
Abstract
A number of transcriptional factors are required for pluripotency of stem cells. NANOG, a homeobox transcription factor, plays a critical role in regulating embryonic stem cell (ESC) pluripotency. The expression level of NANOG is tightly regulated, and perturbation in its expression level can lead to significant difference in the morphology, expression of cell surface markers, and growth factor dependence of human and mouse ESCs. Here, we demonstrate that the proteolysis of human NANOG is regulated by the ubiquitin-proteasomal pathway. The inhibition of proteasome activity by proteasome inhibitor MG132 showed increase in protein levels of endogenous NANOG in a dose-dependent manner in human ESCs (hESCS). We demonstrated that the inhibition of the proteasome activity and cotransfection with exogenous ubiquitin promotes endogenous ubiquitination of NANOG by coimmunoprecipitation assay. In addition, we showed that both K48- and K63-branched polyubiquitin chains can conjugate with NANOG in vivo. Moreover, NANOG was an unstable protein and exhibited relatively short half-life of about 120 min in hESCs. Pretreatment of hESCs with proteasome inhibitor MG132 inhibits NANOG protein degradation and extends its half-life. Finally, we found that a PEST motif sequence (rich in proline, glutamine, serine, and threonine) from amino acid 47 to 72 located toward the N-terminus of NANOG was shown to target the protein for degradation. Deletion of the PEST motif reduced ubiquitination of NANOG, leading to NANOG stabilization. Collectively, these results indicate that the expression level, stability, and activity of NANOG are modulated by post-translational mechanisms.
Introduction
E
The expression level of these transcriptional factors is crucial for determining the development of distinct cell fate of ESCs. Apart from transcriptional initiation and attenuation, expression of these transcriptional factors is also regulated post-transcriptionally at the level of mRNA stability, translation, and protein stability [16 –18]. Recently, NANOG is reported to undergo phosphorylation and shown that prolyl isomerase Pin1 interacts with phosphorylated Ser/Thr-Pro motifs of NANOG, leading to increased stability and activity of NANOG by inhibiting its ubiquitination [19]. However, these transcriptional factors, including Oct3/4, Klf4, and c-Myc, have relatively short half-life. The ubiquitin-proteasomal pathway (UPP) is responsible for the degradation of many short-lived regulatory proteins in vivo [20,21]. It is reported that these transcriptional factors of ESCs, including Oct3/4, c-Myc, Klf4, and NANOG, are proteolyzed through the UPP [16,17,19,22,23]. The process of ubiquitination is catalyzed by 3 enzymes: the ubiquitin-activating enzyme (E1), the ubiquitin-conjugating enzyme (E2), and the ubiquitin-protein ligase (E3) [24,25]. The E1 enzyme activates ubiquitin in an ATP-dependent reaction by forming a thiol ester bond with the COOH-terminal glycine of ubiquitin. The activated ubiquitin is then passed to an active cysteine site of an E2 enzyme. Finally, E3 enzyme recognizes E2 and facilitates the transfer of the activated ubiquitin from E2 to a lysine residue of a targeted protein through an isopeptide bond formation. After the formation of multiubiquitination on the protein substrate, target proteins are subjected to proteolysis by the 26S proteasome [24,25].
In this report, we demonstrated that the proteolysis and stability of NANOG protein is regulated by the UPP. We have also shown that NANOG undergoes both K48- and K63-branched polyubiquitination. Our studies illustrate that inhibition of proteasome activity extends the half-life of NANOG in human ESCs (hESCs). Further, we have identified a potential PEST motif sequence within NANOG and demonstrated its importance in the enhancement of NANOG ubiquitination and subsequently protein degradation.
Materials and Methods
Cell lines, tissue culture, and reagents
The hESC lines H9 (WiCell Research Institute, Madison, WI;
Generation of expression plasmids and transfections
The DNA fragment encoding full-length human NANOG in pcDNA3.1/myc expression vector was obtained from Prof. Jae-Hwan Kim (CHA University). The pEFIRES-HA-ubiquitin, pEFIRES-HA-ubiquitin-R48K, and pEFIRES-HA-ubiquitin-R63K constructs were obtained from Prof. Yossi Yarden (Weizmann Institute, Rehovot, Israel). PEST motif-deleted NANOG was generated and subcloned into pcDNA3.1-myc-tagged expression vector and labeled as NANOGΔPEST. Three micrograms of all the expression constructs were transfected using polyethyleneimine transfection reagent (Polysciences, Warrington, PA). After 48 h of transfection, the cells were harvested and lysed using a lysis buffer (1% Triton X, 150 mM NaCl, 50 mM Tris-HCl pH 8, and 1 mM phenylmethylsolfonyl fluoride (PMSF) for 20 min. Western blot analysis was carried out using an anti-myc antibody (9E10; Santa Cruz Biotechnology, Santa Cruz, CA) or anti-HA antibody (Santa Cruz Biotechnology).
Antibodies
Anti-NANOG (sc-30328), anti-ubiquitin (sc-8017), anti-myc (9E10) (sc-40), anti-HA (sc-7392), and anti-β-actin (sc-47778) antibodies were purchased from Santa Cruz Biotechnology).
Cell treatments with inhibitors
H9 and CHA15 hESC lines were treated with proteasome inhibitor MG132 at a concentration ranging from 0 to 10 μM. NCCIT cells and 293T cells at a density of ∼2.5×105 cells per 6 well dishes were transfected with expression plasmids. After 48 h of transfection, cells were treated with proteasome inhibitor MG132 at a concentration ranging from 0 to 50 μM.
