
Abstract
Understanding the normal development of individual neural subtypes provides an essential framework for the design of rational approaches to embryonic stem cell differentiation for in vitro studies and cell replacement therapies. Of particular interest and a particular challenge are the cells that build-up the telencephalon. Recent research has unraveled key developmental mechanisms contributing to the generation of specific telencephalic cells. We focus on morphogens and transcription factors known to regulate distinct developmental processes. These include early anterior/posterior patterning, dorsal/ventral patterning, and generation of progenitor domains and neuronal specification into major classes of telencephalic cells: glutamatergic projection neurons, different subtypes of γ-aminobutyric acid-ergic interneurons and projection neurons, as well as cholinergic interneurons and projection neurons. Based on a comparison with in vivo telencephalic neurogenesis, we propose that the specific combinations of transcription factors expressed during development can serve as milestones for the in vitro differentiation of embryonic stem cells toward specific telencephalic neurons.
Overview on Telencephalic Neuronal Cell Diversity and Developmental Mechanisms
A
At the cellular level the telencephalon is populated by a large diversity of neurons. The neuronal cell types and their progenitor domains in mouse embryonic telencephalon are shown in Fig. 1.
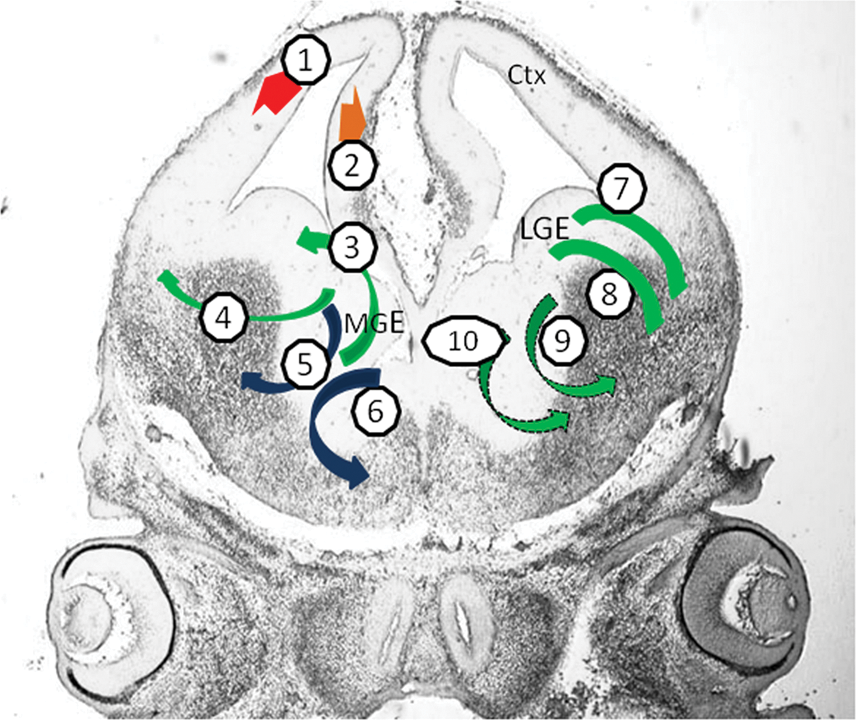
Telencephalic neuronal cell diversity. Glutamatergic projection (pyramidal) neurons in neocortex (1, red arrow) and hippocampus (2, orange arrow) are generated in the dorsal telencephalon. GABA-ergic neurons (green arrows) and cholinergic neurons (blue arrows) originate from the medial, lateral and caudal ganlionic eminences (MGE, LGE and CGE) in the ventral telencephalon. MGE generates different subclasses of GABA-ergic and cholinergic interneurons as well as cholinergic projection neurons (3–6): somatostatin (3) and parvalbumin (4) subclasses of GABA-ergic interneurons, from ventral and dorsal MGE, respectively; cholinergic interneurons that populate the striatum (5) and cholinergic neurons that project mainly to the cortex and hippocampus (6). Calretinin expressing GABA-ergic interneurons of the olfactory bulb (7) and GABA-ergic projection neurons that populate that striatum (8) arise from LGE. Calretinin (9) and/or vasoactive intestinal peptide (10) subclasses of GABA-ergic cortical interneurons (dotted arrows) arise from CGE (posterior plane, not shown). Coronal section, embryonic day (E) 13 mouse brain, showing the cortical primoridium (Ctx) and the MGE and LGE. Color images available online at
Glutamatergic projection (pyramidal) neurons represent the majority of cortical neurons, are generated in the dorsal telencephalon, and include many subtypes. Each subtype is characterized by a specific combination of laminar position, morphology, marker expression, and connectivity pattern [4,10,11]. γ-aminobutyric acid (GABA)-ergic cortical interneurons originate in the ventral telencephalon and subsequently migrate dorsally into the developing cortex. Other cell types originating from the ventral telencephalon include GABA-ergic and cholinergic interneurons and GABA-ergic and cholinergic projection neurons that populate the striatum, pallidum, and other forebrain ventral regions.
