
Five out of 11 ICMs isolated from blastocysts derived from 4 female baboon oocyte retrievals, ICSI, and embryo culture successfully attached on MEF within 24 h (Fig. 1). ICMs from expanded blastocysts showed a higher rate of attachment to MEF than nonexpanded blastocysts (4 out of 7, and 1 out of 4, respectively). One cell clump detached from the MEF feeder layer after a week, whereas the other 4 cell remained attached and started proliferation into cell colonies with distinct ESC-morphology, including flat colonies with well-defined borders, aggregated colonies of cobblestone-like cells with high nucleus-to-cytoplasm ratio, and prominent nucleoli identifiable under an inverted microscope.
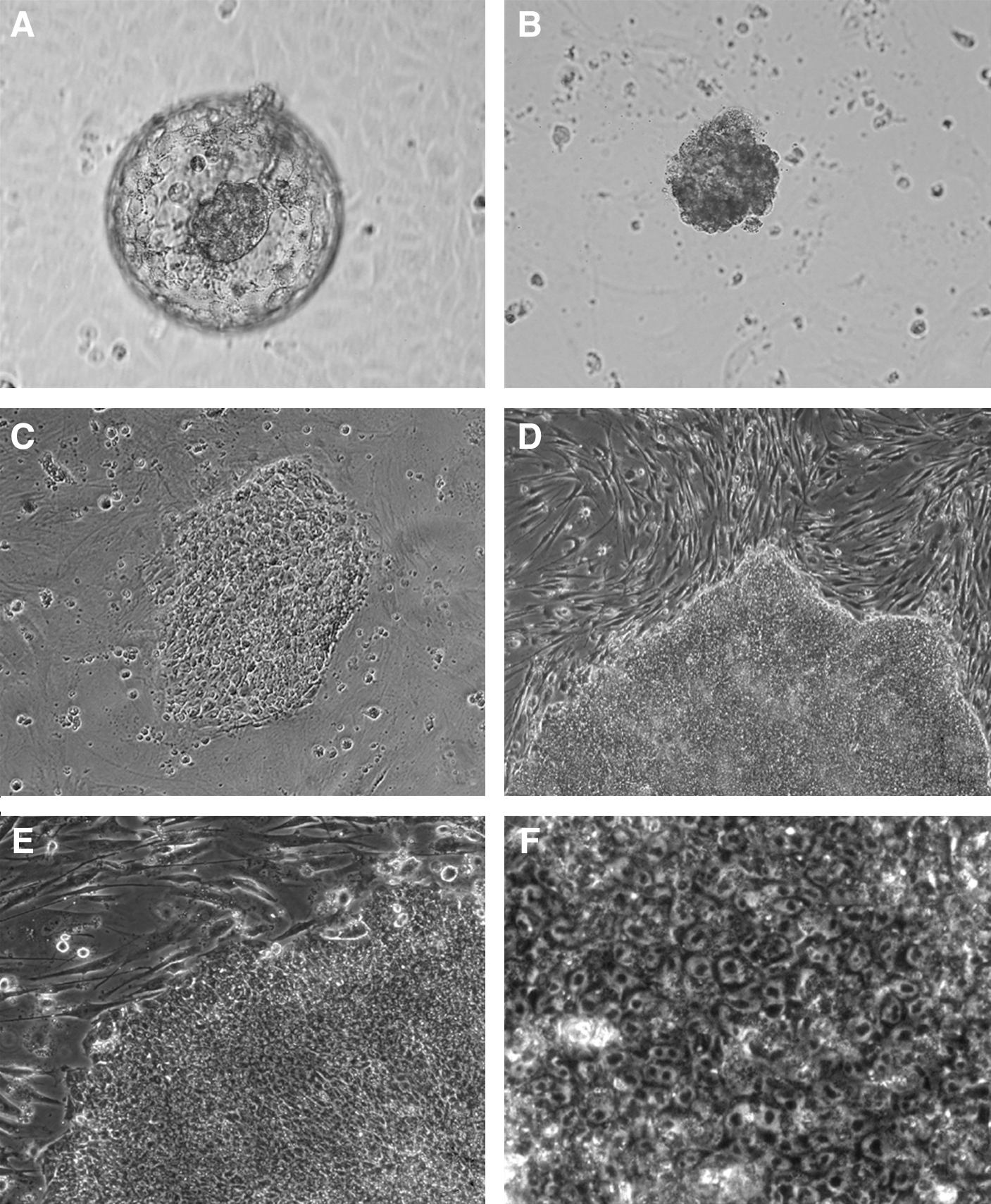
Representative micrographs of baboon embryonic stem cell (ESC) derivation: expanded baboon in vitro-cultured blastocyst
Among the 4 cell lines that successfully attached and expanded on MEF, 3 were stably passaged for an extended period (UT-1: 482 days, 65 passages; UT-2 and −3: 439 days, 55 passages) with morphology similar to human and other primate ESC lines. The fourth line was lost a month after the initial ICM plating due to areas showing a high portion of differentiation and unsuccessful subcultural of undifferentiated areas.