In vivo ubiquitination assay
For in vivo ubiquitination assay, 3 μg of pcDNA3.1-myc-NANOG and pEFIRES-HA-ubiquitin were cotransfected into 293T cells. Cell lysates were immunoprecipitated with an anti-myc and blotted with an anti-HA antibody. For endogenous ubiquitination assay, NCCIT cell lysate was immunoprecipitated with an anti-NANOG antibody and immunoblotted with an anti-ubiquitin antibody. In addition, NCCIT cells were transfected with pEFIRES-HA-ubiquitin and immunoprecipitated using an anti-NANOG antibody and immunoblotted with an anti-HA antibody. 293T cells were used for the expression of the NANOG and cotransfected with pEFIRES-HA-ubiquitin R48K or pEFIRES-HA-ubiquitin R63K for identification of K48- or K63-branched NANOG ubiquitination. The cotransfected cell lysate was immunoprecipitated with an anti-myc antibody and blotted with an anti-HA antibody. For checking the intensity of ubiquitination, 293T cells were cotransfected with NANOG or NANOGΔPEST with pEFIRES-HA-ubiquitin and performed immunoprecipitation assay with an anti-myc antibody and blotted with an anti-HA antibody. After 48 h of transfection, the cells were lysed in a lysis buffer (1% Triton X-100, 150 mM NaCl, 50 mM Tris-HCl pH 8, 1 mM PMSF) for 20 min. Cell lysates were centrifuged at 13,000 rpm at 4°C, and supernatant was collected into a fresh Eppendorf tube. The cell lysates were incubated with respective antibodies overnight at 4°C. Twenty microliters of protein A/G Sepharose (Santa Cruz Biotechnology) was added to the lysate and incubated at 4°C for 1 hr. Beads were washed with lysis buffer for 3 times, elution was performed with 2×sodium dodecyl sulfate loading dye, and boiled for 5 min. The eluted samples were loaded onto 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel followed by western blotting.
PEST motif sequence analysis of NANOG
The epestfind server (
Stability assays
H9, CHA15 hESC lines and NCCIT cells were treated with CHX (100 μg/mL). 293T cells were transfected with either pcDNA3.1-NANOG or pcDNA3.1-NANOGΔPEST. Forty-eight hours after the transfection, cells were treated with CHX (100 μg/mL), and cells were subsequently harvested at different time intervals. Equal amount of proteins from each treated sample was analyzed by western blot.
Results
Upregulation of endogenous NANOG protein by the proteasome inhibitor MG132
One of the mechanisms to regulate the total turnover of many proteins is by the UPP. To examine whether NANOG protein is regulated by the UPP, we analyzed the expression level of NANOG protein in the presence of proteasomal inhibitor MG132. If NANOG is degraded through the UPP, inhibition of proteasomal activity could increase the level of NANOG protein due to the accumulation of ubiquitinated NANOG. Thus, hESCs were treated with different concentration of proteasome inhibitor MG132 for 12 h. The pluripotent hESCs showed sensitivity to proteasome inhibitor MG132 at concentrations above 10 μM, leading to morphological changes. In contrast, the morphology of NCCIT and 293T cells was not affected by MG132 even at 50 μM. Our present observation may be justified in the light of previous reports that have shown similar cytotoxic effect of MG132 on hESCs [28]. Therefore, we investigated the effect of MG132 on pluripotent hESCs at concentration up to 10 μM. MG132 was added with a concentration-dependent manner to the culture media of H9 and CHA15 hESCs, and investigated the expression level of NANOG protein. Western blot analysis showed that the endogenous NANOG protein was increased when hESCs were treated with MG132 (Fig. 1A, B lanes 2 and 3). Further, we checked the effect of MG132 treatment on endogenous level of NANOG in human embryonic carcinoma cell line NCCIT, which did not show any morphological change with MG132 treatment. For this purpose, we treated NCCIT cells with MG132 with a concentration-dependent manner for 12 h and the total NANOG was immunoprecipitated and blotted with an anti-NANOG antibody. The endogenous level of NANOG protein is increased along with the increasing concentration of MG132 (Fig. 1C). Additionally, we tested the effect of proteasome inhibitor on 293T cells transfected with constant amount of myc tagged-NANOG. After 48 h of transfection, cells were incubated with increasing concentration of MG132 for 6 h and analyzed by western blot analysis. Treatment with the increasing concentration of MG132 resulted in dose-dependent accumulation of NANOG in 293T cells (Fig. 1D). These results suggest that the UPP might be involved in the regulation of NANOG protein turnover.
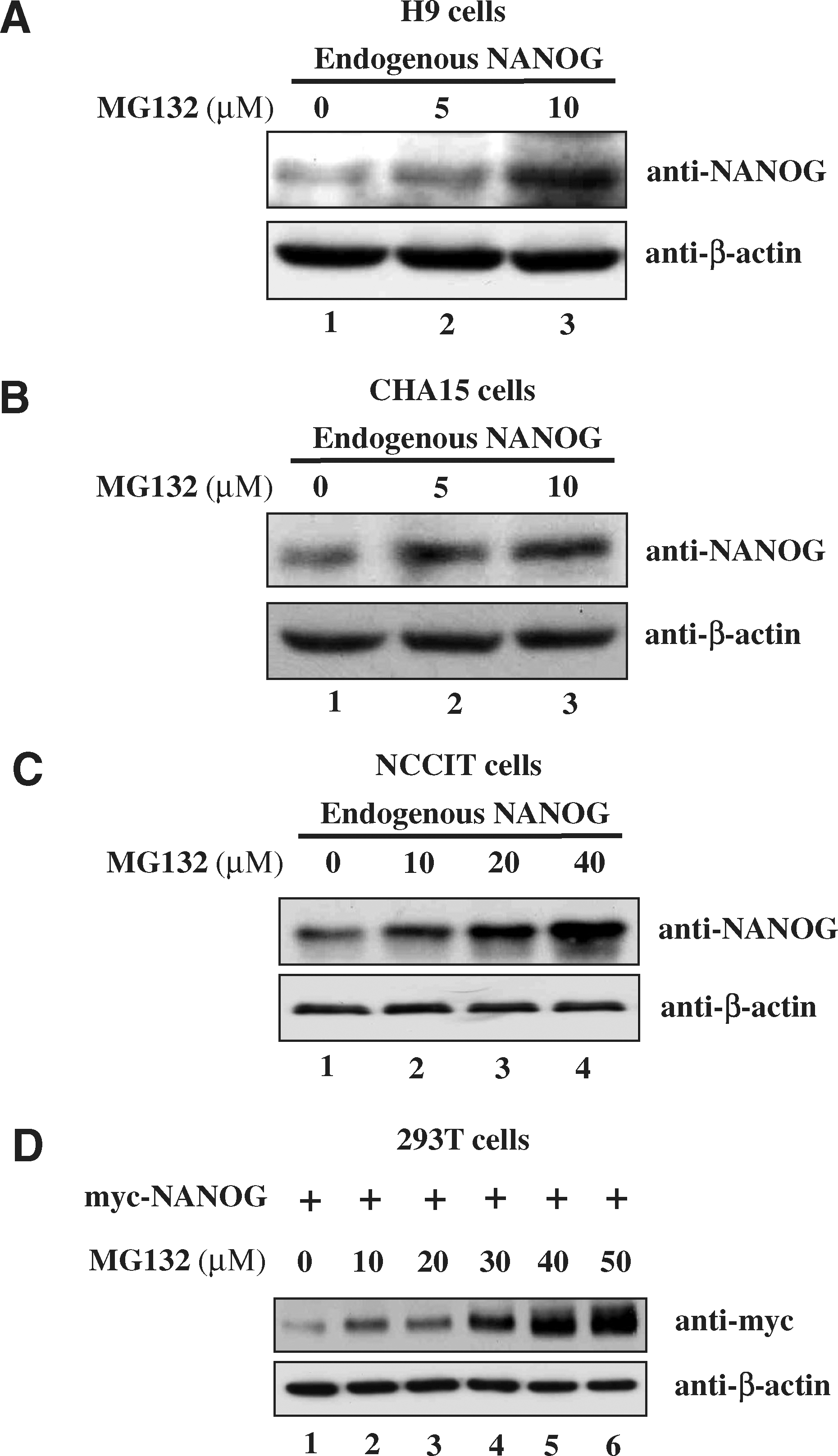
Protein levels of NANOG in human embryonic stem cells (hESCs) are regulated by the ubiquitin-proteasomal pathway.