Different subclasses of GABA-ergic interneurons arise from different progenitor domains in the ganglionic eminences:
Somatostatin subclasses of GABA-ergic interneurons, as well as a population of neuropeptide Y-expressing interneurons that ultimately reside in the cortex and the basal ganglia are generated in ventral MGE [9,12].
Parvalbumin subclasses of GABA-ergic interneurons, constituting the majority of the cortical interneurons, are generated in dorsal MGE [9,12].
Calretinin-expressing and/or vasoactive intestinal peptide-expressing GABA-ergic cortical interneurons are produced primarily in CGE [13,14]. Calretinin-expressing GABA-ergic interneurons, which ultimately reside in the olfactory bulb, arise from the dorsal LGE [15].
GABA-ergic projection neurons, such as the medium spiny neurons (MSN), which constitute the majority of the striatal neurons, express DARPP32 and calretinin and arise from the ventral LGE [15,16].
Cholinergic neurons in the telencephalon (both interneurons and projection neurons) are generated in the MGE. Cholinergic interneurons populate the striatum; cholinergic projection neurons populate the pallidum and the septum and project mainly to neocortex and hippocampus [17].
Cajal-Retzius neurons are a transient population expressing reelin and playing a key role in the formation of the cerebral cortex. They form the cortical layer I and die during the first postnatal week [18].
The mechanisms that orchestrate the generation of these diverse telencephalic neuronal populations have started to emerge during the past 10 years from a series of dye-labeling and fate-mapping studies as well as from genetic loss-of-function and gain-of-function experiments.
A fundamental feature of the nervous system development is the precise temporal sequence of cell type generation. The first neural cells, neuroepithelial (NE) cells, arise from the pluripotent stem cells of the early blastocyst that differentiate from the ectoderm toward the neuroectoderm through a process named neural induction [19,20].
Morphologically, NE cells are columnar epithelial cells that form the neural plate and later on the ventricular zone of the neural tube. They are considered to be primordial neural stem cells that give rise to various types of neurons, followed by glial cells [1,2,21]. The elaborate processes by which NE cells progress toward telencephalic neurons can be divided into several discrete stages:
Early anterior/posterior (A/P) patterning. The NE cells in the neural plate acquire an A/P identity; the anterior ones give rise to the telencephalic primordium.
Dorsal/ventral (D/V) patterning. Once the neural tube is formed and the telencephalic primordium is established, it is subdivided into discrete territories where the NE cells proliferate and transform into neural progenitors that reside in the ventricular zone and in the subventricular zone. In the dorsal telencephalon the progenitor cells are radial glia (RG) and basal (or intermediate) progenitors (BPs) [4,22 –24]. Different progenitor domains are formed in the ventral telencephalon: MGE, LGE, and CGE.
