
Abstract
The restricted gene expression of a differentiated cell can be reversed by forming hybrid with embryonic stem cells (ESCs). The resulting hybrid cells showed not only an ESC-specific marker expression but also a differentiation potential similar to the pluripotent fusion partner. Here, we evaluated whether the tetraploid fusion hybrid cells have a unique differentiation potential compared with diploid pluripotent cells. The first Oct4-GFP–positive cells were observed at day 2 following fusion between ESCs and neurosphere cells (OG2+/−
Introduction
M
In our previous study, we observed that the somatic cell nuclei of the fusion hybrid between ESCs and neurosphere cells (NSCs) reestablished genome-wide gene expression patterns, except for the Xist gene [10]. When male NSCs were fused with ECCs or ESCs, the active X chromosomes of the ECCs, which express low levels of Xist, gradually changed to the somatic-specific active X chromosome (no expression of Xist). Ambrosi et al. also reported that the gene expression profile of the fusion hybrid cells slightly differ from that of the ESCs [11]. These studies indicate that the characteristics of the reprogrammed fusion hybrid cells are very similar, but not identical, to those of ESCs.
A recent study suggested that hybrid cells were able to generate chimeras at a high rate after blastocyst injection, comparable to that of diploid ESCs [12]. However, we hypothesize that tetraploid hybrid cells have a differentiation potential that differs from that of diploid ESCs. In the present study, we evaluated whether tetraploid pluripotent hybrid cells have a unique differentiation potential compared with diploid pluripotent cells. We found that tetraploid hybrid cells formed a chimera and contributed to the placenta after blastocyst injection.
Materials and Methods
ESC propagation
ESCs were grown on mouse embryonic fibroblast feeder cells inactivated with 0.01 mg/mL Mitomycin C in standard ESC media, high-glucose DMEM (Gibco BRL) containing 15% fetal calf serum (FCS; Hyclone), 1× penicillin/streptomycin/glutamine, 1× nonessential amino acids (Gibco BRL), 0.1 mM β-mercaptoethanol, and 1000 U/mL leukemia inhibitory factor (LIF) (ESGRO; Chemicon).
NSC culture
The ROSA26 strain was crossed with the OG2 transgenic strain [13] over several generations to produce compound homozygous mice for the lacZ (lacZ+/+) and Oct4-GFP (Oct4-GFP +/+) transgenes supporting the assumption that the 2 markers are on separate chromosomes. To derive NSCs, ROSA26/OG2 homozygous male mice were crossed with outbred ICR females to produce litters of pups hemizygous for both transgenes. Brain tissue was collected from 1- to 5-day-old ROSA26/OG2 hemizygous mice. The cortex was dissected from the brain of each mouse and enzymatically dissociated in 0.7 mg/mL hyaluronic acid, 0.2 mg/mL kynurenic acid, and 1.33 mg/mL trypsin in Hank's balanced salt solution with 2 mM glucose at 37°C for 30 min. The dissociated cells were passed through a 70-μm nylon mesh (Falcon) to remove large clusters. The cells were centrifuged at 200 g for 5 min and collected by centrifugation in 0.9 M sucrose in 0.5× Hank's balanced salt solution for 10 min at 750 g. The cell pellet was resuspended in 2 mL of culture medium, placed on top of 10 mL of 4% bovine serum albumin in Earle's blanced salt solution (EBSS) solution, and centrifuged at 200 g for 7 min. The culture medium was supplemented with 20 ng/mL epidermal growth factor (EGF; Collaborative Biomedical Products), 20 ng/mL basic fibroblast growth factor, B27 supplement (Life Technologies), 8 mM HEPES, 2 mM glutamine, 100 U/mL penicillin, and 100 mg/mL streptomycin in DMEM-F12 medium (Life Technologies). Primary neurospheres were cultured for 5–8 days and used for fusion.
Cell fusion and subsequent culture
ESCs were mixed with NSCs in a ratio of 1:1 and were washed in phosphate-buffered saline (PBS). The mixture was centrifuged for 5 min at 130 g in conical tubes (Falcon). The supernatant was thoroughly removed, and 1 mL of a prewarmed solution of 50% polyethylene glycol 1500 (PEG1500; Roche) was added to the cell pellet over 1 min. An additional 20 mL of Dulbecco's modified Eagle's medium (DMEM) was added to the cell suspension over 5 min with constant stirring. The cells were centrifuged at 130 g for 5 min to remove the PEG and were further washed gently with DMEM and cultured in ESC medium supplemented with 20 ng/mL of EGF and basic fibroblast growth factor. Cells were selected in ESC medium containing 250 μg/mL G418 the next day after PEG fusion to eliminate nonfused ESC for 9–10 days. During the G418 selection, these ESC hybrid clones were subcultured every 2 or 3 days. Nonfused NSCs did not form an ESC-like colony.