Proteolysis of NANOG by the UPP
To further investigate whether NANOG protein turnover is regulated by the UPP, we checked the interaction between ubiquitin and NANOG in 293T cells. For this purpose, 293T cells were transfected with a pcDNA3-myc expression vector containing NANOG alone, or together with HA-ubiquitin plasmid. Immunoprecipitation was carried out with an anti-myc antibody, followed by western blot with an anti-HA antibody. A typical high-molecular-weight smear bands of ubiquitin molecules were detected in cells cotransfected with myc-NANOG and HA-ubiquitin, but not in cells transfected with myc-NANOG alone (Fig. 2A lane 4). Further, we demonstrated endogenous ubiquitination of NANOG in NCCIT cells by treating with MG132 (40 μM) for 12 and 24 h and immunoprecipitated with an anti-NANOG antibody followed by western blot analysis using an anti-ubiquitin antibody. The increased levels of high-molecular-weight smear were observed in NCCIT cells treated with MG132 (Fig. 2B). In addition, we examined whether NANOG could be ubiquitinated by transfecting exogenous HA-ubiquitin into NCCIT cells and immunoprecipitation of ubiquitin–protein conjugates was detected using transfected cell lysate treated with or without MG132. Western blot analysis revealed that the accumulation of ubiquitinated NANOG was detected more in ubiquitin transfected MG132-treated cells (Fig. 2C). These results demonstrated that NANOG interacts with ubiquitin and undergoes polyubiquitination through the UPP. Therefore, we carried out our further investigation to identify the potential motifs or sequences responsible for the rapid degradation of NANOG by the UPP.
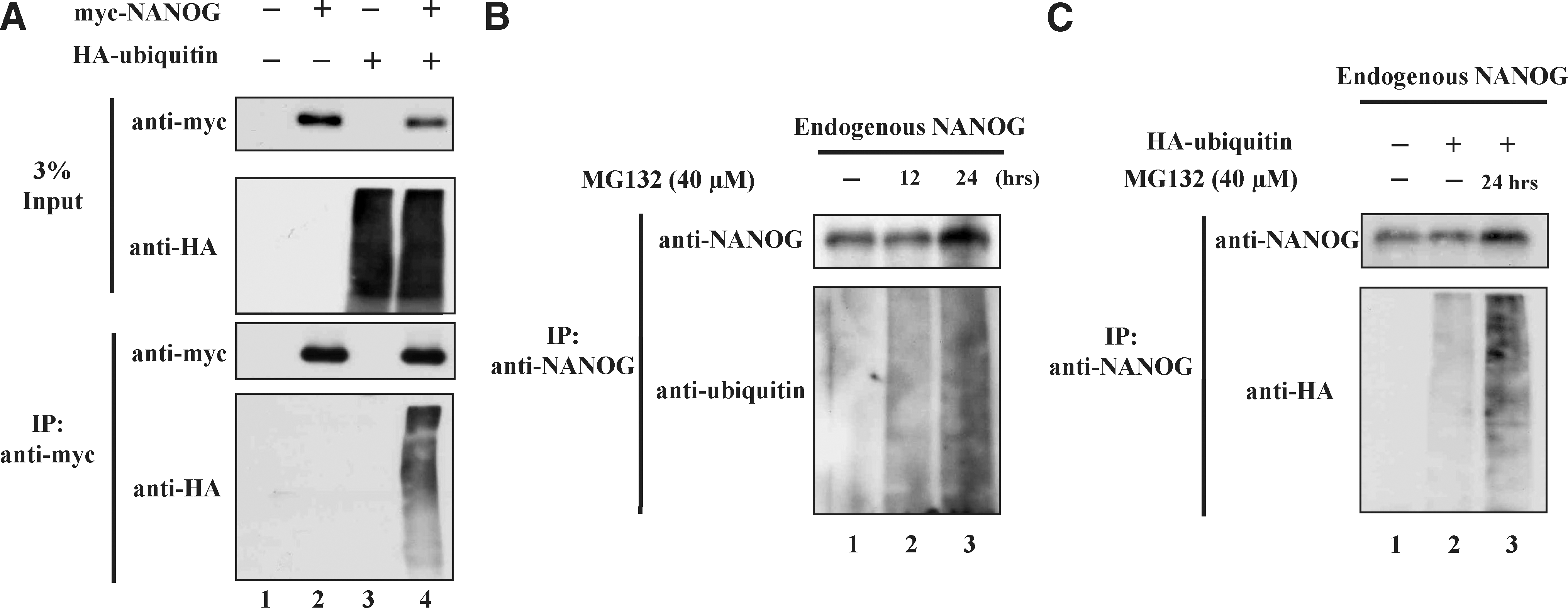
In vivo ubiquitination of NANOG. 293T cells were transfected with pcDNA3-myc-NANOG and pEFIRES-HA-ubiquitin separately and together.