Neuronal specification. Each of the progenitor domains produces specific types of neurons that further develop different neurotransmitter identities and connectivity patterns.
The molecular mechanisms controlling the neural cell fate in these stages have been the focus of intense research in recent years [3,5,10,25 –29]. Extrinsic factors named morphogens can induce 2 or more different cell fates in a concentration-dependent manner by modulating the expression and activity of specific transcription factors (TFs). The TFs in turn also modulate the secretion of morphogens. The combinatorial expression of these TFs instructs each unique progenitor population to generate progenies that are committed to specific neural fates.
The morphogens known to play a role in telencephalon development are sonic hedgehog (SHH), fibroblast growth factors (FGFs), bone morphogenetic proteins (BMPs), Wingless/INT proteins (WNTs), and retinoic acid (RA). They are secreted from specific centers during early stages of development named organizers [30]. Genetic evidence based on loss- and gain-of-function studies indicates that the role of these morphogens can be rather complex. Depending on the developmental stage it ranges from establishment of general patterning characteristics within the NE cells to neuronal specification [3,4,6,22 –24,31 –35].
The main TFs involved in the early patterning and specification belong to homeobox domain and basic helix-loop-helix families, but other TFs such as zinc-finger proteins also have essential functions. The main homeobox domain containing TF gene families are paired-box (eg, Pax6), forkhead box (eg, Foxg1, FoxP1, and FoxP2), NK2 homeobox (eg, Nkx2.1), orthodenthicle homeobox (eg, Otx1 and Otx2), sine-oculis homeobox (eg, Six3), GS homeobox (eg, Gsh2), distal-less homeobox (eg, Dlx1, Dlx2, Dlx5, and Dlx6), LIM homeobox (eg, Isl1, Lhx2, Lhx6, and Lhx8), empty-spiracle homeobox (eg, Emx1 and Emx2), T-box (eg, Tbr1 and Tbr2), cut-like homeobox (eg, Cux1 and Cux2), homeobox/POU domain (eg, Brn2), SATB homeobox (eg, Satb2), and TALE homeobox (eg, Meis2). Other TFs, such as Mash1, Ngn1, Ngn2, and Olig2, belong to basic helix-loop-helix class. Gli family (Gli1, Gli2, and Gli3), Ctip2, and Fezf2 are zinc-finger encoding genes.
In the next sections we review the recently acquired knowledge about the role of these morphogens and TFs in mouse embryonic telencephalon development, followed by development-related approaches for directed differentiation of embryonic stem (ES) cells. The molecular mechanisms related to neurotrophic factors, Notch, extracellular matrix proteins, and cell-cycle signaling, are not reviewed here.
Based on a comparison with in vivo telencephalic neurogenesis, we propose that the expression of specific combinations of TFs can serve as milestones for the differentiation of ES cells toward specific types of telencephalic neurons.
In Vivo Mouse Telencephalic Neurogenesis: Stage-Related Morphogens and TFs
Early A/P patterning
A/P patterning starts to emerge in parallel with neural induction, before and during gastrulation. At E8.5, in regions of the embryo protected from the influence of caudalizing factors, such as WNTs, BMPs, and RA, or where their antagonists are secreted, the NE cells develop an anterior character and form the prospective forebrain (future telencephalon and diencephalon) [6,30,36 –38]. FGFs (eg, FGF8, FGF3) are expressed early on at the anterior tip of the neural plate and then maintained in the anterior limit of the neural tube named anterior neural ridge [35]. Although not a primary inducer of the telencephalic fate, FGF signaling influences the telencephalic gene expression [35,39]. The telencephalic neuroepithelium is first characterized by the expression of the TFs FoxG1 (also named BF1) [40], Pax6 [41,42], and Gli3 [43]. The anterior phenotype also expresses TF genes like Six3, Otx1, and Otx2. Six3 is expressed in the telencephalic and eye field regions of the prospective forebrain [44]. Otx2 is required for the induction of brain structures anterior to the hindbrain, whereas Otx1 is expressed in the midbrain and telencephalon [45,46].