Karyotype analysis
A 50% confluent cells in a 6-cm dish were treated with 0.3 mg/mL Nocodazole for 3 h. Cells were recovered by trypsinization and replated on gelatin-coated (0.1% in PBS) dishes for 30 min to induce attachment of feeder cells. Nonattached cells were recovered and treated with hypotonic (0.56%, w/v) KCl solution for 15 min. The cells were spun down at 500 rpm, fixed by washing 3 times in fresh fixative (3:1 methanol:acetic acid), and dropped onto clean glass slides. The slides were air-dried, stained with 3% Giemsa (Sigma), and observed under a microscope.
Blastocyst injection
Blastocysts were obtained from B6C3F1 nontransgenic mice. ESC-NSC hybrid cells were prepared by trypsinization and placed in a drop of PBS under mineral oil. The blastocysts were placed in an adjacent drop of PBS containing 10% fetal bovine serum (v/v) and 15–20 sorted green florescent protein (GFP)-positive cells were picked up under fluorescence microscope with the injection pipette and injected into the blastocyst. Eight to 15 injected blastocysts were transferred to the uterus of each pseudo-pregnant ICR mice.
X-gal staining
To stain the cells, culture dishes were washed with PBS and fixed with 4% formaldehyde at 4°C for 1 h. The fixed plates were washed with PBS and incubated overnight at 37°C in PBS supplemented with 1 mg/mL 5-bromo-4-chloro-3-indolyl-galactosidase (X-gal; Promega, Madison, WI), 5 mM K2Fe(CN)6, 5 mM K4Fe(CN)6, and 1 mM MgCl2.
For whole fetal embryo staining, collected fetuses were rinsed with PBS and fixed for 1 h at 4°C in 4% formaldehyde. Fetuses were rinsed 3 times at room temperature in PBS supplemented with 5 mM EGTA, 0.01% deoxycholate, 0.02% NP40, and 2 mM MgCl2. The specimens were washed with PBS and then stained in X-gal staining solution as described above. Blue staining was visualized by light microscopy.
Flow cytometry
Hybrid cells were dissociated with 0.25% trypsin/EDTA, neutralized with DMEM with 10% FCS, washed with PBS, and then filtered through a 40-μm nylon mesh to remove large clusters. The cells were resuspended in PBS containing 0.2% bovine serum albumin and the aliquots of 5 × 106 cells/mL were transferred into polystyrene tube (Falcon 2054). Highly intense GFP-positive cells were isolated by fluorescence-activated cell sorting (FACS) with a FACStar Plus (Becton Dickinson) equipped with dual 488 nm argon and 633 nm helium neon lasers. For separation of diploid cells, dissociated hybrid cell mixture was treated 0.5 g/mL colcemid for 12 h, washed with PBS, and stained with Hoechst (5 g/mL) for 90 min at 37°C. Cells were washed with PBS and processed as described above.
Reverse transcription–polymerase chain reaction
Total RNA was extracted using ToTALLY RNA kit (Ambion) and reverse transcribed by RETROscript kit (Ambion). The resultant RNA was subjected to DNase treatment and removal to remove any contaminating DNA (Ambion). For reverse transcription–polymerase chain reaction (RT-PCR), 1 μg of total RNA was used with a RETROscript kit (Ambion) following the manufacturer's protocol. Primer sequences are Oct4-For 5′-gaacagtttgccaagctgctg-3′, Oct4-Rev 5′-ccggttacagaaccatactcg-3′; EGFP-For 5′-tgaagttcatctgcaccacc-3′, EGFP-Rev 5′-tcaccttgatgccgttcttc-3′; Rex-1-For 5′-aaagtgagattagccccgag-3′, Rex-1-Rev 5′-tccccatccccttcaatagca-3′; nanog-For 5′-agggtctgctactgagatgctctg-3′, nanog-Rev 5′-caaccactggtttttctgccaccg-3′; GluR6-For 5′-gctgttcagtccatctgcaa-3′, GluR6-Rev 5′-tccacatcaagagcgaagag-3′; HNF3β-For 5′-agaagcaactggcactgaagga-3′, HNF3β-Rev 5′-gtagtgcatgacctgttcgtag-3′; α-cardiac myosin-For 5′-ggcacagaagatgctgacaa-3′, α-cardiac myosin-Rev 5′-cgaacatgtggtggttgaag-3′; βH1-For 5′-ctcaaggagacctttgctca-3′, βH1-Rev 5′-agtccccatggagtcaaaga-3′; α-fetoprotein-For 5′-aggaggagtgcttccagaca-3′, α-fetoprotein-Rev 5′-tgcgtgaattatgcagaagc-3′; and β-actin-For 5′-cgtgcgtgacatcaaagagaagc-3, β-actin-Rev 5′-atctgctggaaggtggacagtgag-3′.