NANOG is subjected to both K48- and K63-linked polyubiquitination
The ubiquitin chains may exist in different forms based on the lysine (K) site used for these interubiquitin linkages. K6, K11, K29, K48, and K63 are the several lysine sites utilized for the ubiquitin chain formation. Among them, polyubiquitin chains formed through K48 and K63 are very well studied. Therefore, we examined the type of ubiquitin chains that are formed on the NANOG by using 2 mutant ubiquitin constructs such as ubiquitin-R48K and -R63K in which all the lysines are substituted to arginine except at K48 or K63, so that only K48- or K63-branched ubiquitin chains could be conjugated to NANOG upon cotransfection. Immunoprecipitation assay followed by western blot analysis showed high-molecular-weight smear of ubiquitin molecules in the cells cotransfected with myc-NANOG and HA-ubiquitin-R48K or HA-ubiquitin-R63K (Fig. 3 lanes 5 and 6). This result suggests that both K48- and K63-branched ubiquitin chains could be attached to NANOG.
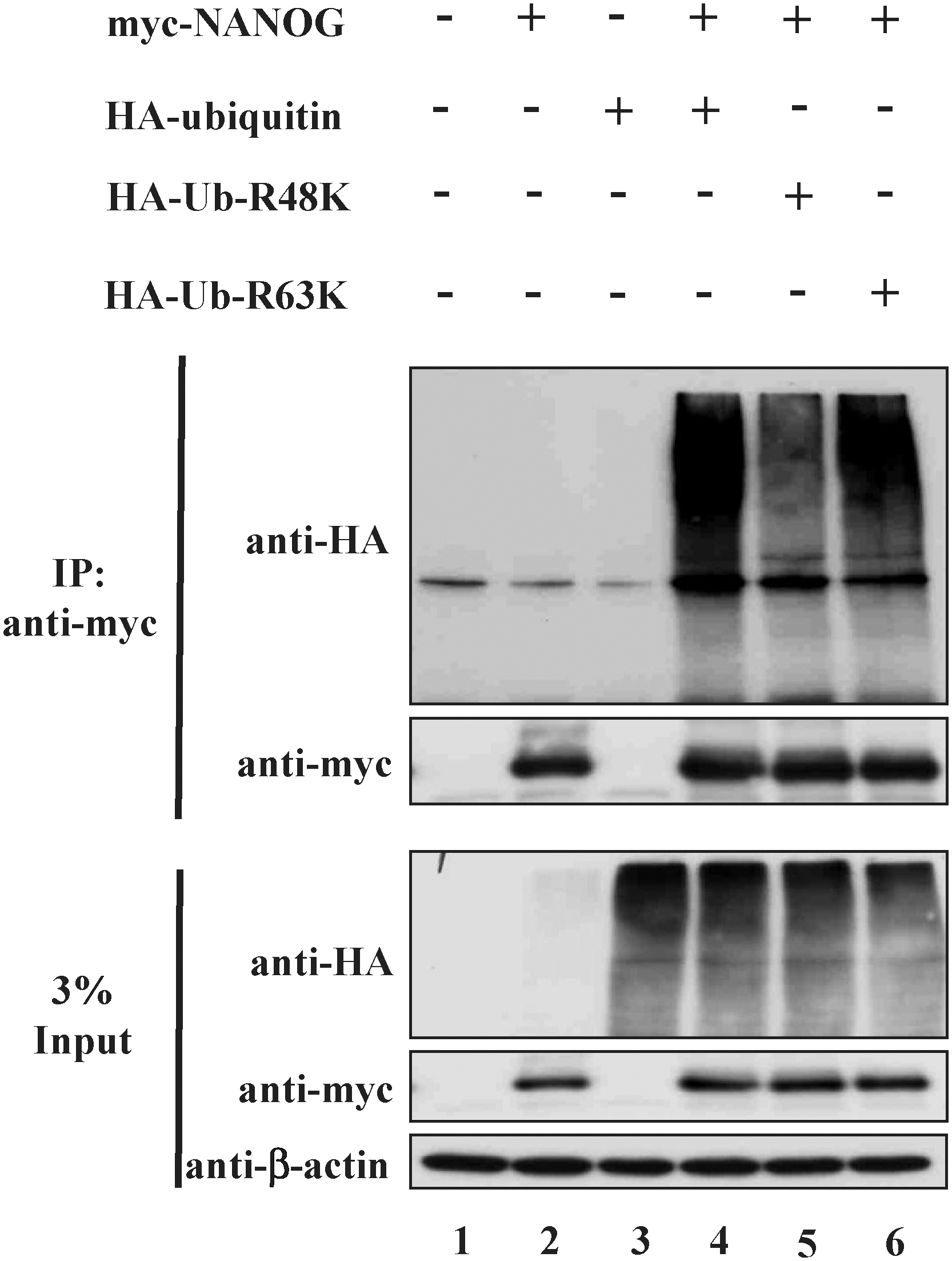
NANOG is subject to both K48- and K63-linked polyubiquitination. 293T cells were transfected with pcDNA3-myc-NANOG, pEFIRES-HA-ubiquitin, pEFIRES-HA-ubiquitin-R48K, and pEFIRES-HA-ubiquitin-R63K. Ubiquitination of NANOG was confirmed by coimmunoprecipitation with an anti-myc antibody and immunoblotted with an anti-HA antibody. Each experiment was repeated at least 3 times.
NANOG exhibits a relatively short half-life
To access the stability of endogenous NANOG, we treated H9 and CHA15 hESCs with CHX (100 μg/mL), which inhibits new protein synthesis. The cells treated with CHX were harvested at 0, 60, 120, 180, and 240 min and analyzed for endogenous NANOG. Western blot analysis showed that the protein level of NANOG was reduced significantly within 4 h of CHX treatment (Fig. 4A, B). NANOG exhibited a relatively short half-life about 120 min in the presence of CHX in hESCs. Similar results were obtained on NCCIT cells treated with CHX (Fig. 4C).