D/V patterning
With regard to the location and timing of telencephalic progenitor generation, different morphogens and TFs are involved in their patterning. Pax6, WNTs, and BMPs pattern the telencephalic progenitors dorsally [5,32 –34,47], whereas SHH patterns them ventrally [31,48 –50]. FoxG1 and FGFs are involved in both ventral and dorsal patterning [3,33,35,51 –55]. Gli3 is likely to be the convergence point for WNT and SHH signaling in regulating the dorsal versus ventral fate of the telencephalic progenitors, and recently has been designated as the main regulator of the telencephalic D/V patterning [3,43,56,57]. Gli3 functions primarily as a transcriptional repressor, whereas Gli1 and Gli2 function primarily as transcriptional activators. Only Gli3 repressor is required for the suppression of ventral telencephalic markers in the dorsal telencephalon, whereas the 3 Gli TFs in their activator forms are involved in the specification of distinct progenitors [57]. The domains of the main TFs and morphogens and the key genetic pathways that interact to pattern the embryonic telencephalon are presented in Fig. 2.
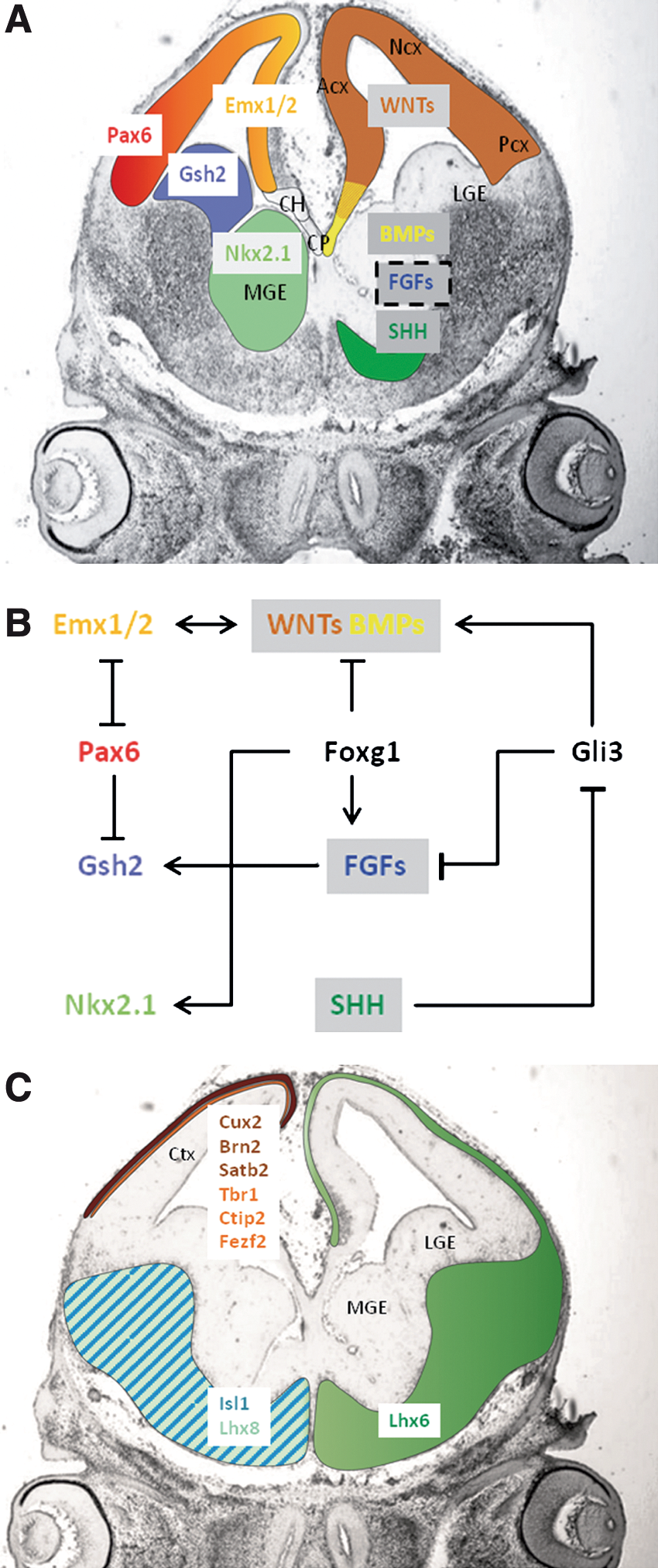
Key transcription factors and morphogens related to the development of mouse embryonic telencephalon.