In vitro differentiation
ESCs were recovered by trypsinization and replated on gelatin-coated (0.1% in PBS) dishes for 30 min to induce attachment of feeder cells. Nonattached cells were recovered and seeded onto a bacteriological dish in DMEM (10% FCS) in the absence of LIF for 4 days. After embryoid bodies were formed, they were treated with 0.5 M retinoic acid for 4 days in bacteriological dish and plated onto a tissue culture dish for 8–12 days. These differentiated cells obtained were used for RNA isolation and RT-PCR.
Results
Reprogramming of Oct4-GFP in ESC-NSC hybrids
The Oct4 gene is active in ESCs and germ cells, whereas it is inactive in somatic cells. Reestablishment of pluripotency in NSCs by fusion with ESCs can be estimated by examining the GFP-positive signal, because NSCs were prepared from OG2/ROSA26 hemizygous transgenic mice carrying the ubiquitously expressed lacZ transgene and GFP under the Oct4 promoter (Oct4-GFP). Reprogramming of Oct4-GFP in ESC-NSC hybrid cells was examined in 3 independent PEG-mediated fusion experiments. After fusion, these ESC hybrid clones were subcultured in G418-containing ESC medium every 2 or 3 days for 9–10 days and then maintained in nonselective ESC medium. Oct4-GFP expression was assessed by GFP fluorescence as shown in Fig. 1. The first GFP-positive cells were observed at day 2, about 45 h after cell fusion (Fig. 1A, B). This lag time for reprogramming of the somatic nuclei was almost identical to the 48 h previously reported by Tada et al. (2001), indicating that the different fusion methods (electrofusion vs. PEG-mediated fusion) produce similar results [9]. Most of the remaining cell colonies expressed GFP during subsequent selection with neomycin (Fig. 1C, D), and colony morphology was similar to that of ESCs. These Oct4-GFP–positive cells could be recovered from cryopreservation (data not shown). To