In this study a lower concentration 4 ng/mL bFGF, commonly used for human ESC culture, was also tested and we found high percentage of differentiation in all 3 ESC lines and consequent low survival rate postpassaging, compared to the effect of higher concentration 15 ng/mL bFGF. This may indicate that the requirement of bFGF in the culture medium to maintain undifferentiated status of baboon ESC is higher than it is in human and some other NHP ESC protocols.
In separate attempts to derive ESC from morula-stage baboon embryos, 31 pronase-treated zona-free morula-stage baboon embryos were plated on MEF feeders, and among those, 4 showed increase in blastomere number. Cell colony expansion with resemblance of ESC-like morphology was found in 1 of those 4 embryos after a week. The only morula-derived cell colony growth discontinued after 2 weeks post MEF plating. Poor results were also found in attempts of plating 15 baboon 2–8 cells stage embryos, with no blastomere remained attached on MEF after a week.
Karyotyping
Baboon ESC lines displayed normal baboon karyotypes of 20 autosomes and 2 sex chromosomes, including 2 male (UT-1 and −2: 42, XY) and 1 female (UT-3: 42, XX) cell lines (Fig. 2A–C).
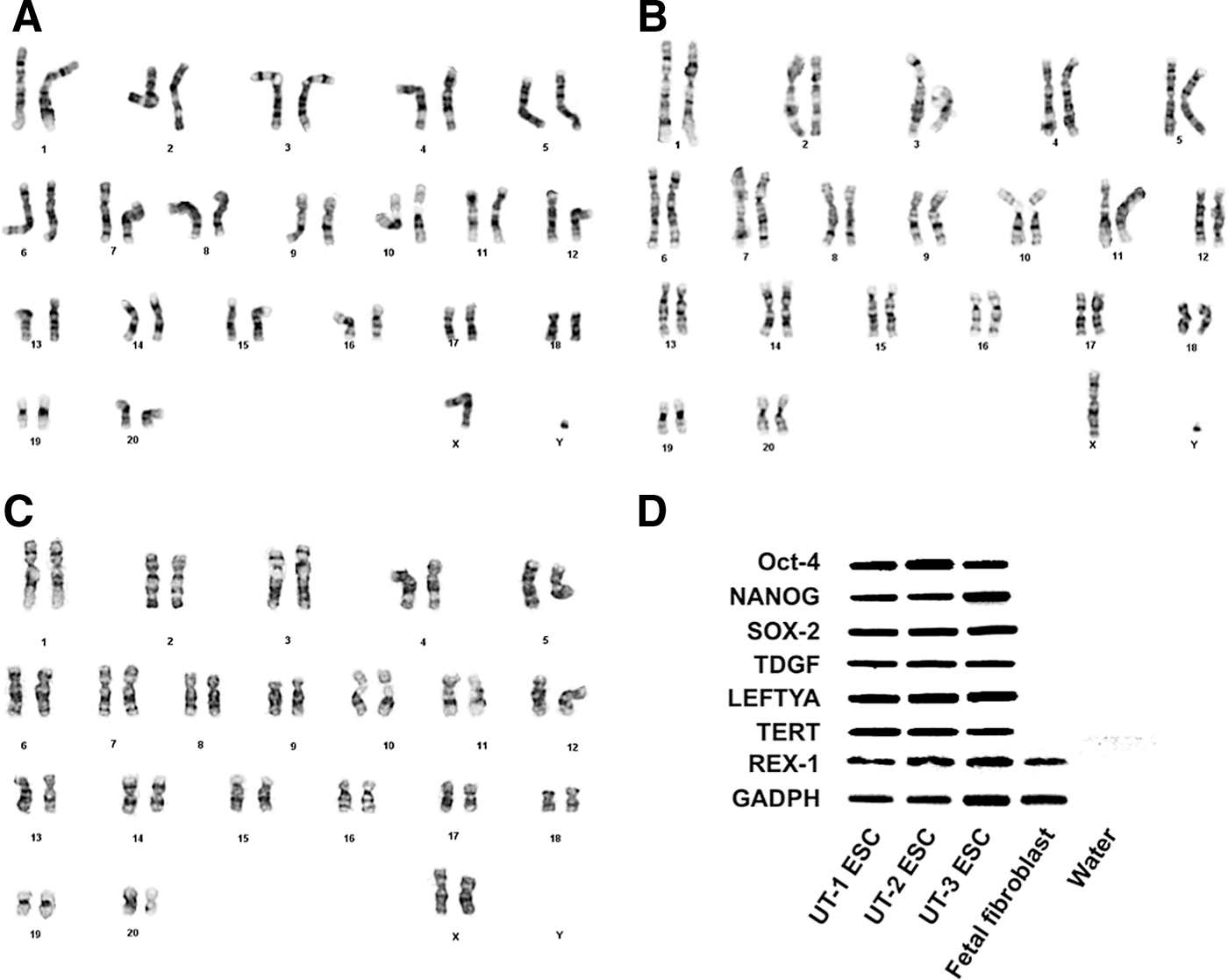