Dorsal patterning involves at first Pax6, which is essential for setting up the sharp border between ventral and dorsal telencephalon. At present, the signals that regulate the expression of Pax6 remain unknown. At the neural tube stage, its expression is downregulated in ventral region. Pax6 interacts with Gli3 to promote dorsal fate [58]. Gli3 is partially required to maintain Pax6 expression [43]. Pax6 is further expressed in the lateral and ventral parts of the dorsal telencephalon, and downregulated in its medial part [37] (Fig. 2). At the progenitor cell type level, Pax6 is known to be expressed in RG. The other dorsal progenitors—BPs—lack Pax6 and express Tbr2 and Cux1/2 [59,60].
Initially, Gli3 is present in almost the entire telencephalic neuroepithelium. Further on, its expression is markedly reduced in the MGE. Some of the cooperative dorsalizing actions of Pax6 and Gli3 seem to be mediated through Emx1 and Emx2, which seem to act genetically downstream of Gli3. Pax6 cooperates with Emx genes to specify telencephalic precursor cells [61,62]. Emx1 and Emx2 are expressed in the medial part of the dorsal telencephalon (Fig. 2).
Gli3 determines the dorsal identity of the precursors by inducing WNTs and BMPs [3,29,57]. WNTs are expressed in the dorsal telencephalon with the exception of the cortical field [33] (Fig. 2). BMPs are expressed dorsomedially, especially in the cortical hem and cortical field [34,63,64] (Fig. 2A). Specific WNTs are involved in the expansion of defined progenitor cell populations. Canonical WNT signaling promotes cortical progenitor proliferation. Emx2 might promote WNT gene expression [3,57,65,66]. BMPs and WNTs regulate Emx1/2 expression and promote the development of the hippocampus [67].
FoxG1 collaborates with FGFs in partitioning the dorsal telencephalon into the anterior–lateral neocortex and the posterior–medial territories [54,55]. In addition, FGFs and FoxgG1 cooperate with Lhx2, which acts as a selector gene that specifies cortical fates, both hippocampal and neocortical. Lhx2 also represses expansion of the cortical hem and antihem [68,69].
SHH is centrally involved in the ventral patterning of the nervous system [31,48,70]. Expression of SHH is first observed at embryonic day (E) 8.5 in structures adjacent to the ventral telencephalon, by E9.5 in the MGE [31]. The SHH-mediated induction of Nkx2.1 expression is the hallmark of the initiation of MGE development. At E9.5, Nkx2.1 appears within the ventral telencephalic domain, defines the MGE at the molecular level, and persists in this region throughout development (Fig. 2). Around E10.0 the expression of Gsh2 accompanies the emergence of the more dorsally positioned lateral domain that later forms the LGE (Fig. 2). The mutual antagonism between Pax6 and Nkx2.1 and then Gsh2 is required for the correct positioning of the D/V boundary [71 –73].
After E10.0, ventral patterning is already established and SHH activity is predominantly required for the proliferative control of progenitors. At later stages, LGE/CGE regions will depend on SHH signaling [31].