confirm that these cells were hybrids, we examined the morphology of nuclei and nucleoli of GFP-positive cells and determined karyotypes. The fused GFP-positive cells contained enlarged nuclei with multiple nucleoli (Fig. 1E, F), and tetraploid karyotypes (70–80 chromosomes per cell) were observed in the fusion mixture (Fig. 1G). The GFP-positive cells were sorted by FACS and used for further experiments. Sorted cells are all X-gal positive (Supplementary Fig. S1, B; Supplementary Data are available online at
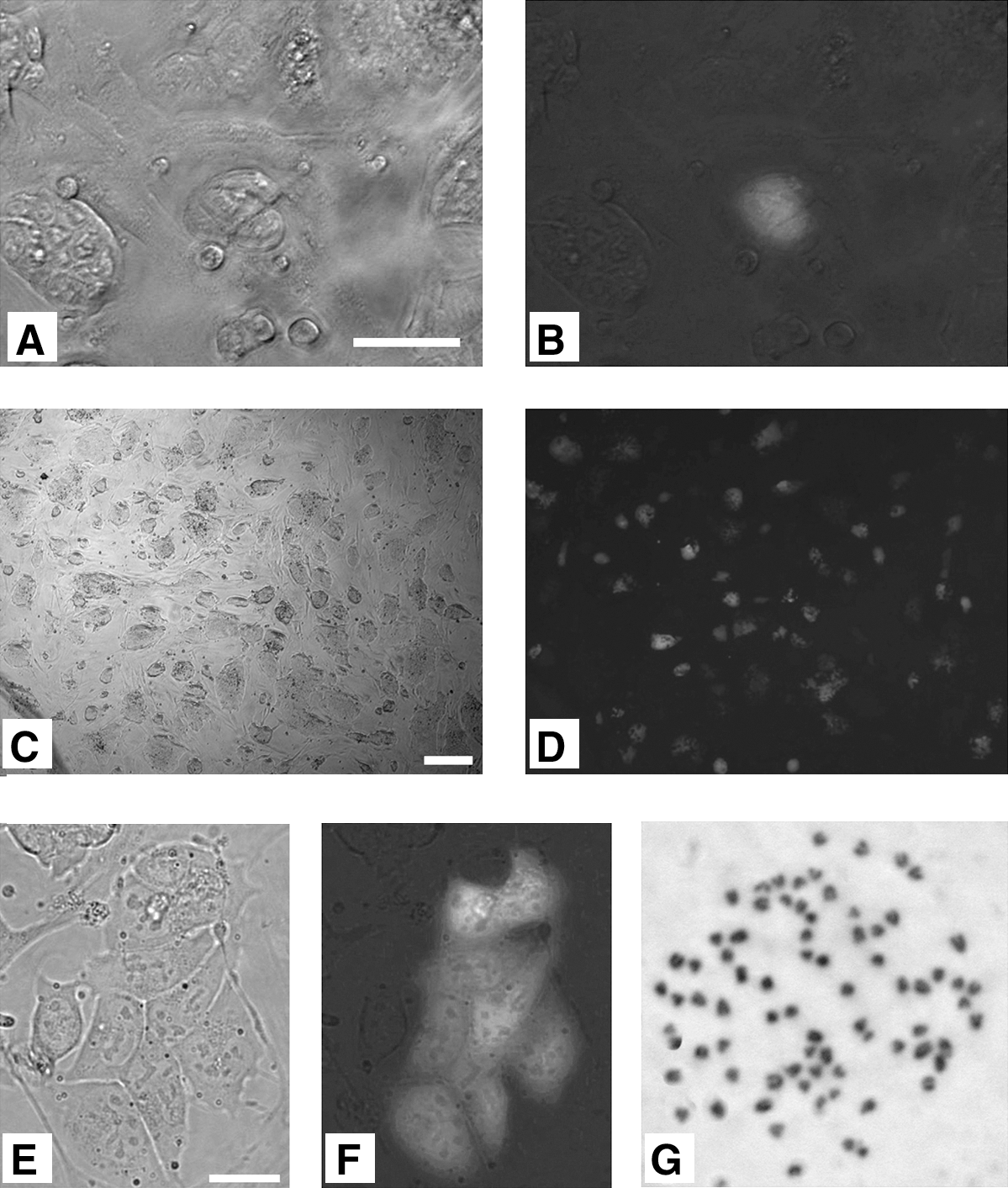
Reprogramming NSCs by fusion with ESCs. Bright field
Reprogramming rate of ESC-NSC hybrids
Immediately after fusion induction, the cell mixture is thought to consist of unfused single ESCs, unfused single NSCs, ESC-ESC fused cells, NSC-NSC fused cells, and ESC-NSC hybrid cells. After G418 selection, only nonfused single NSCs, NSC-NSC fused cells, and ESC-NSC hybrids are expected to survive. Because NSCs were prepared from OG2/ROSA26 hemizygous transgenic mice, all the NSCs and the fused hybrids were lacZ positive and neomycin resistant. Of these cell types, only ESC-NSC hybrids would be able to form ESC-like colonies (Fig. 2A). After 10 days of G418 selection, however, unfused ESCs in the fusion mixture (indicated by arrows) were resistant to G418 (Fig. 2B, C). The colonies indicated by arrowheads are the Oct4-GFP–negative ESC-NSC hybrids. Thus, some of G418-resistant Oct4-GFP–negative cells may be hybrid cells (X-gal stained). The reprogramming rate, therefore, was estimated by counting colonies that were both Oct4-GFP positive and lacZ positive at early and later passages (Table 1). In 3 different experiments, about 94.5% of the hybrid cell NSC genomes were reprogrammed by fusion with pluripotent ESCs (passage 5), and the reprogramming rate was stable over the subsequent passages (96.4% at passage 13).