Baboon ESC lines displayed normal baboon karyotypes of 2 male ESC lines (UT-1 and −2: 42, XY) and 1 female ESC line (UT-3: 42, XX)
Real-time RT-PCR analysis
Positive real-time RT-PCR amplification by primer sets for common NHP ESC pluripotency markers OCT-4 (POU5F1), NANOG, SOX-2, LEFTYA (LEFTY2), TDGF, TERT, REX-1, and housekeeping gene GAPDH were found in all 3 baboon ESC lines. PCR amplified products were electrophoresed and photographed (Fig. 2D). No positive amplification was found in the control group of baboon second trimester fetal fibroblasts except for REX-1, and water showed negative with all markers.
ICC staining with pluripotency markers
Fluorescent ICC staining on 3 baboon ESC lines showed positive expression of OCT-4, SSEA-3, SSEA-4, TRA-1-60, and TRA-1-81 (Fig. 3). The control staining of 4′,6-diamidino-2-phenylindole showed positive staining on nuclei of all cell types, including the MEF feeder. The staining with OCT-4, TRA-1-60, and TRA-1-81 were more consistent than the mixed intensity of expression with cell surface glycophingolipids SSEA-3 and −4. Steady expression of SSEA-4 was found in all 3 ESC lines, although not as strong as OCT-4, TRA-1-60, and TRA-1-81. The expression intensity of SSEA-3 immunostaining is slightly lower in UT-2 and UT-3 than UT-1.
