
Abstract
Conventionally, researchers remove spontaneously differentiated areas in human pluripotent stem cell (hPSC) colonies by using a finely drawn glass pipette or a commercially available syringe needle. However, when extreme differentiation occurs, it is inefficient to purify the remaining undifferentiated cells, as these undifferentiated areas are too small to be isolated completely with the mechanical method. Antibodies can be utilized to purify the rare undifferentiated cells; however, this type of purification cannot be used in xeno-free culture systems. To avoid the loss of valuable hPSCs, we developed a novel method to isolate undifferentiated hPSCs from extremely differentiated colonies that could be easily adapted to xeno-free culture conditions. This protocol involves dissecting away differentiated areas, dissociating the remaining colony into clumps, seeding small clumps into new dishes, and picking undifferentiated colonies for expansion. Using this method, we routinely achieve completely undifferentiated colonies in one passage without the use of antibody-based purification.
Introduction
H
hPSCs are routinely cultured on animal or human feeder cells in knockout-serum replacement (KSR) medium [6 –9] or on feeder-free matrix in chemically defined media [10 –12]. No matter which culture system is used, spontaneous differentiation of hPSCs is unavoidable. According to reports and our experience, suboptimal and nonstandard growth conditions, such as expired growth factors and reagents [3], inappropriate feeder cell density or poor quality feeder layers [13,14], unsuitable culture systems [15,16], inappropriate handling [17,18], freezing/thawing [17,19,20], derivation of new hPSC lines [21,22], or selection following gene transfection can cause extreme differentiation in hPSC colonies.
Conventionally, researchers have removed partially differentiated areas in hPSC colonies using a finely drawn glass pipette or steel needle attached to a syringe [3,23 –26]. Although this method works well in removing partially differentiated areas of hPSC colonies, this approach is not effective for extremely differentiated (>80%) hPSC colonies. If extreme differentiation occurs in hPSC colonies, it has been nearly impossible to remove completely differentiated areas.
Here, we report a new method, a combination of mechanical and enzymatic passaging, which simplifies the isolation of undifferentiated hPSC populations from extremely differentiated colonies. Using this method, it is quick and easy to derive completely undifferentiated, xeno-free colonies in a single passage.
Materials and Methods
Culture systems
Feeder-free culture of hPSCs
hESC lines, H1 and H9 (WiCell), and hiPSC lines, BJ-EOS clone 4YA and 4YF (James Ellis Laboratory; Ontario Human iPS Cell Facility, Hospital for Sick Children), were cultured on Matrigel (BD Biosciences)-coated 35-mm dishes (Nunc) in mTeSR1 medium (StemCell Technologies) following the manufacturer's instructions. Cells were clump passaged every 4 days using 1 mg/mL dispase (StemCell Technologies).
Feeder-dependent culture of hPSCs
hPSC lines were cultured on human foreskin fibroblast (HFF) feeder cells in KSR culture medium consisting of KO-Dulbecco's modified Eagle's medium supplemented with 20% serum replacement, 1.0% NEAA, 2 mM GlutaMax, 0.1 mM β-mercaptoethanol, 25 units of 25 μg/mL Pen-strep, and 10 ng/mL basic fibroblast growth factor (bFGF) (all from Invitrogen). Cells were passaged every 5–6 days using a mechanical cutting method or by clump passaging using 1 mg/mL of Type IV collagenase (Invitrogen). HFF feeder cells were prepared as described previously [27]. In addition, we used another feeder-dependent culture system (HFF feeder and HEScGRO medium consisting of basal HEScGRO medium supplemented with 20 ng/mL bFGF) as an alternative culture system to isolate undifferentiated cells from extremely differentiated hPSC colonies appearing in feeder-free culture conditions.