Circumstances that perturb SHH or FGF signaling result in a disruption of the ventral telencephalic patterning [52]. SHH signaling is required to maintain the expression of FGFs. SHH promotes FGFs expression indirectly, by inhibiting Gli3 repressor function [3,52,56] (Fig. 2). In addition, FoxG1 can induce the expression of the fgf genes and it is absolutely required for the establishment of ventral identity in the telencephalon [3,53,55]. Differential activation of FGF signaling in the MGE versus the LGE/CGE may be functionally responsible for the differential fates of progenitors.
RA is known for its caudalizing effect but it was shown to contribute to the patterning of the lateral telencephalon and probably participates in setting-up the D/V boundary by acting on Gsh2, Meis2, and BMPs [5,74 –76]. It could be also implicated in the differentiation of the RG into BPs [61,77].
Neuronal specification
The mechanisms of neuronal specification in the dorsal telencephalon have been extensively studied in the context of cerebral cortex development. The dorsal progenitors produce neurons, in a tightly controlled temporal order from E10.5 to E17.5. Pax6, Ngn1, and Ngn2 instruct glutamatergic identity and inhibit astroglial differentiation [78 –81]. Pax6 is a direct activator of Ngn2 in early cortical progenitors [82], although Pax6 can promote neurogenesis in an Ngn-independent pathway as well [47]. The induction of differentiation by Ngns involves the sequential activation of the expression of other TFs such as NeuroD, Tbr1, and Tbr2 [78]. NeuroD has been implicated in the terminal differentiation of the hippocampus [83]. WNTs promote neuronal differentiation in different progenitor cell populations [84]. In late cortical progenitors, Ngn1/2 is induced by WNTs [81,83]. BMPs inhibit neurogenesis but could participate in late neuronal specification and maturation of different subpopulations [85].
The differentiation of specific populations of projection neurons is controlled by neuronal subtype-specific genes, which have only begun to be identified. The timing of cortical neurogenesis is encoded within lineages of individual progenitor cells, with different locations [86].
The earliest born neurons form a layered structure termed the preplate, which is later split into the superficial marginal zone and the deeply located subplate. The cortical plate, which will give rise to 6-layered neocortex, begins to develop between these 2 layers. The later born neurons arriving at the cortical plate migrate past earlier born neurons [4,86].
Projection neurons in the mammalian cerebral cortex comprise 2 broad classes: those that extend axons within the cortex, including those that cross the corpus callosum to the contralateral hemisphere, and those that form connections with subcortical targets. The subcortical projection neurons are located primarily in cortical layers V and VI, and the neurons with cortical and callosal projections are found in all layers but are particularly abundant in layers II through IV. During development, neurons in different layers are generated in an inside-first, outside-last order, and newly postmitotic neurons are specified to adopt the laminar positions characteristic of their birthdays [4,25,87].
Neurons that populate distinct cortical layers express specific combinations of markers:
Tbr1, Fezf2, Ctip2, and Otx1 are related to differentiation of certain glutamatergic neurons, especially in deep cortical layers (V-VI) and subplate neurons. The most studied are the subcerebral projection neurons of layer V, which express at a high-level Ctip2, Fezf2, and Otx1 [10,46,59,86,88 –90].
Cux2 and Lhx2 are expressed in the BPs and further in upper-layer neurons (II-IV). Brn2 and Satb2 are related to the development of late cortical plate and callosal neurons [60,88,91]. Tbr2 is expressed in BPs, supporting the hypothesis that upper-layer neurons are generated in a late time point via the BPs [24,59,88,89].
Tbr1 is also expressed in the Cajal-Retzius cells of layer I during development. FoxG1 and midline signaling factors are additionally implied in the fate of these cells [92,93].
Regarding ventral neuronal specification, spatial biases have been found in the generation of individual neuronal populations in the MGE and LGE/CGE. However, a causal link between the function of these spatially restricted TFs and individual neuronal types has to be more investigated.
Mash1 is the main neurogenic TFs in the ventral telencephalon and is involved also in the neurotransmitter identity specification, being a selective instructor of GABA-ergic identity [79,92 –98]. Olig1/2 can promote both neuronal and oligodendroglial fates while inhibiting astrogliogeneis [99].