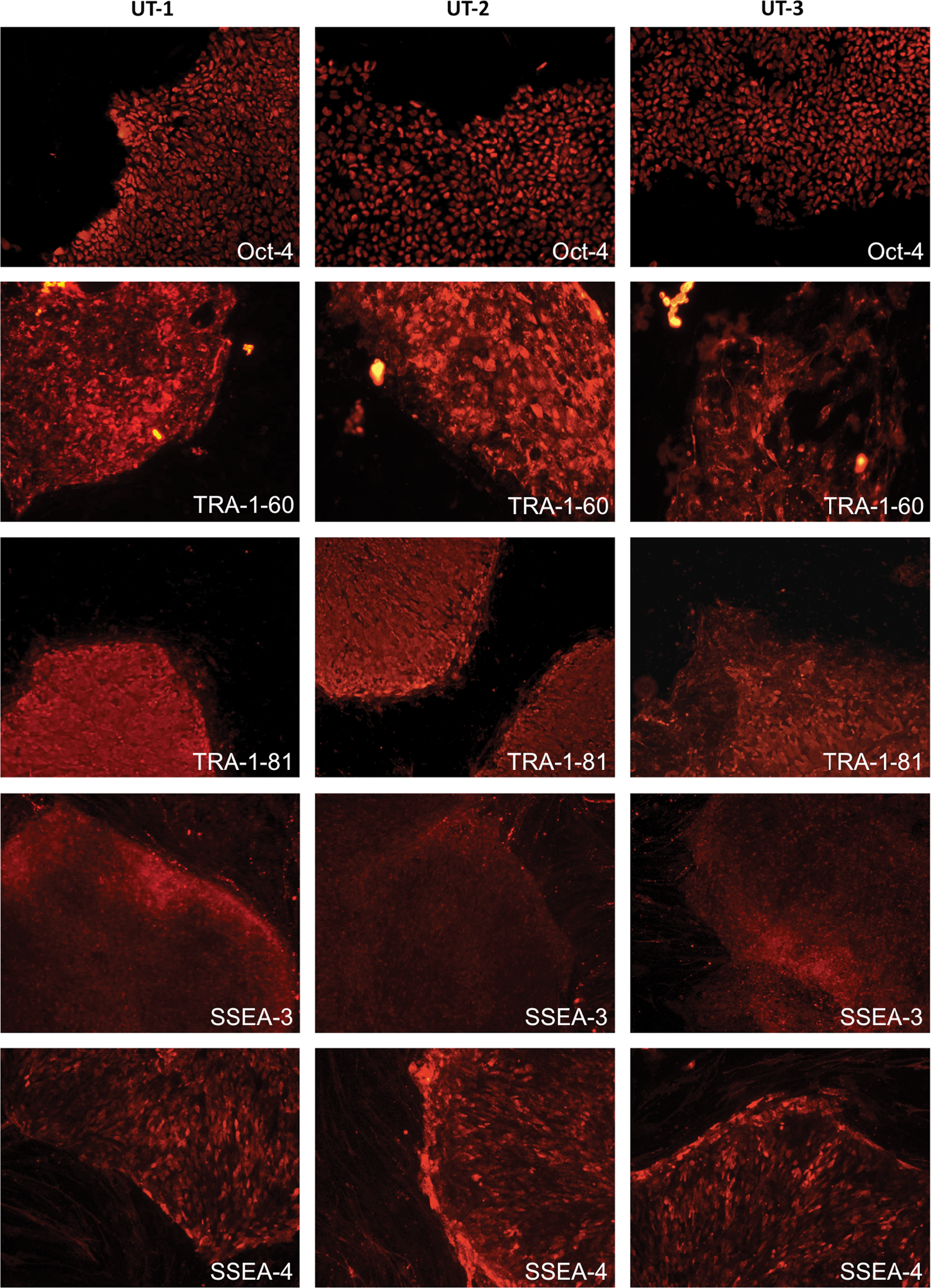
Baboon ESC lines demonstrated positive fluorescent immunocytochemical staining for pluripotency markers OCT-4, TRA-1-60, TRA-1-81, stage-specific embryonic antigen (SSEA)-3, and SSEA-4. Color images available online at
Baboon ESC-derived teratoma formation
Teratomas were detectable by palpation after 8–10 weeks, and dissection was performed on weeks 12 and 17. Thirteen teratomas from 3 ESC lines were retrieved, 6 from kidney capsule, 6 from testis, and 1 in the scrotal cavity adjacent to testis. H&E staining revealed structures in all 3 germ layers, including ectoderm, endoderm, and mesoderm (Fig. 4). The formation of teratomas varied by different injection sites observed in this study. ESC injected into the kidney capsule formed teratomas faster than those into the testis and resulted in larger teratomas. Teratomas formed in the testis injection sites were smaller, with a higher percentage of cystic structure. Injection in the kidney capsule resulted in higher percentage of teratomas (75%, 6 out of 8) than that in testis (60%, 6 out of 10; or 70%, 7 out of 10 including the teratoma found adjacent to the testis). Baboon ESC injections administrated i.m. in hind legs of NOD/SCID mice did not yield any teratoma in this study (0 out of 8).