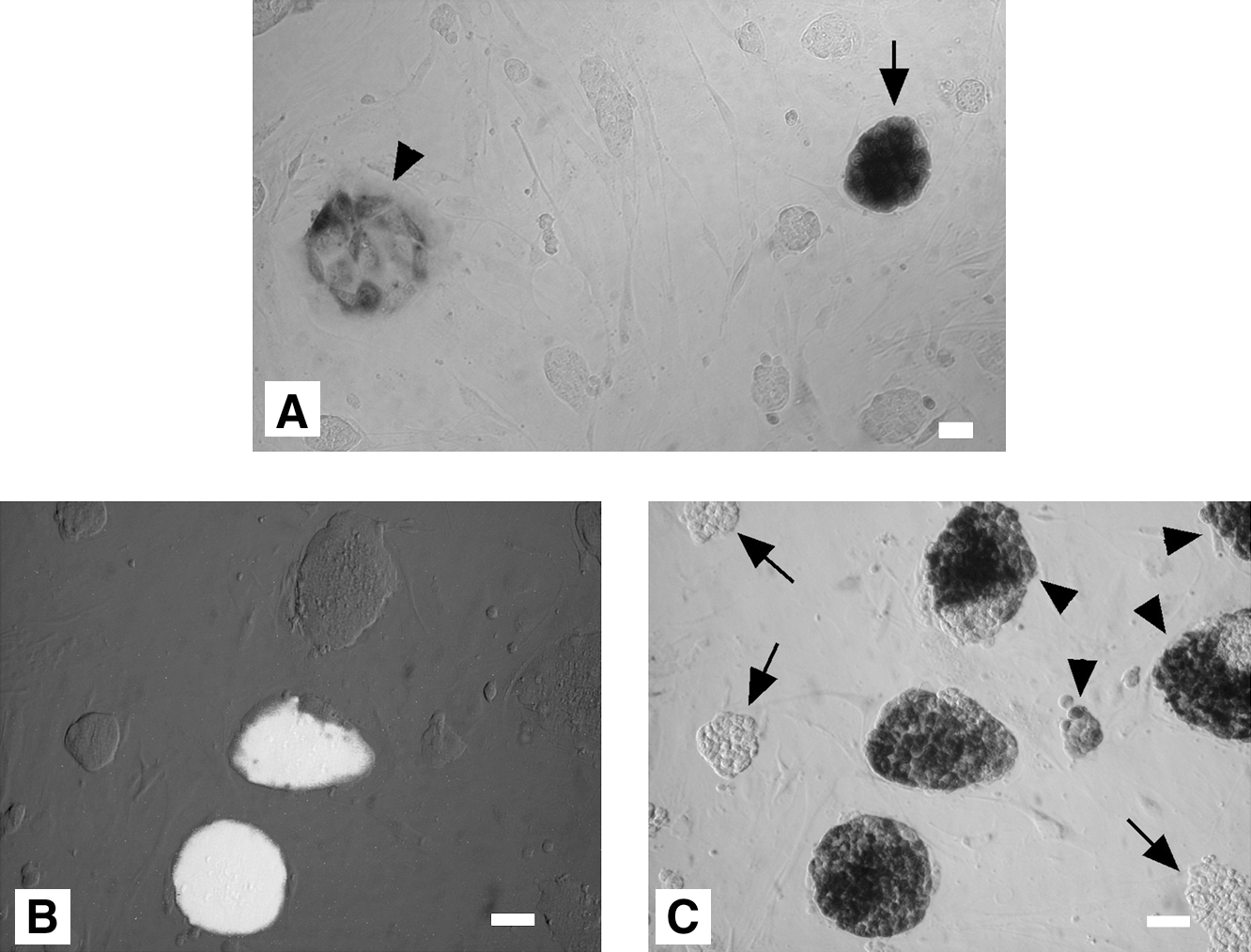
X-gal staining of ESC-NSC hybrid cell colonies.
ESC-NSC hybrid cells expressing Oct4-GFP.
ESC-NSC hybrid cells.
ESC, embryonic stem cell; NSC, neurosphere cells.
Embryonic and extraembryonic developmental potential of ESC-NSC hybrids
To further characterize the hybrid cells, gene expression patterns were compared with normal E14 ESC lines by RT-PCR analysis (Fig. 3). The hybrid cells expressed markers of undifferentiated ESCs, including Oct4, Rex-1, and nanog, but did not express glutamate receptor 6 (GluR6), which is a neural cell marker. These results show the reactivation of pluripotency-specific markers and inactivation of tissue-specific markers. Hybrid cells were able to form embryoid bodies, which were seeded into tissue culture dishes in the absence of leukemia inhibitory factor. Embryoid bodies were differentiated into a variety of cell types in the monolayer culture. Although undifferentiated hybrid cells did not express the 3 germ layer markers, differentiated hybrid cells expressed ectoderm (GluR6), endoderm (HNF3β and α-fetoprotein), and mesoderm (βH1 and α-cardiac myosin) markers (Fig. 3). This finding indicates that the differentiation potential of the hybrid cells was similar to that of normal ESCs.