Nkx2.1 and Gsh2 progenitor domains do not give rise to homogeneous populations of neurons, but it is a temporal specification of cortical interneuron subtypes [25,98,100 –102]. The further subdivision of these broad domains occurs by the spatially restricted expression of additional TFs [12,103,104]. Dlx genes (Dlx1, 2, 5, and 6) are expressed in ventral progenitors and neurons in MGE, LGE, and CGE, and are likely to play a role in neural specification [95,105,106].
Notably, it has been shown that the expression of SHH is required during distinct developmental windows for the specification of neuronal identity, and it is mediated mainly via the activator forms of Gli1, Gli2, and Gli3 [31,57]. FGF signaling may ultimately influence the generation of cell diversity within the ventral telencephalon [33,52].
As a general pattern, the earlier-born ventral cells give rise to projection neurons, whereas the more dorsally positioned later-born cells generate interneurons. MGE is characterized by the early production of cholinergic projection neurons from its ventral part, followed by the late production of GABA-ergic and cholinergic interneurons from the dorsal domains, at later time points.
Two TFs are detected at E12.5 in the MGE only, Lhx6 and Lhx8 (or Lhx7) [103]. Lhx6-expressing neurons have characteristics of proto-GABA-ergic neurons, whereas Lhx8 is involved in the differentiation of specific cholinergic neurons. The cortical interneurons derived from MGE express Lhx6 [107 –109] (Fig. 2). Later, the cholinergic interneurons in the striatum express Lhx8. The cholinergic projection neurons express Lhx8 or Isl1 [17,108]. The differentiation of a common precursor into mature subtypes is regulated by the combinatorial activity of the Lhx6, Lhx8, and Isl1 (Fig. 2). Thus, it appears that a LIM homeodomain transcriptional code might determine cell-fate specification and neurotransmitter identity in neuronal subpopulations of the ventral telencephalon.
Ventral LGE generates GABA-ergic projection neurons that also express Isl1 during early specification, followed by the expression of other striatal specific TFs such as FoxP1, FoxP2, Ctip2, and Meis2 [110 –112]. Later, dorsal LGE generates interneuron that migrate to the olfactory bulbs [15,102].
Telencephalic Differentiation of Mouse ES Cells
Mouse ES (mES) cells have emerged as a powerful tool for developmental biology. Several reports described different methods for the in vitro generation of neural cells from mES cells. The use of developmental morphogenetic cues has allowed the targeted differentiation of cell types with positional identity, for example, the directed differentiation of specific neuronal populations, such as spinal motor neurons [113] or midbrain dopaminergic neurons [114]. The ability of mES cells to generate forebrain-like progenitors has been reported, and also their capacity to differentiate into GABA-ergic and glutamatergic neurons [115 –119]. Many of the recent ES-derived neural differentiation methods yield a high degree of neural induction, both in suspension and in adherent culture, generating higher percentages of NE cells, which express early neural markers such as Sox1 and nestin, and lack pluripotent or endodermal/mesodermal markers (eg, Nanog, Oct4, brachiury, and α-fetoprotein).
A challenging problem remains the homogeneous generation of specific subtypes of telencephalic neurons. Lately, several studies have focused on mES-derived telencephalic progenitors and the specific neuronal populations they generate in vitro and in vivo [117,118,120 –122]. In the following section, we focus exclusively on a selection of recently published protocols that exemplify the developmental approach to generate specific telencephalic populations, pointing on the relevant TFs and morphogens in each stage.
Early A/P patterning
The generation of defined telencephalic precursors from mES cells based on studies of forebrain development was first documented by the Sasai group in Japan [118]. Significant improvements of the protocol (regarding neural induction success and telencephalic patterning) were published by the same group [120], or others [122], during the following years.