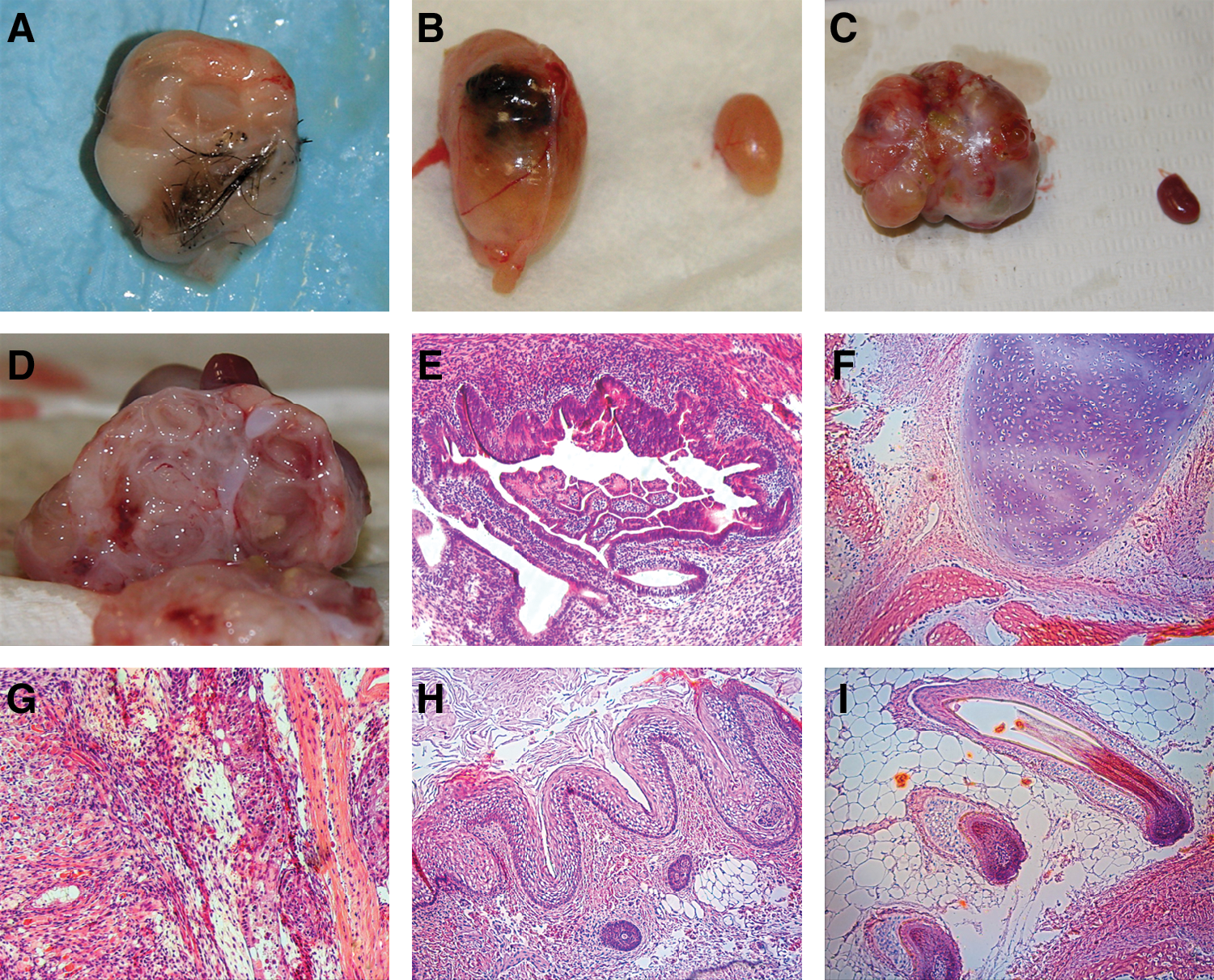
Teratoma dissection from ESC-injected NOD/SCID mice showed a mixture of well-differentiated tissues, including hair, pigment epithelium, and cartilages
EB formation and initial in vitro differentiation
A pilot study of in vitro differentiation of EB formation showed development of spherical EB in the medium. When embedded in Matrigel, initial protrusions were formed from EB into the Matrigel starting a week after the embedding (Fig. 5A–C). Spontaneous differentiation of ESC was observed, with majority in neural-lineage morphology. In addition, we have seen spherical hollow shape cellular structure bearing a resemblance to blastocyst, with mural TE-like layer on the outside and tiny cell clumps inside (Fig. 5D). Although the data are preliminary, there is implication for further development of induced differentiation with cell lineages derived from the baboon ESC.