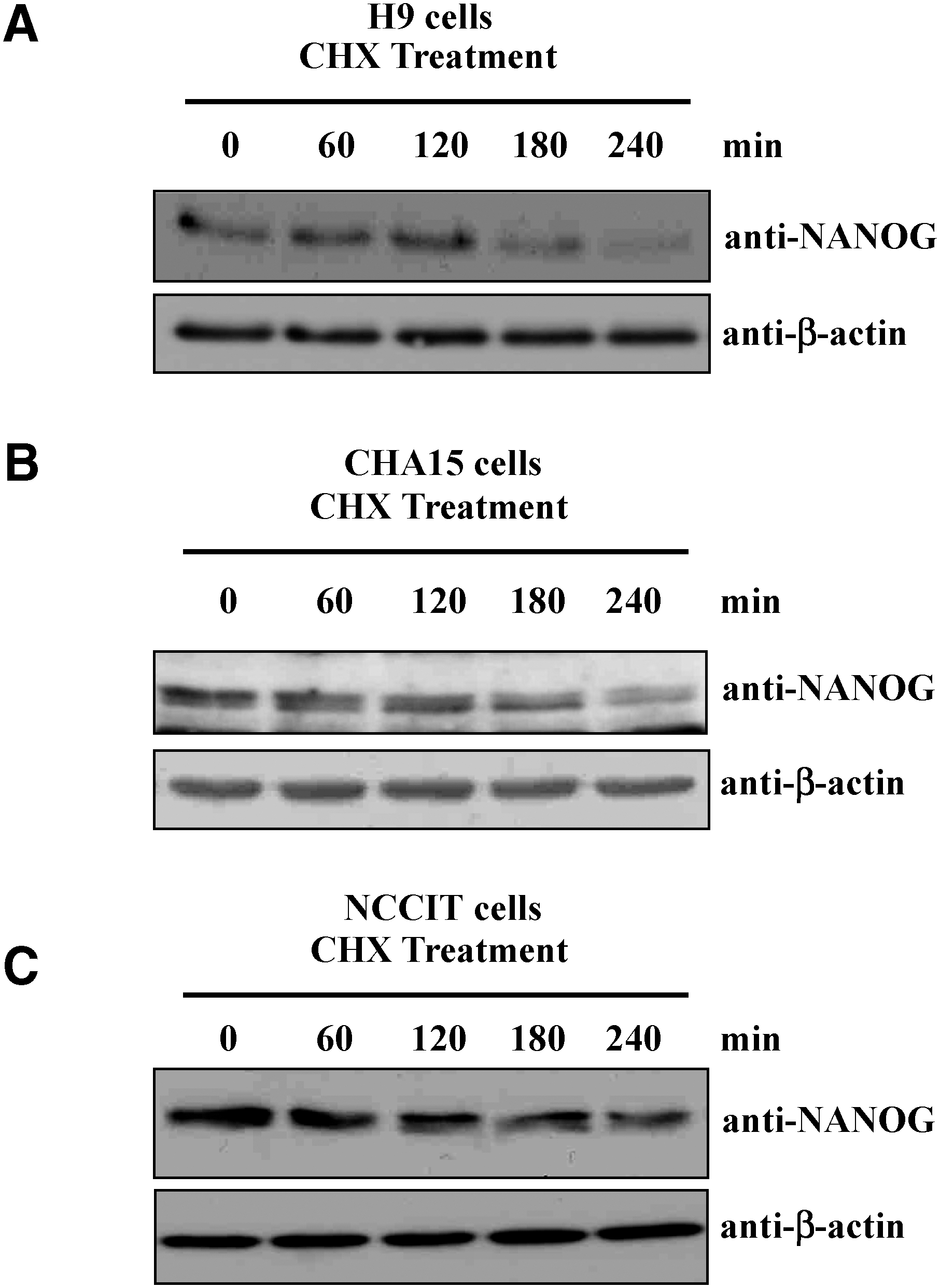
Half-life of endogenous NANOG.
Half-life of NANOG is altered by proteasome inhibitor MG132
We further accessed the stability of endogenous NANOG in H9 and CHA15 hESCs in the presence of proteasome inhibitor. hESCs were pretreated with MG132 (10 μM) for 12 h just before incubating with CHX (100 μg/mL). The cells treated with CHX were harvested at 0, 60, 120, 180, and 240 min in the presence or absence of MG132 and analyzed for endogenous NANOG by western blot analysis. The half-life of NANOG was shown to extend in hESCs pretreated with proteasome inhibitor MG132 (Figs. 5A, B). The same blots were stripped and checked for β-actin to confirm the equal loading of samples. The effect of MG132 on half-life of NANOG was verified in 293T cells transfected with constant amount of myc-NANOG and pretreated with MG132 (50 μg/mL) before incubating with CHX reagent. When 293T cells were pretreated with MG132, CHX degradation of NANOG was inhibited (Fig. 5C). The same blots were stripped and checked for β-actin to confirm the equal loading of samples. These results suggest that NANOG is an unstable protein, and its half-life is regulated by the UPP.