Rapid isolation of undifferentiated hPSCs from extremely differentiated colonies
A flow chart outlining the procedure for purifying undifferentiated hPSCs from extremely differentiated colonies is shown in Fig. 1. If only few (ie, 1–3) small undifferentiated areas are located in a colony, the completely differentiated parts of the colony are cut as much as possible and then the remaining parts containing undifferentiated hPSCs are cut into cell clumps. When several (ie, >3) island-like, undifferentiated cell areas are scattered throughout a colony (ie, extreme differentiation), we directly cut the colony into cell clumps. Cell clumps are transferred into 1.5-mL centrifuge tube containing 20–40 μL prewarmed 1 mg/1 mL enzyme solution, dissociating them with dispase for 5–10 min at 37°C (for feeder-free conditions) or with collagenase IV for 15–30 min at 37°C (for feeder-dependent conditions). Following the addition of 200 μL culture medium into the tube, cells are dissociated by vigorous micropipetting, topped up with media to 1 mL, centrifuged at 375 g for 1 min, and resuspended with small fresh medium. Cells were divided equally into culture medium with or without Y27632 (10 μM) in feeder-containing or feeder-free culture wells of 4-well plates or 35-mm dishes. In each case, Y27632 was only used to promote the attachment of hPSCs. One day after plating cell clusters, Y27632-containing media were replaced with Y27632-free media.

Flow chart for isolation of undifferentiated hPSCs from extremely differentiated colonies.
Two to 3 days after seeding cell clusters, some undifferentiated, semidifferentiated, and differentiated colonies appeared in culture dishes or culture wells. For the next week, differentiated colonies, as well as the differentiated parts of semidifferentiated colonies, were removed on a daily basis. For feeder-free conditions, we used a 200-μL micropipette. For feeder-dependent conditions, we used a 30G1/2 needle attached to a 1-mL syringe. Differentiated cells were aspirated away upon refeeding the cells with fresh medium. One week later, the undifferentiated colonies were dissociated into cell clumps with 1 mg/1 mL dispase or collagenase IV (as above) and replated into fresh culture wells or dishes. We routinely transferred cells cultured in feeder-free conditions over to feeder-free conditions or in feeder-dependent conditions over to feeder-dependent conditions. In some cases, we transferred cells cultured in feeder-free conditions over to feeder-dependent conditions and vice versa.
Characterization of isolated hPSC sublines
Among all hPSC sublines isolated from extremely differentiated colonies, 4 randomly selected sublines, H1 (from feeder to feeder), H9 (from feeder-free to feeder), 4YA (from feeder to feeder-free), and 4YF (from feeder-free to feeder-free), were further characterized.
Immunocytochemistry was used to examine the expression of pluripotency markers using antibodies: OCT-4 (sc5279; Santa Cruz), SSEA-4, TRA-1-60, TRA-1-81, and Nanog. hPSC colonies were fixed in 4% paraformaldehyde for 15 min at room temperature and then permeabilized with 0.1% Triton-X 100 in phosphate-buffered saline (PBS) for 15 min at room temperature. The samples were then washed 3 times with PBS and incubated with PBS containing 10% normal goat serum for 30 min at room temperature. Primary antibody or negative control (mouse immunoglobulin G2b; Caltag) was applied at a 1:100 dilution at 4°C overnight. After washing 3 times with PBS, cells were incubated with the appropriate secondary antibody (Alexa Fluor 546 goat anti-mouse, 1:300; Invitrogen).
Karyotype analyses were carried out using the G-banding method. Briefly, cells were incubated with 0.1 μg/mL of colcemid at 37°C for 1 h, then trypsinized, resuspended, and incubated in 68 mM KCl for 20 min at room temperature. The samples were then fixed with 3:1 methanol:glacial acetic acid and dropped onto prepared slides to make the chromosome spreads. The dried slides were baked for 90 min at 80°C, treated with 0.05% trypsin for 30 s to 1 min, and then stained with Giemsa and Leishman's solution. At least 10 metaphase spreads were analyzed for each subline.
In vitro and in vivo differentiations were also carried out to investigate the pluripotency of hPSC sublines. The method for in vitro differentiation via embryoid body (EB) formation was described previously [3]. hPSCs were spontaneously differentiated as EBs and the resulting cell types were analyzed by marker immunofluorescence using monoclonal antibodies against β-tubulin III (Sigma T8578; 1:1,000) (ectoderm), smooth muscle actin (Sigma A2547; 1:1,000) (mesoderm), and α-fetoprotein (Sigma A8452; 1:1,000) (endoderm). Antibody localization was detected using Alexa Fluor 546 goat anti-mouse (1:300; Invitrogen).