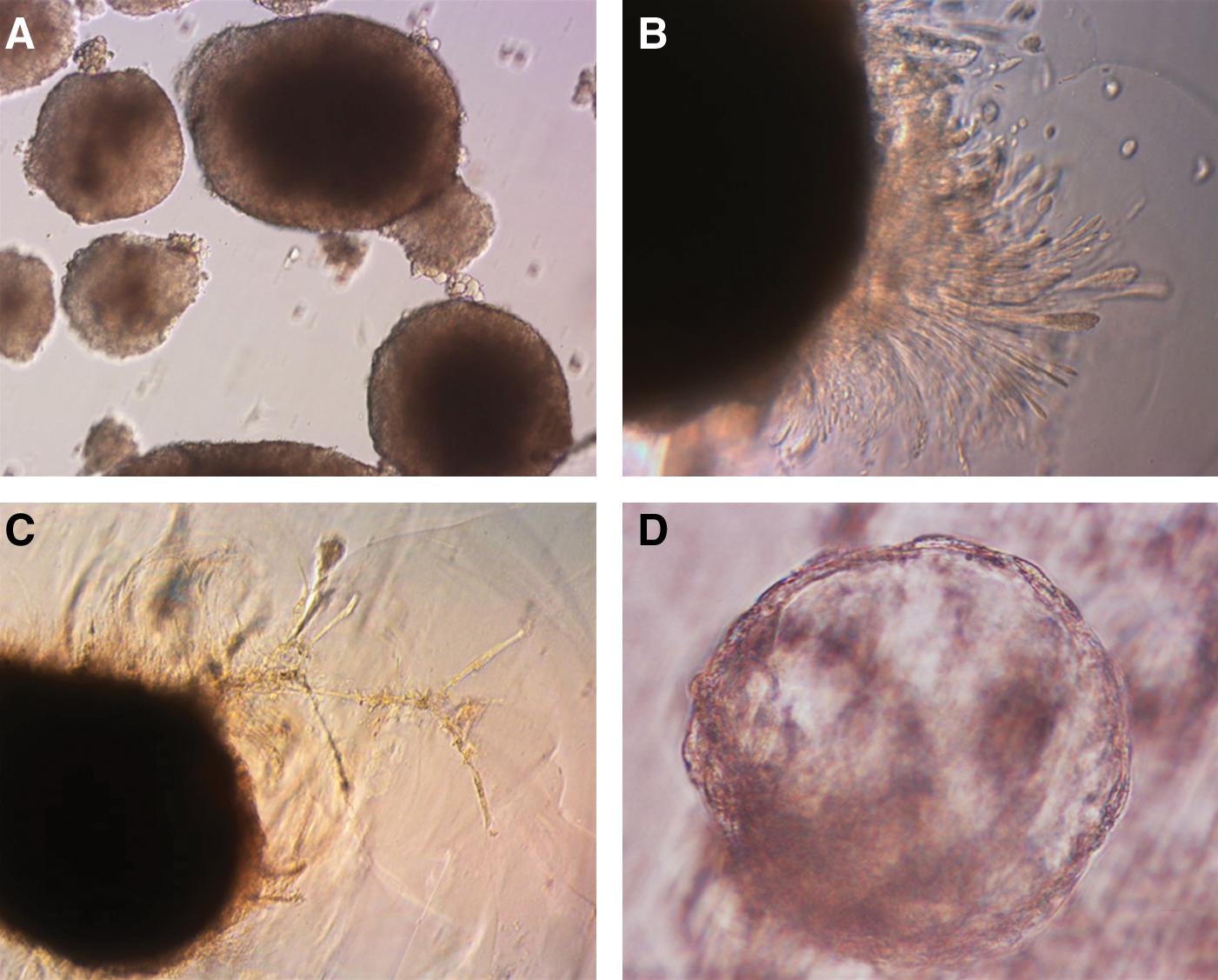
Baboon ESC aggregation formed spherical embryoid body in culture
Discussion
In this study we report the laser-assisted isolation of ICM from IVF-ICSI derived from baboon blastocysts, and successful derivation and characterization of 3 novel baboon ESC lines. Because of the physiological similarities between NHP and humans, NHP ESCs hold great potential for preclinical assessment research of stem cell therapeutic applications. Among various NHP species utilized for biomedical studies, ESC from baboon is less characterized as compared to studies in other NHP species. All of our baboon ESC lines were derived from preimplantation embryos at expanded blastocyst stage with high-quality ICM morphology. Although it is possible to derive ESC from early stage mouse and human single blastomeres [27 –29], as well as from poor quality human preimplantation embryos [30], we have not been able to derive ESC from morula-stage baboon embryos. Further investigation in the modification of our current protocol may enhance the success rate of baboon ESC derivation from morula-stage embryos or single blastomeres.
We applied laser-assisted dissection, instead of immunosurgery, to isolate ICM from baboon blastocysts. The ICM was intact after laser dissection and such technique may possibly eliminate the risk of exposing ICM cells to pronase and complement, which can accidentally damage the cells during immunosurgery. Previous study showed that laser-assisted isolation technique is reliable to harvest ICM for ESC derivation [31]. In addition, we utilized laser at a lower intensity setting to trim out leftover mural TE cells that appeared to be annealed to the ICM, a method that can be useful to minimize potential subsequent outgrowth and differentiation of trophoblastic cells from the ICM cell cluster placed onto MEF during initial ESC derivation. It is possible that such laser-assisted ICM isolation and potential TE cell trimming may have contributed to the high percentage success of initial ICM attachment to MEF and subsequent baboon ESC maintenance and expansion.