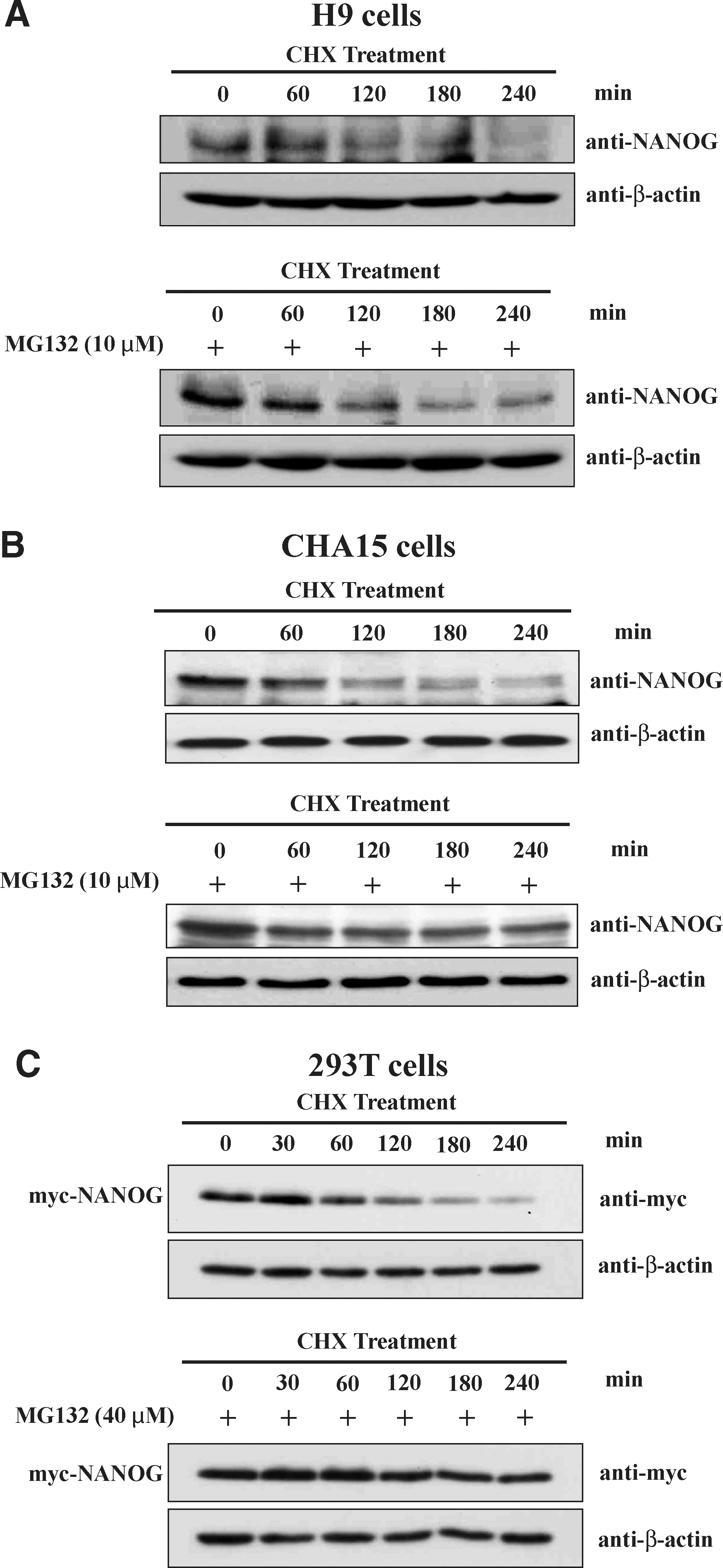
Proteasome inhibitor MG132 extends the half-life of both endogenous and exogenous NANOG.
PEST motif of NANOG signals for the rapid proteolysis and ubiquitination of NANOG
PEST motif sequence is reported in many short-lived proteins and predicted to target proteins for rapid degradation through the UPP [29,30]. We have previously shown that USP36 containing PEST motif undergoes rapid degradation via the UPP [29]. Therefore, bioinformatics tools were used to identify PEST motif sequence within NANOG. The epestfind program revealed that NANOG has a potential PEST sequence having a PEST score 10.34, which consists of 26 amino acids starting from 47 to 72 at its N-terminal region (Fig. 6A). We next hypothesized that the deletion of PEST motifs from NANOG sequence might enhance the stability of NANOG protein by reducing its protein degradation. To examine this possibility, we created a PEST motif-deleted NANOG construct (Fig. 6B) and performed ubiquitination assay along with NANOG to check its ubiquitination status in 293T cells. We cotransfected myc-NANOG or myc-NANOGΔPEST with HA-ubiquitin into 293T cells and exogenous NANOG proteins were immunoprecipitated. Western blotting analysis with an anti-HA antibody showed reduced high-molecular-weight ubiquitin smear bands conjugated with NANOGΔPEST (Fig. 6C, lane 6) when compared to NANOG (Fig. 6C, lane 5).
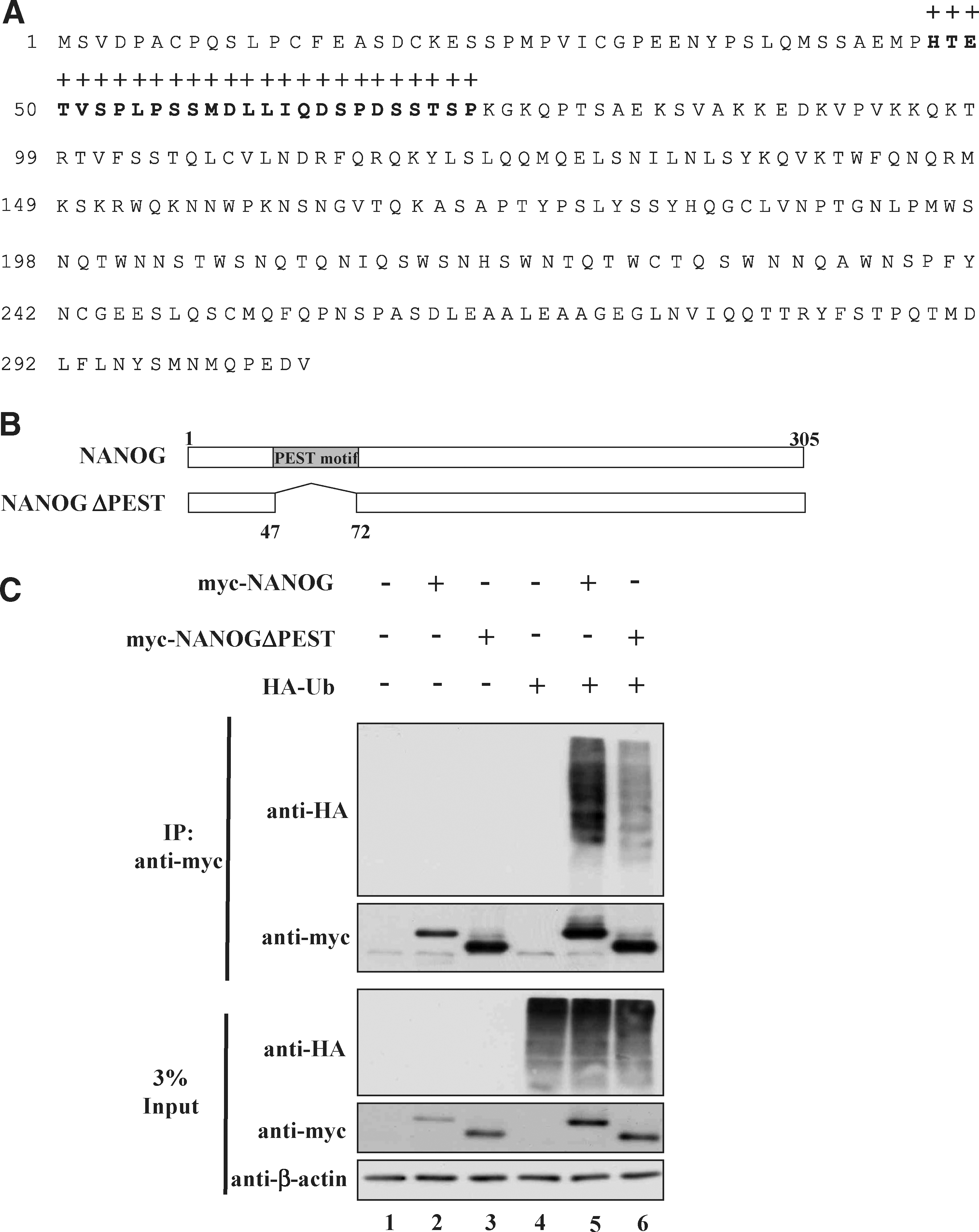
NANOG-containing PEST motif sequence enhances its ubiquitination.