For in vivo differentiation, hPSC colonies were cut into small clumps, centrifuged, and resuspended in culture medium without bFGF. These clumps were injected into the rear leg of 6–8-week-old severe combined immunodeficient mice (106 cells/injection). Mice were sacrificed at 8–10 weeks postinjection, and the resulting teratomas were fixed with 4% paraformaldehyde and embedded in paraffin. The paraffin blocks were sectioned at 5 μm, stained with hematoxylin and eosin, and observed under a bright-field microscope.
Results
Isolation of undifferentiated hPSCs from extremely differentiated colonies or clones
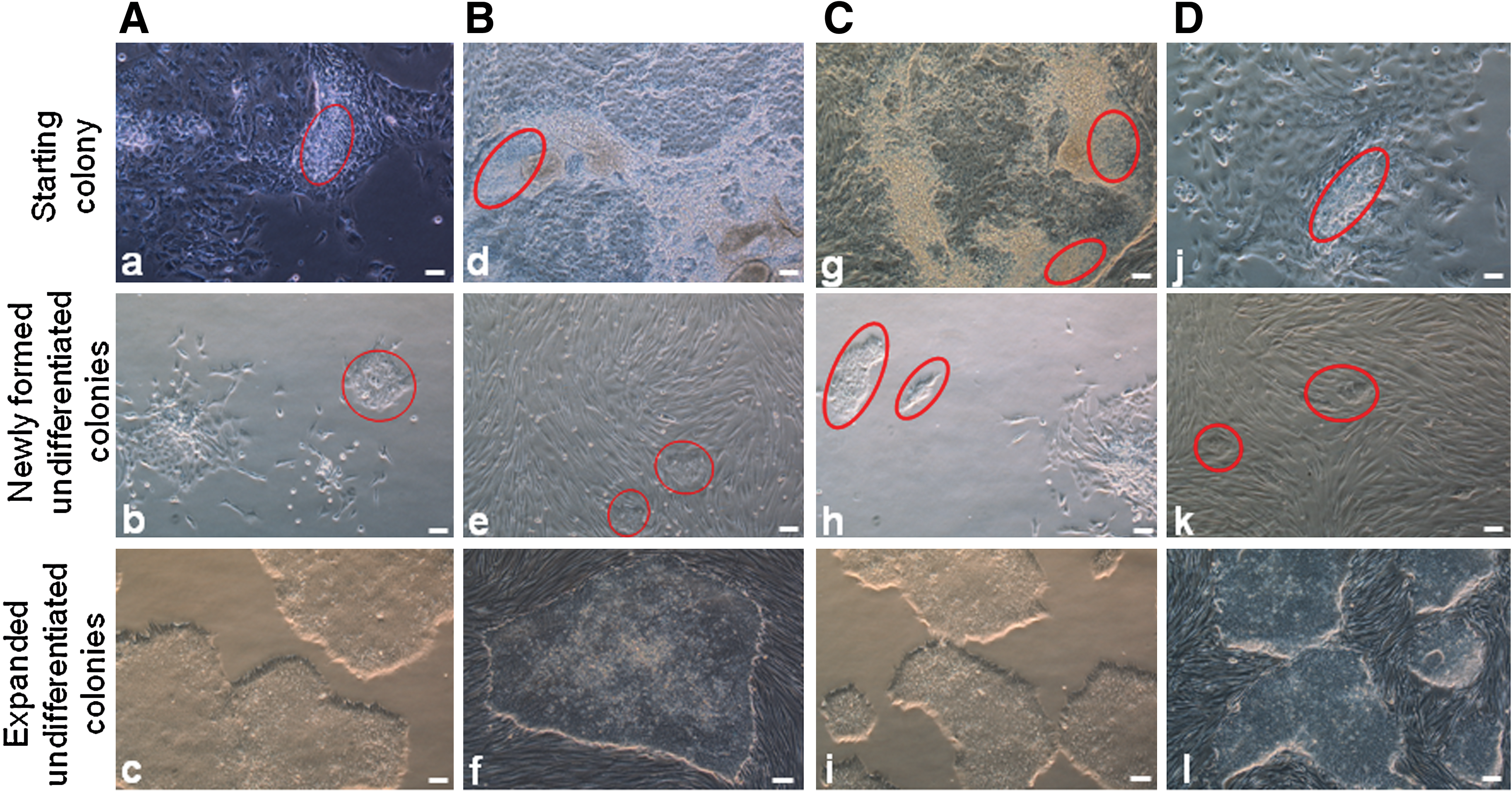
We initially developed our method using H1 and H9 hESC lines. Two experimental groups were designed for this initial trial study using feeder-dependent conditions (Table 1). Group 1 consisted of 3 hESC colonies with extreme differentiation, which were pooled for each experiment. Group 2 each had only one colony with extreme differentiation for each experiment and was repeated 3 times. In this study, Y27632 was only used to promote attachment of hPSCs. One day after plating cell clusters, Y27632-containing media would be replaced with fresh media without it. Two to 3 days after seeding cell clusters, undifferentiated colonies appeared in culture media with or without Y27632, respectively. In most cases, the numbers of undifferentiated colonies formed in the presence of Y27632 were 2 or 3 times greater than those formed in its absence (Table 1, Fig. 2A: a, b).