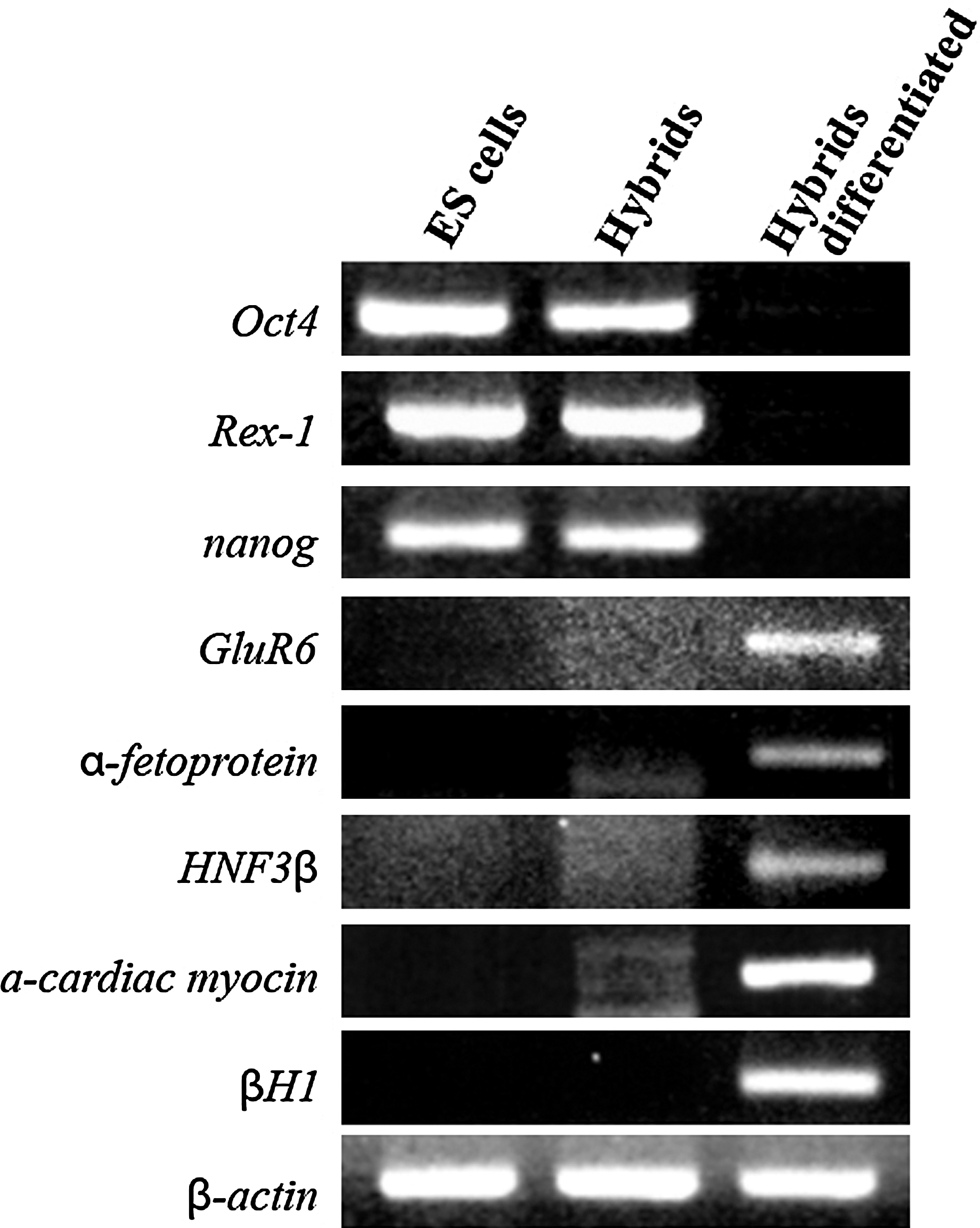
Reverse transcription–polymerase chain reaction analysis of pluripotency- and tissue-specific markers in fusion hybrid and differentiated hybrid cells. Fusion hybrid cells express pluripotency markers (Oct4, Rex-1, and nanog), but downregulated on differentiation. The differentiated hybrid cells expressed markers specific to ectoderm (GluR6), endoderm (HNF3β and α-fetoprotein), and mesoderm (βH1 and α-cardiac myosin).
To evaluate the contribution of hybrid cells during embryonic development, the Oct4-GFP–positive hybrid cells were microinjected into normal diploid blastocysts (Fig. 4A, B). Five chimeric embryos (13.5 days post coitum (dpc)) were generated by blastocyst injection. The fusion of OG2/ROSA26 double transgenic NSCs with E14 ESCs allowed us to determine the tissue contributions from the hybrid cells by X-gal staining for the lacZ marker (NSC-derived). Hybrid cells were detected in the chimera (Fig. 4C) and contributed to all 3 germ layer tissues (Supplementary Fig. S2). Contributions of hybrids to fetal tissues were detected in 5 of 18 transferred embryos by X-gal staining. Interestingly, the hybrid cells contributed to the placenta as well as embryonic tissues; 1 of 5 chimeric embryos showed placental contribution of hybrid cells (Fig. 4C, D), possibly because of the tetraploidy of the hybrid cells. Diploid ESCs normally do not contribute to the placenta, as shown by the tetraploid complementation assay [14,15]. When ESCs were injected into tetraploid blastocysts or aggregated with tetraploid 4-cell stage embryos, diploid–tetraploid chimeric embryos were formed, resulting in completely ESC-derived F0 generation mice [16,17]. This technology utilizes the fact that tetraploid cells give rise to the placental tissue of the chimera, whereas the fetal portion is derived entirely from diploid ESCs. We have performed blastocyst injection and aggregation with diploid pluripotent cells, ES cells, and induced pluripotent stem (iPS) cells. However, we could not detect placental contribution of ES cells (0/12) and iPS cells (0/80) after blastocyst injection or aggregation (representative figure was shown in Fig. 4D). Taken together, these results suggest that fusion hybrid cells can give rise to extraembryonic tissues as well as the 3 embryonic germ layers.