NANOG containing PEST motif sequence regulate its protein stability
We further analyzed the stability of NANOG and PEST motif-deleted NANOG by transfecting constant amount of both the plasmids into 293T cells and treated with CHX after 48 h of transfection. The NANOGΔPEST showed partial stabilization of protein when compared with the NANOG (Fig. 7). These results suggest that a PEST motif sequence within the sequence of NANOG protein is required for an efficient NANOG ubiquitination and the deletion of this PEST motif results in increased stability of NANOG protein by suppressing its ubiquitination.
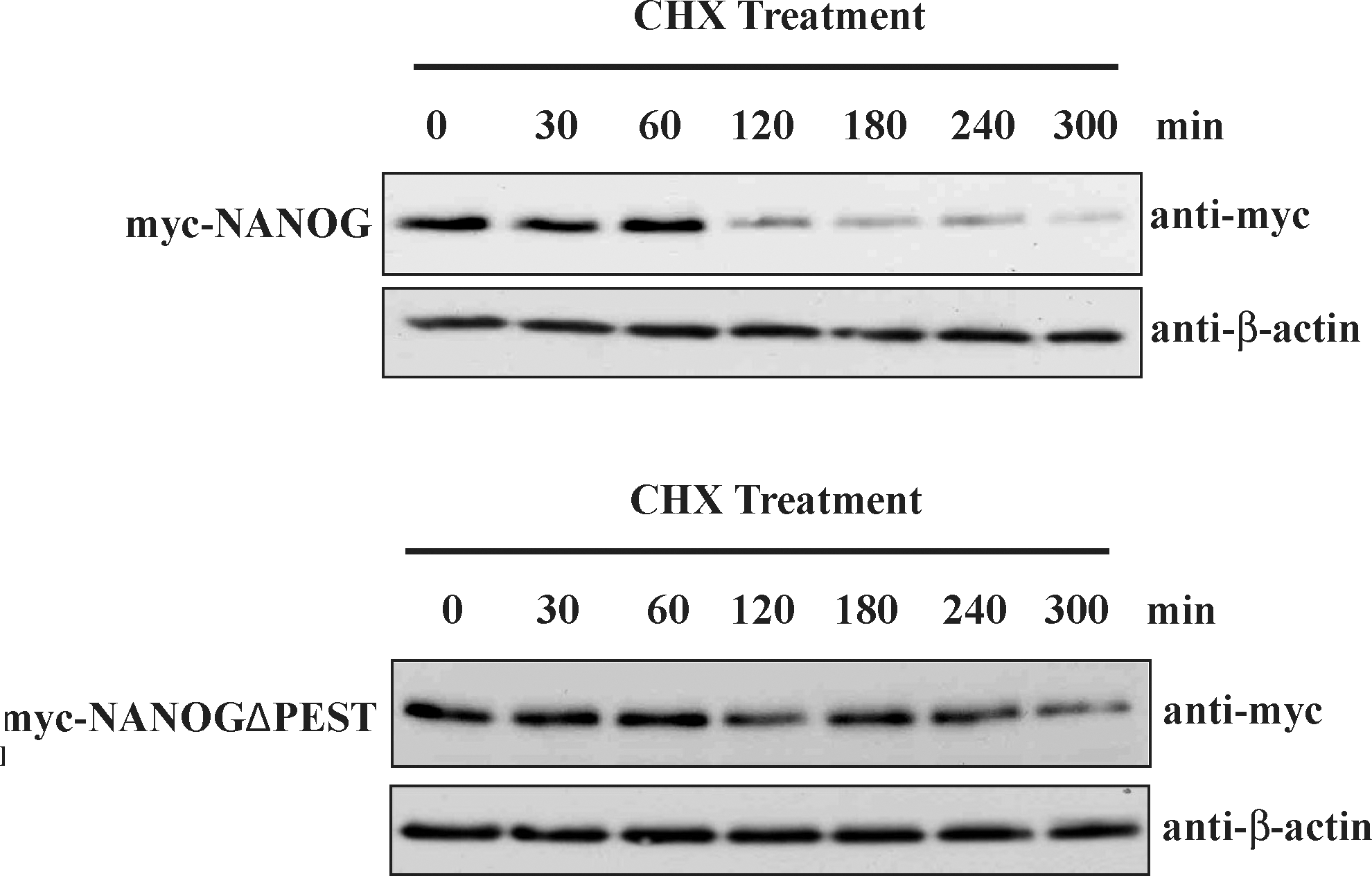
NANOG-containing PEST motif sequence regulates its protein stability. 293T cells were transfected with equal amount (1 μg) of myc-NANOG or myc-NANOGΔPEST. Forty-eight hours after transfection, cells were incubated with CHX (100 μg/mL) for the indicated time points and cells were analyzed by western blot analysis. The same blots were stripped and reprobed with an anti-β-actin antibody to confirm equal loading of the samples. At least 3 individual experiments were performed in each study.