Comparison of newly formed hESC colonies at 2 days after plating cell clumps in medium without and with Y27632.
Similar results were observed using feeder-free conditions (Table 2, Fig. 2A: c, d). Because of the absence of feeders, we were able to observe colony formation microscopically, as early as 1 day following the seeding of cell clusters. Both in feeder-free and feeder-dependent culture systems, in the presence of Y27632, microscopic colonies formed consisting of a handful (ie, 5–10) of cells (Fig. 2B: a, c). In contrast, colonies formed in the absence of Y27632 were considerably larger (Fig. 2B: b, d). We and others have found that Y27632 is necessary to support the passaging of smaller ESC clumps [10,20,28,29]. In conditions where drug is not used, smaller clumps likely die (Fig. 2B: b, d), perhaps reflecting the noticeable difference in colony numbers following final clump passaging. Similar results were also obtained when we applied this technique to human iPSCs (Table 3). Figure 3A, B represents the isolation of undifferentiated hPSCs in the same culture system. Although there are no 2 identical colonies in these experiments, the trend is obvious that undifferentiated hESC colonies formed in the presence of Y27632 are more robust than those formed in the absence of Y27632 and that Y27632 greatly prompted the attachment of hESCs, especially the small cell clusters.
hPSC re-isolation using the same or alternative culture system.
N/A, not available.
We also explored the transfer of rescued colonies from feeder-dependent to feeder-free conditions and vice versa (Fig. 3C, D). In some cases, extreme differentiation occurring in one culture system can imply that the culture system is not optimal because of batch-to-batch variability of reagents. Here, the alternative culture system can be used to isolate undifferentiated hPSCs. From extremely differentiated H9 ESC colonies grown in feeder-free conditions and 4YA iPSC colonies grown in feeder-dependent culture system, we obtained completely undifferentiated colonies in one generation when we switched to feeder-dependent and feeder-free culture conditions, respectively (Table 4).
Cell and colony morphology after expansion of undifferentiated hPSCs
After expansion of undifferentiated hPSC colonies, cells displayed a small and round morphology accompanied by a high nucleus-to-cytoplasm ratio, a notable presence of 1–3 nucleoli, and a typical spacing between cells. In feeder-dependent culture conditions, the morphology of hPSC colonies was the same as that of their parental cell colonies grown in feeder-containing conditions. hPSC colonies were elliptical or circular in shape and formed a distinctly smooth border between hPSC colonies and the feeder cells (Fig. 3B: f and D: l). Unlike hPSCs grown on feeder cells in KSR medium, hPSCs cultured on Matrigel in mTeSR medium formed fewer multilayer and monolayer colonies (Fig. 3A: c and C: i). In summary, the newly isolated hPSCs possess the same cell and colony morphologies as their parental cells.
Characterization of newly isolated hPSCs
After expansion of newly isolated hPSCs under feeder-dependent and feeder-free conditions, we examined hPSCs for specific attributes of markers. Four randomly drawn hPSC sublines (H1, H9, 4YA, and 4YF) were assayed for expression of pluripotency markers. Using immunofluorescence, Oct4, SSEA-4, TRA-1-60, TRA-1-81, and Nanog were positive in all 4 cell lines (Fig. 4). Karyotype analysis was also performed using G-banding method. All 4 cell lines exhibited normal karyotypes (Fig. 5).