The morphology of the 3 baboon ESC lines derived in our study is more similar to human and other NHP ESCs, with flattened cell colonies of loose aggregates, as compared to the domed shape of mouse ESC colonies. Occasionally, the center part of colonies formed an upward structure due to the high density of cell proliferation after the first few days in each passage, and often with subsequent differentiation starting from such central area, which required the removal of the upward structure or direct splitting the whole colony to new MEF feeder layer. Interestingly, baboon ESC morphology is more similar to human ESC than to rhesus monkey ESC. Among the 3 ESC lines, 1 was female karyotype and the other 2 male. A recently published study on ESC lines derived from another baboon species (P. anubis) demonstrated derivation of 2 ESC lines of female karyotype [13]. Future comparison between the 2 male ESC lines we derived and the female ESC lines may reveal any uniform or diverse characteristics of baboon ESC lines in different species as well as gender origins.
Fluorescent ICC staining on 3 baboon ESC lines showed positive expression of pluripotency markers. We found mixed intensities of expression with SSEA-3, compared to the more steady expression of SSEA-4, in those 3 ESC lines. All demonstrated pluripotency by forming teratomas in SCID mice. SSEA-3 and −4 are not essential for human ESC pluripotency [32], and expression of these markers is vague in African green monkey ESC [12]. SSEA-3 expression is absent in cynomolgus monkey ESC [7], whereas both SSEA-3 and −4 are expressed in the rhesus monkey ESC [3,10]. In addition, SSEA-1 expressed in mouse ESC is not expressed in ESC of primate species [7,10,11,33,34]. A similar phenomenon of mixed expression patterns of pluripotency markers has been found in marmoset ESC lines. One study showed weak immunostaining with SSEA-3 and TRA-1-81 as compared to markers SSEA-4 and TRA-1-60 [4], whereas another study showed consistent expression of all 4 markers [11]. With the variety of ESC characteristics in different species, an array of ESC markers, no single marker, should be used to characterize new ESC lines.