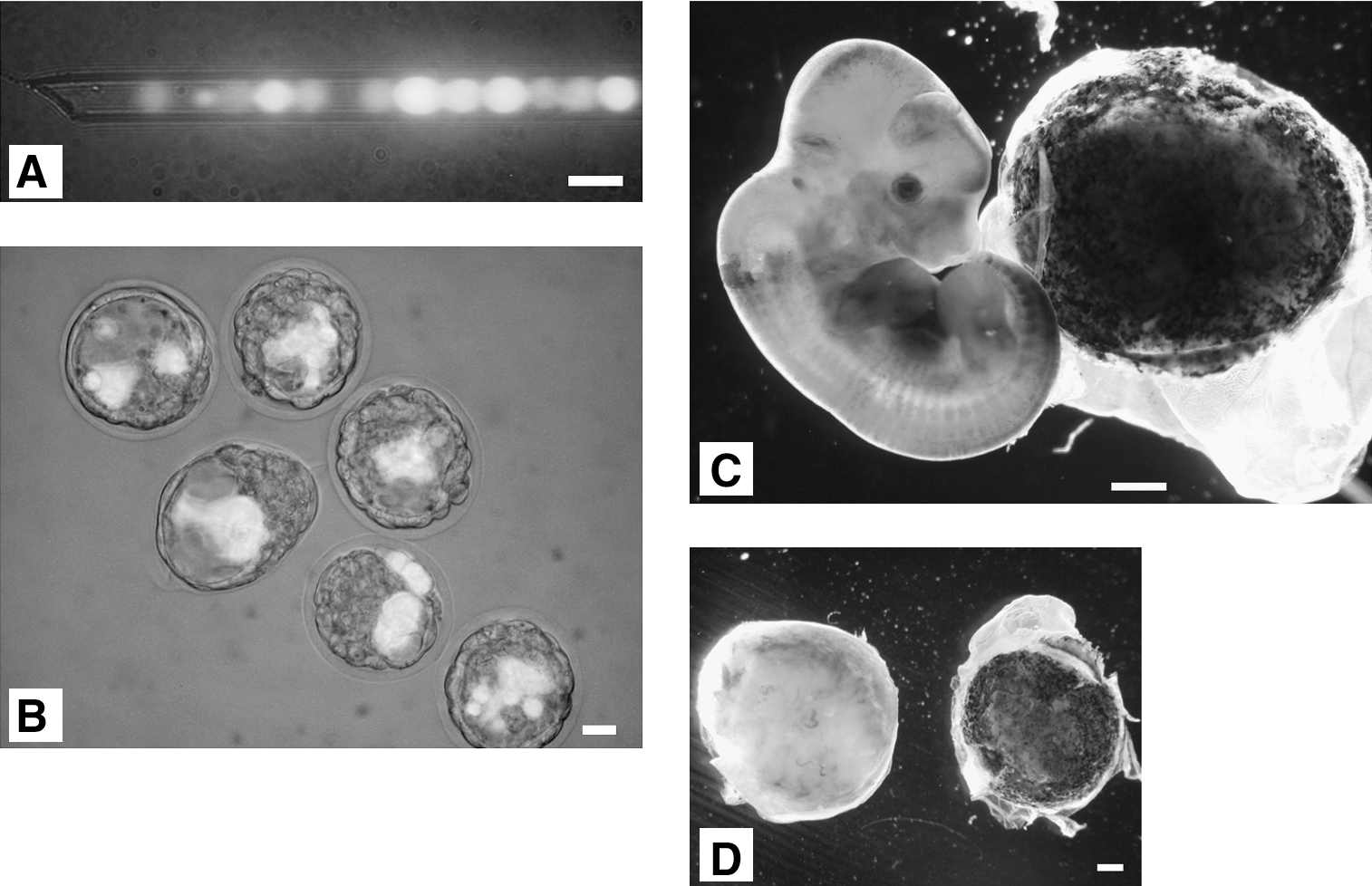
Generation of chimeric embryos after injection of hybrid cells into blastocysts and transfer to the surrogate mother.