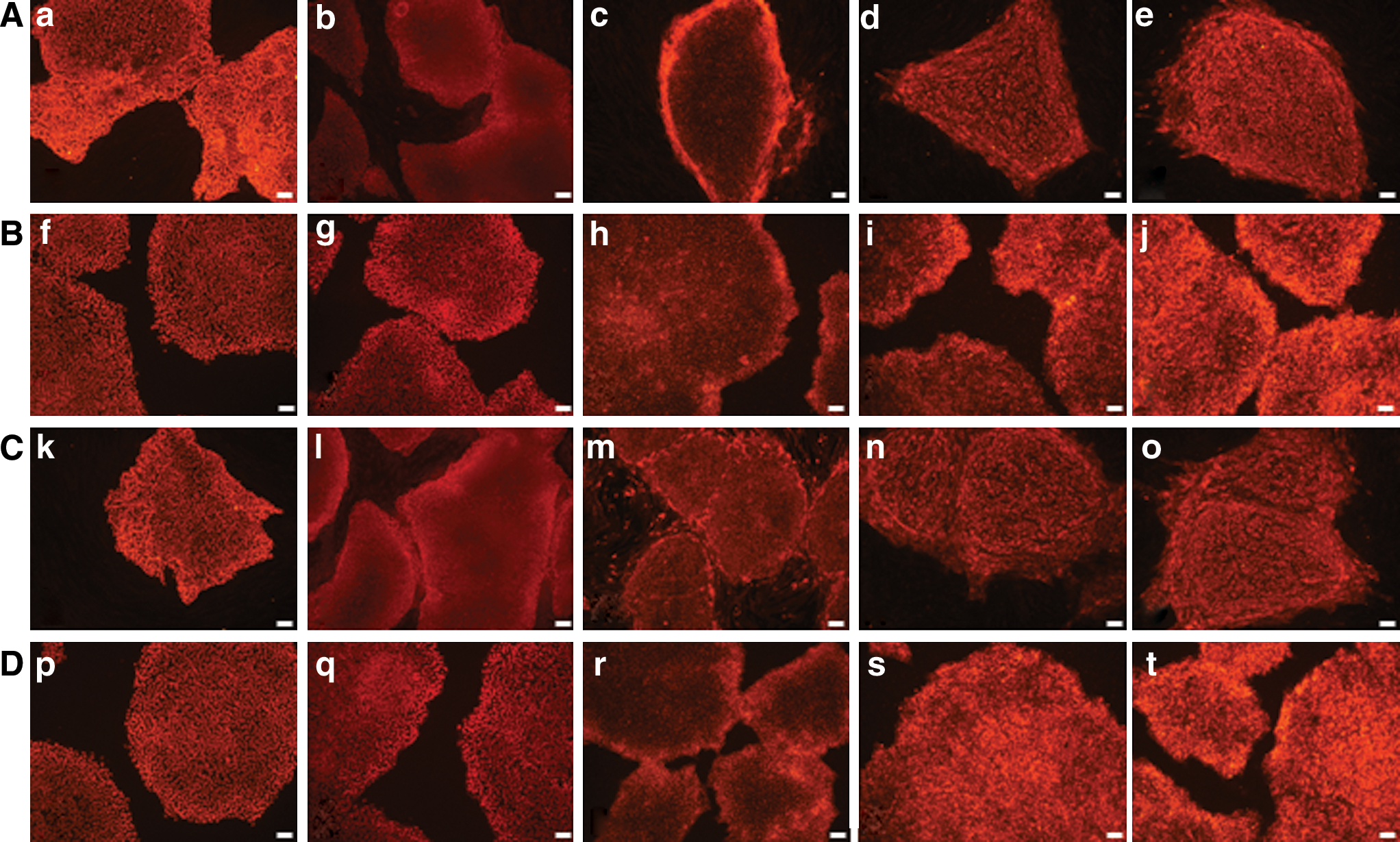
Pluripotency marker expression in hPSCs isolated from extremely differentiated colonies. hPSC clones isolated from extremely differentiated colonies:
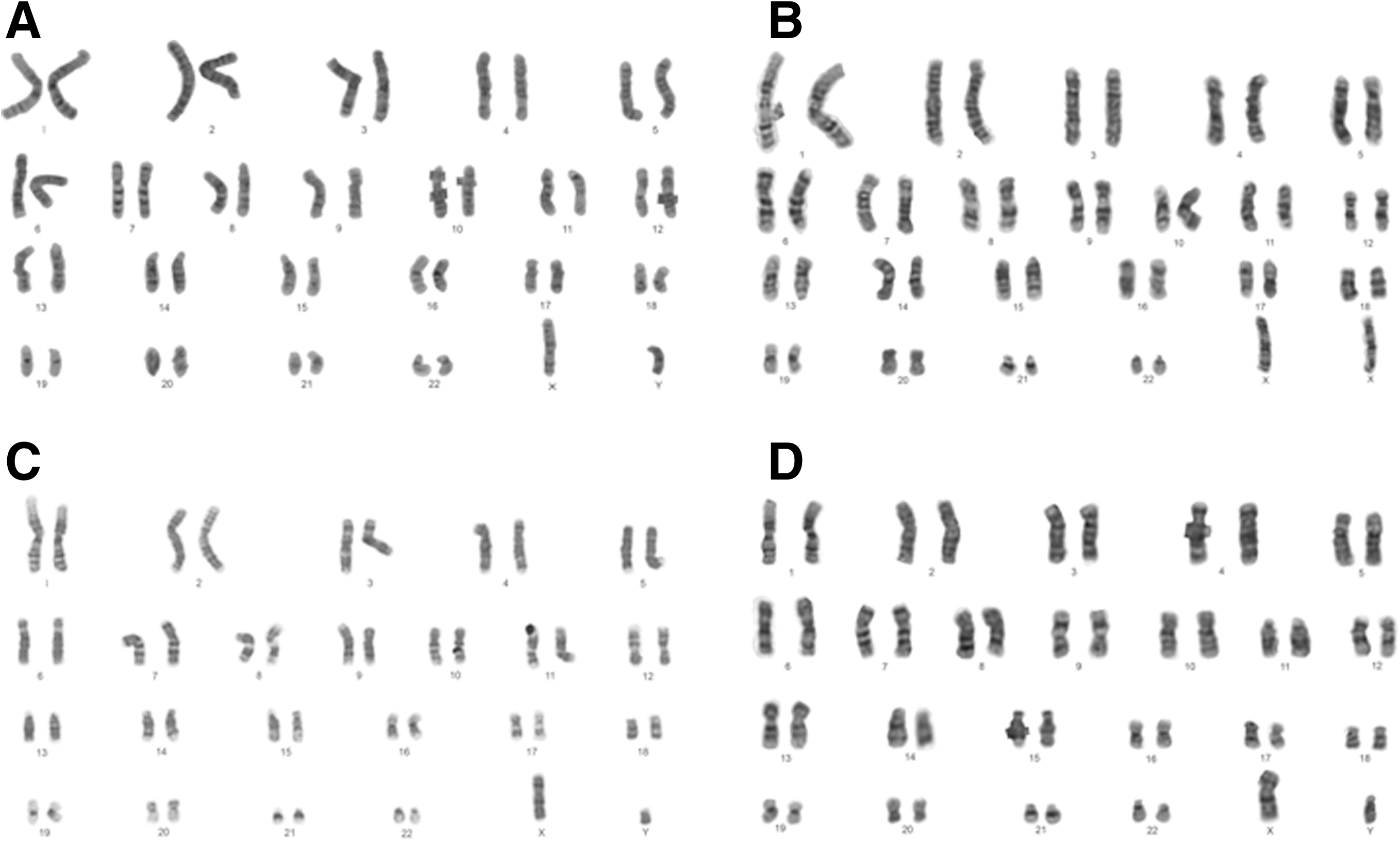
Karyotype analysis of hPSCs isolated from extremely differentiated colonies. Four sublines show normal karyotypes:
To characterize the differentiation ability of newly isolated hPSCs, we performed spontaneous in vitro differentiation via EB formation and analyzed these cells for markers of differentiation using immunofluorescence. Day 4 EBs were generated from hESC and hiPSC lines (Fig. 6A: a, e). After an additional 10 days of differentiation, immunofluorescence analysis revealed positive staining for β-tubulin III, smooth muscle actin, and α-fetoprotein, indicating the presence of ectoderm, mesoderm, and endoderm cells, respectively (Fig. 6A: b–d, f–h).