Among pluripotency gene markers assessed by RT-PCR, REX-1 was expressed in all 3 cell lines as well as the baboon second trimester fetal fibroblast cells used for control. Differences in REX-1 expression were found in ESCs derived from various primate species, including human [34,35], African green monkey [12], cynomolgus monkey [12], as well as in a recently published study on olive baboon ESC lines [13], but negative in rhesus monkey ESC lines [10]. Another study showed that REX-1 is expressed in early passages of human ESC-derived fibroblast-like cells [36]. The finding of REX-1 amplification in the early pregnancy baboon fetal fibroblasts, where all other pluripotency markers were absent, suggests that REX-1 expression is conserved in certain types of differentiated fetal baboon tissues. REX-1 as pluripotency marker should be combined with other commonly used markers when characterizing new ESC lines in NHP.
Teratomas formed in SCID mice injected with baboon ESC demonstrated the ability to differentiate in vivo. ESC injected into the kidney capsule formed larger teratomas faster, whereas those injected into the testis showed smaller teratoma formation and also a higher percentage of cystic structure, similar to the previous observation on human ESC-derived teratomas [37]. Our findings are in agreement with previous studies in human ESC that injections into kidney capsule results in more consistent teratoma formation than those in the testis or i.m. sites [38].
The preliminary study of EB formation showed the capability of baboon ESC lines derived in this study to differentiate in vitro. The cellular protrusions of EB into the 3-dimensional environment of Matrigel were similar to those previously reported in human EB and rhesus monkey embryos. All showed cellular proliferation that may mimic trophoblastic cell outgrowth when interacting with extracellular matrix in a 3-dimensional setting [39 –42]. Susceptibility to spontaneously differentiation with neural lineages as default, as shown in the present study, has been found in other species ESC culture, including mouse and rhesus monkey [43,44]. In addition, although we did not characterize the spherical structure bearing a resemblance to blastocyst, ESC-derived trophoblastic cell formation has been demonstrated in humans [5,40,45] and rhesus monkey [3,42], and the possibility of de novo embryo-like derivation from ESC deserves future investigation.
The cell culture protocol we applied to derive baboon ESC was based on human and rhesus monkey ESC culture methods, with modifications, including a higher concentration of bFGF and absence of LIF. In this study we found that the requirement of bFGF to maintain baboon ESC colonies in undifferentiated status is higher than it is indicated in human and other NHP ESC protocols. LIF has been widely used in primate ESC derivation and maintenance protocols [4,5,11,12], and later found less or not effective for maintaining undifferentiated status of ESC lines from human, cynomolgus, and rhesus monkey [7,10,34]. LIF receptor message is low or variable in human ESC lines compared to mouse ESC lines [34].
Conclusion
We described the derivation and characterization of 3 baboon ESC lines from IVF-ICSI-derived blastocysts. These cell lines, along with other previously established NHP ESCs, have the potential to extend our knowledge in developmental biology, regenerative medicine, and broaden opportunities for future investigations and applications in basic science and translational research, including ESC gene regulation and modification, epigenetics, genetically identical stem cell derivation from the patient or host, and preclinical safety assessment of in vivo stem cell therapy.
Footnotes
Acknowledgments
We acknowledge the faculty and staff at University of Texas Health Science Center at San Antonio for their assistance and technical services: Gregory A. Langone on teratoma paraffin block preparation; Dr. Cun Li and Dr. Michiyo Nakamura for providing baboon second-trimester fetal fibroblasts; Department of Laboratory Animal Research on baboon surgical procedure assistance and animal care; Dr. Victoria Frohlich at Core Optical Imaging Lab, and Edward A. Kalmykov at Dr. Bruce J. Nicholson's laboratory, Department of Biochemistry, on assisting partial microscopic image capture; Dr. Marlo M. Nicolas at Department of Pathology on examining images of teratoma H&E slides. We thank Dr. Shoukhrat M. Mitalipov at the Oregon Health and Science University for advice on ESC culture and ICC protocol. We are grateful to the AT&T Foundation for sponsoring this study.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.