Discussion
The expression level of the transcription factors such as Oct3/4, c-Myc, NANOG, Klf4, and SOX2 is very crucial in regulating the fate of pluripotent stem cells and germ cell precursors. A small change in the expression level of these transcriptional factors leads to dramatic effect on differentiation or oncogenesis [31 –34]. These transcriptional factors exhibit relatively short half-lives. In general, the UPP is the main cause for the degradation of short-lived regulatory proteins [20,21]. Indeed, Klf4, c-Myc, and Oct3/4 undergo ubiquitin-mediated proteolysis through 26S proteasome pathway.
We initiated our investigation to check whether NANOG, a master gene of ESC pluripotency, is regulated by 26S proteasome pathway. We checked the expression level of transfected NANOG in the presence of proteasome inhibitors to confirm the role of proteasome in NANOG protein degradation. The expression level of NANOG was significantly increased along with the increasing concentrations of MG132. Recently, Szutorisz et al. have investigated the effect of chemical inhibition of proteasome activity on transcription of a tissue-specific locus in mouse ESCs. Treatment with the proteasome inhibitor MG132 showed increase in the level of intergenic transcription in a tissue-specific locus in ESCs. It was reported that proteasome inhibition could trigger the activation of promoters, in turn, inducing intergenic transcription in a tissue-specific locus in ESCs. Treatment with MG132 up to a concentration of 20 μM does not affect cell survival and cell morphology of ESCs [35]. In contrast, Assou et al. have reported great sensitivity of ESCs upon treatment with proteasome inhibitor MG132. The morphology of hESC colonies were changed to large patches of differentiation even at a low concentration of MG132 (500 nM), while the morphology of hES-differentiated fibroblast HD90 or HS181 cells were not affected by MG132 even at a high concentration. Further, RT-PCR analysis on ESCs treated with MG132 showed decrease in the expression level of OCT3/4, SOX2, and NANOG [28]. In line with these results, we also observed similar changes in the morphology of hESCs treated with MG132. In our study, the changes in morphology of hESCs were observed at concentrations above 10 μM MG132 treatment. Beyond this concentration, the ESC colonies were transformed into large patches of differentiation. In contrast, cells such as NCCIT and 293T transfected with MG132 at a concentration up to 50 μM were found to be unaffected. In our study, the concentration of MG132 treatment on hESCs was limited to 10 μM and we further investigated its effect on translational level of NANOG in ESCs. Our results suggest that inhibition of proteasome activity increases the translational level of NANOG in hESCs.
In addition, we confirmed by coimmunoprecipitation assay that the NANOG interacts with ubiquitin molecules and undergoes polyubiquitination. Our results suggest that NANOG is degraded by the UPP, which was similar to the earlier report [19]. Although there are several lysine sites such as lysine (K) 6, 11, 29, 48, and 63 that are utilized for chain formation, only K48- and K63-linked chains are well studied. K48-linked chains are responsible for targeting proteins to proteasomal degradation, whereas K63-linked chains results in diverse functions by modifying the structure of target proteins, localization or activity [36]. Therefore, we wished to check whether NANOG undergoes K48- or K63-linked polyubiquitin chains. Our observations were confirmed by coimmunoprecipitation assay that both K48- and K63-linked polyubiquitin chains could conjugate on the NANOG protein.
In the present work, we also show that the endogenous NANOG protein is an unstable protein having relatively short half-life. NANOG exhibited short half-life of about ∼120 min, when the ESCs were treated with the CHX reagent, which inhibits new protein synthesis. In contrast, pretreatment of cells with MG132 led to significant increase in the stability of NANOG, suggesting that the UPP is responsible for the total turnover of the NANOG protein. Recently, we and others have reported that the rapid turnover of proteins is regulated by several PEST motif signals that could induce the protein degradation [29,30]. Here, we demonstrated that NANOG contains PEST motif sequence signals for ubiquitination and subsequent protein degradation. Recently, Moretto-Zita et al. have reported that the 4 potential Ser/Thr-Pro motifs that are phosphorylated in NANOG promotes the interaction between NANOG and prolyl isomerase Pin1, thus resulting in NANOG stabilization by inhibition of its ubiquitination. NANOG with mutated phosphorylation sites cannot interact with Pin1, causing impaired self-renewal and decreased stability of NANOG protein. Thus, Pin1 activity was shown to be critical for stabilizing NANOG protein by suppressing its ubiquitination in ESCs [19]. Our investigation revealed that 3 of the 4 Ser/Thr-Pro phosphorylation sites (as reported by Moretto-Zita et al.) that are responsible for binding with Pin1 lie within the PEST motif of NANOG. PEST motif sequences have been suggested to function as a degradation signal [37,38] and several reports have showed the importance of PEST motifs located at amino or carboxyl terminus as required for tracking proteins into the UPP [39 –42]. The short-lived proteins containing these PEST motifs that are enriched in proline (P), glutamic acid (E), serine (S), threonine (T), and aspartic acid (D) residues would destabilize any secondary structure and forms disordered loops in the globular proteins that are often preferred targets for ubiquitin-proteasome dependent and independent protein degradation [37]. Thus, the deletion of PEST motif sequence in NANOG might greatly reduce the inherent susceptibility for proteolysis resulting in NANOG stabilization. It was also reported that NANOG is expressed discontinuously in pluripotent cells in the embryo [1]. In this context, one can speculate that the fluctuating expression of NANOG in mouse ESCs might be due to regulation by its post-translational modifications such as phosphorylation and ubiquitination during pluripotency. Taken together, these results support that NANOG is regulated by the UPP and the N-terminus of NANOG-containing PEST motif sequence is responsible for the stability and degradation of NANOG.
Footnotes
Acknowledgments
We would like to thank Professor Jae-Hwan Kim (CHA University) for providing us pcDNA3-myc-NANOG and Professor Yossi Yarden (Weizmann Institute) for pEFIRES-HA-ubiquitin, pEFIRES-HA-ubiquitin-K63R, and pEFIRES-HA-ubiquitin-K48R constructs. We wish to thank Sai Kiran Sharma at the University of Alberta, Canada for his critical comments on our article. We also thank Hey-Jin Lee and Jung-Mi Park for their initial assistance for this project. This research was supported by a grant (SC 5130) from Stem Cell Research Center of the 21st Century Frontier Research Program funded by the Ministry of Education, Science and Technology, Republic of Korea.
Author Disclosure Statement
No competing financial interests exist.