Discussion
In the present study, we demonstrated that NSCs could be reprogrammed to the pluripotent state with high efficiency by fusion with ESCs. The reprogramming rate was estimated by counting cells that were both Oct4-GFP positive and lacZ positive (indicative of successfully reprogrammed ESC-NSC fusion hybrids; endogenous Oct4 activation of NSCs) of the X-gal–stained cells (indicative of ESC-NSC fusion hybrids irrespective of reprogramming). Cell fusion efficiently reprograms somatic cell to the pluripotent state. In the present study, reprogramming efficiency after fusion with ESCs was about 95%, indicating that once somatic cells fusion had occurred, about 95% of the somatic cells could be successfully reprogrammed (Table 1). In contrast, the reprogramming efficiency of iPS cells has been reported to be about 0.01%–0.1% [18]. However, accurate reprogramming efficiency of iPS cells is not easy to determine because of the difficulty in counting somatic cells that are not reprogrammed but are successfully infected by all 4 reprogramming factors, Oct4, Sox2, Klf4, and c-Myc.
Interestingly, not all the hybrid cells expressed Oct4, and the timing of Oct4 expression varied among the hybrid cells. There are several possible explanations for this finding. First, instead of reactivation of Oct4-GFP in the NSC nuclei, Oct4 may be extinguished by the somatic partner during hybrid cell formation [19]. Shimazaki et al. reported that Oct4 expression was downregulated in all hybrids between ECCs and fibroblasts [20]. Second, we did not synchronize the cell cycles of ESCs and NSCs. The different cell cycle stages of ESCs and the somatic cells could have affected the efficiency of reprogramming the hybrid cells. In nuclear transfer, efficient reprogramming is associated with high activity of maturation-promoting factor in the recipient oocyte, and the donor cell-cycle stage is crucial in generating a successful euploid clone [21]. Although ESCs are rapidly dividing cells with short G1 and G2 phases [22] and possess cell-cycle control that differs from differentiated cells, the reprogramming ability of ESCs may be affected by the cell cycle stage of the somatic partner [23]. Third, even in experiments where the cell cycles of recipient oocytes and donor cells were synchronized, abnormal Oct4 expression was frequently observed in mouse blastocysts after nuclear transfer of somatic cells [24,25]. Oct4 was incorrectly expressed more frequently than any other related gene [25], suggesting that it is especially prone to reprogramming errors after manipulation and hybridization [24]. Fourth, neurospheres consist of cells in various stages of differentiation. A minority of the cells within an individual neurosphere are multipotent stem cells, whereas the majority of cells are progenitor or more specialized cells [26]. Therefore, the NSCs prepared for fusion represent a heterogeneous group, with some of the brain cells having less potential for reprogramming.
We demonstrated that the Oct4-GFP–expressing ESC-NSC tetraploid hybrids contributed to placental tissue (extraembryonic) as well as embryonic tissues. In contrast, diploid pluripotent stem cells such as ESCs or iPS cells do not contribute to the placenta.
The term “pluripotency” is defined as the ability to give rise to most tissues of an organism or to all 3 embryonic germ layers (ie, mesoderm, endoderm, and ectoderm) after blastocyst injection and transfer to the surrogate mother. Although somatic cells are not pluripotent, once they express Oct4 after fusion, the somatic cells give rise to all 3 germ layers [9] and the embryo can develop to term [27] despite a limited distribution of tetraploid cells at different developmental stages [15,28]. ESCs or Oct4-expressing hybrids [27,29] can contribute to live-born chimeric mice after blastocyst injection. In our chimeric embryos, the hybrid cells contributed not only to the embryo, but also to the placenta. ESCs contribute normally to epiblast-derived tissues [30], and extraembryonic lineages arise largely from the tetraploid cells [15,17]. Therefore, ESC-NSC hybrid cells may possess both ESC and tetraploid cell characteristics. Live-born tetraploid mice [31] and humans [32,33] have been reported, demonstrating that tetraploid cells can form all essential cell types and produce functional organs [34].
In the present study, we could not determine the germline contribution of hybrid cells in the chimeric embryos. Cells positive for Oct4-GFP (marker for germ cells) were not detected in the gonads of chimeric embryos (data not shown). Although Lavagnolli et al. reported that germ cell-like cells differentiated from fusion hybrid cells, the hybrid cells were found to be near diploid cells [35]. Therefore, the differentiation potential of the pluripotent fusion hybrid cells may be affected by the ploidy of the hybrid cells. The differentiation potential may not be affected by types of somatic cells, as the fusion hybrid cells lose somatic memory and obtain differentiation potential similar to their pluripotent fusion partner [3].
Footnotes
Acknowledgments
This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science, and Technology (grant nos. 20100008528 and 20100028247).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.