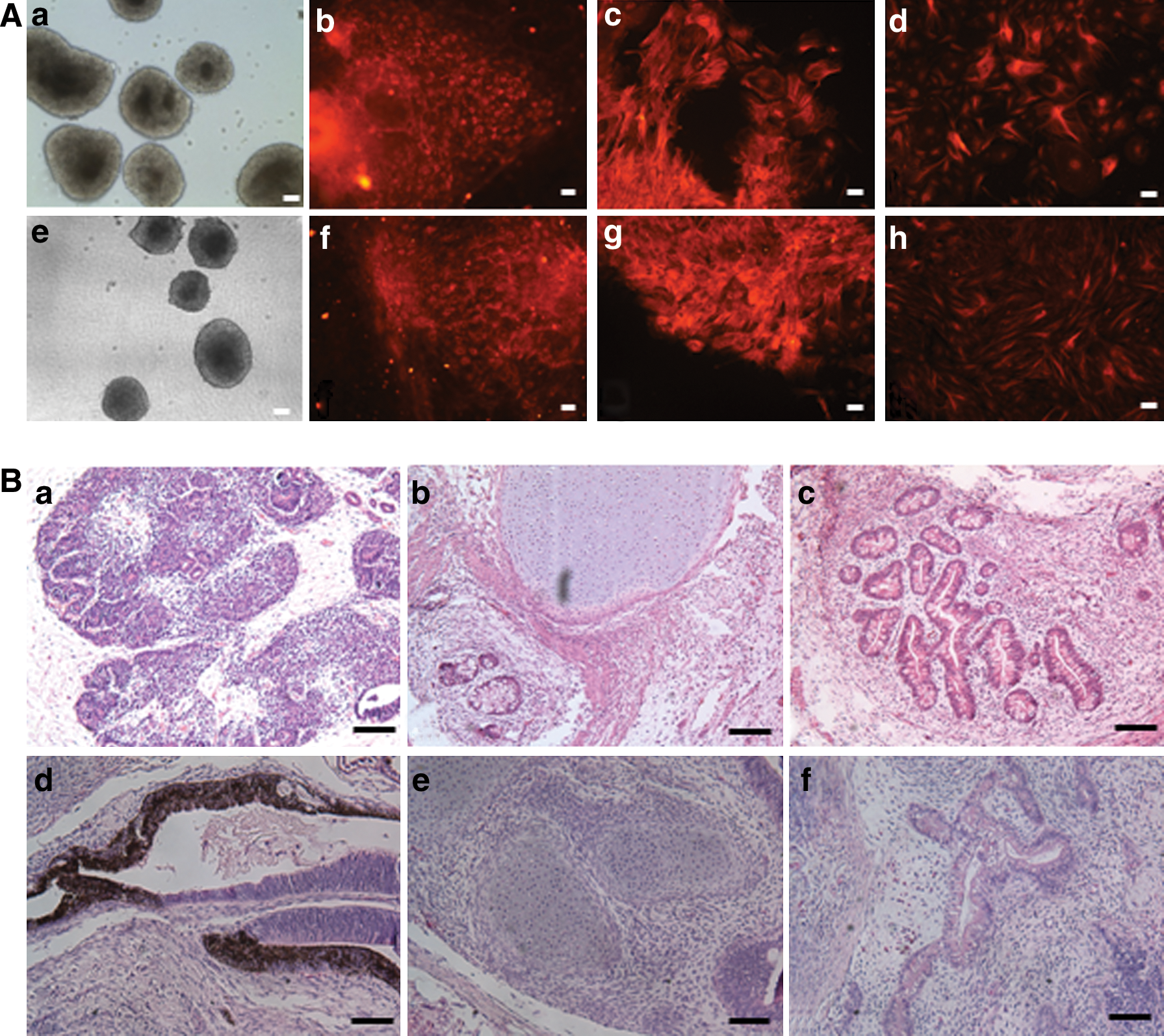
In vitro and in vivo differentiations of hPSCs isolated from extremely differentiated colonies.
We also used the teratoma assay as a test for the pluripotency of resulting hESCs and hiPSCs. For hESCs, rosettes of neural epithelium (ectoderm), cartilage (mesoderm), and gut-like epithelium (endoderm) were all observed (Fig. 6B: a–c). Teratoma derived from hiPSCs contained pigmented cells (ectoderm), cartilage (mesoderm), and columnar gland (endoderm) (Fig. 6B: d–f). These results indicated that the resulting cell lines were all pluripotent following their isolation from extremely differentiated colonies.
Discussion
We have found this method to be very useful for rescuing important clones, which are close to being lost. During our early experiments to derive hESC lines from cryopreserved human embryos [3], a bad batch of bFGF caused extreme differentiation in our newly isolated ESC colonies. Similarly, we have used this technique to rescue the rare colonies (Table 2, group 3), which form after DNA transformation or cellular reprogramming.
Both hESCs and hiPSCs are routinely cultured in both feeder-dependent and feeder-free culture systems, which can be suboptimal from time to time, because of variability of reagents. For feeder-dependent culture, mouse embryonic fibroblasts (MEFs) have been widely used as feeder cells to support hPSC growth. Because MEFs have obvious disadvantages, such as short life span in culture [30] and batch-to-batch variation [14,31], this can be a cause of extreme differentiation. Instead, human fibroblasts have become a popular feeder for the derivation and culture of hPSCs [7,32]. Knockout-serum replacement (KSR) is a defined, serum-free formulation optimized to grow and maintain undifferentiated hPSCs in feeder-dependent conditions. Not all batches of KSR are optimized to support hPSC growth. We have found that it is necessary to screen an optimal batch of KSR with feeder cells for optimal hPSC culture.
In some cases, if extreme differentiation occurs in one culture system, implying that it is suboptimal, the alternative culture system can be used to isolate undifferentiated hPSC cells. Although KSR medium can maintain hPSC growth on HFF feeder cells, we have found that this culture system (HFF feeder/KSR medium) is not suitable for the isolation of undifferentiated hPSCs from extremely differentiated colonies appearing in feeder-free conditions (ie, mTeSR1/Matrigel), as undifferentiated hPSCs seemed unhealthy after transferring them from feeder-free culture conditions to this feeder-dependent culture conditions (data not shown). However, another feeder-dependent culture system (HFFs/HEScGRO medium) worked well for isolating undifferentiated hPSCs.
Compared with feeder-dependent culture systems, the feeder-free culture system (mTeSR1/Matrigel) seemed to be quite stable. mTeSR1 and hESC-qualified Matrigel showed less batch-to-batch variability. In addition, this feeder-free culture system can provide a convenient, fast, and efficient culture environment to isolate undifferentiated hPSCs from colonies with extreme differentiation.
Undifferentiated hPSC colonies are easily dissociated into cell clumps after treatment with collagenase IV (feeder-dependent) or dispase (feeder-free). In contrast, it is difficult to dissociate differentiated colonies or differentiated parts of colonies into cell clumps because of tight junctions between differentiated cells. This means that undifferentiated cell areas are more prone to becoming dissociated from differentiated areas, which often retain their original colony sizes following treatment with collagenase IV or dispase.
Based on our experiments, we concluded that the ROCK inhibitor, Y27632, helped enormously in the rescue of extremely differentiated hESC and hiPSC colonies. Undifferentiated colonies are 2–3 times more readily formed in the presence of Y27632 than in its absence. Previous studies have indicated that Y27632 is safe and efficient at supporting hPSC expansion, and cryopreservation greatly improves hPSC adherence and viability [10,20,28,29]. Using Y27632, we successfully obtained completely undifferentiated hPSC colonies from all extremely differentiated single colonies or multicolonies in a single passage.
A suitable seeding density of cell clusters to culture dishes or wells is also crucial to the success of this method. Based on our experience, cell clumps from a single colony or multicolonies (3–4) with extreme differentiation should be seeded into a 1-cm well or 35-mm dish, respectively. Keeping newly formed cell colonies at a distance is helpful to remove differentiated colonies and pick undifferentiated colonies for further expansion.
To the best of our knowledge, this method has not been previously reported and fills an important technical gap in hPSC culture. Although companies have recently begun to promote the use of flow cytometry for isolating PSCs from differentiated colonies, such as BD's Human Pluripotent Stem Cell Sorting and Analysis Kit (
Footnotes
Author Disclosure Statement
No competing financial interests exist.