Blocking the WNT and BMP pathways by applying the antagonists DKK1 and BMPR1A-FC, respectively, during the first 5 days of suspension culture using serum replacer (SFEB culture) caused neural induction in >90% of mES cells and maintained around 35% of the progenitors as telencephalic, expressing FoxG1 [118]. Using the same culture system and adding a size exclusion criterion for the cellular aggregates, Eiraku et al. reported a greater proportion (up to 70%) of FoxG1-positive telencephalic progenitors [120]. Modifying this suspension culture protocol by using noggin and a mixture of serum replacer and N2 supplement during the first 5 days, Maroof et al. reported also a massive FoxG1-expressing telencephalic progenitor generation [122].
Other protocols using serum-free conditions in adherent culture systems, derived from the method first described by Ying et al., showed improved neural induction and also a primitive forebrain identity [119]. Using the same approach, Gaspard et al. showed that the majority of mES cells in adherent cultures, at low density, in a chemically defined default medium and devoid of serum or any morphogen, but in the presence of insulin, converted to neural progenitors [123]. The telencephalic identity of these progenitors was determined after concomitant dorsal patterning (see below).
In another approach using for neural induction a coculture with stromal cells, the majority of the cells (80%) expressed nestin and FoxG1, in comparison with the RA treatment, where FoxG1 expression was not detected [117].
D/V patterning
In all the studies presented, ES-derived neural progenitors expressed markers of the dorsal and ventral telencephalon in different proportions, at different time points.
Watanabe's protocol generated both dorsal and ventral progenitors, but the SHH treatment on SFEB-induced telencephalic progenitors suppressed Pax6 and induced the ventral marker Nkx2.1. SHH did not cause substantial difference in Gsh2 expression [118]. In the modified SFEB culture the majority of FoxG1-positive cells at day 10 coexpressed the dorsal marker Emx1 [120].
In Maroof's protocol, the cellular aggregates were dissociated at day 5, following by the treatment with SHH and FGF2 in high-density adherent culture. At day 8 the majority of FoxG1 progenitors in the rosette-like clusters expressed Nkx2.1 [122].
At the telencephalic progenitor early stage, in monolayer culture, Gaspard et al. have shown that, the majority of progenitors exhibited a ventral phenotype with a high expression of Nkx2.1, which was attributed to the endogenous SHH signaling. Blocking SHH signaling by cyclopamine converted most of the ventral telencephalic precursors to dorsal progenitors, the majority expressing typical markers of the dorsal telencephalon: Pax6 and Emx1/2 [123].
Neuronal specification
The same studies demonstrated that mES cell-derived progenitors can be directed to neurons that express the excitatory neurotransmitter glutamate and the inhibitory neurotransmitter GABA [118,120,122,123].
The dorsal progenitors generated with both Gaspard's and Eiraku's protocols produced mature neurons with many features of cortical pyramidal neurons, able to form glutamatergic synapses. Both groups showed that the neuronal populations were generated in a temporal manner similar with in vivo corticogenesis. The first neurons generated in their cultures were reelin-positive Cajal-Retzius-like and subplate-like neurons expressing Tbr1, followed by the glutamatergic neurons generated in an inside-first, outside-last manner. The majority of the cells generated by Gaspard expressed markers of deep cortical layer V and VI neurons, like Tbr1, Otx1, Ctip2, and FoxP2. Later a small fraction expressed upper layer markers as Cux1 and Satb2. Eiraku et al. generated also deep layer neurons, positive for Ctip2 and Emx1 in the first 9 days of neuronal differentiation of dorsalized progenitor, and only a small proportion of Brn2, Cux1, and Satb2-expressing neurons in later time points (days 12–19).
Additionally, in Eiraku's suspension culture system, a striking polarized cellular organization was observed, neural progenitors (Emx1, Pax6, and Tbr2 positive) occupy deeper layers of the cellular aggregates, and neurons accumulated at their periphery, following an arrangement highly reminiscent of a nascent cortical primordium [120]. Altogether, these data constitute the first proof that a structure with morphological features of an early CNS can emerge as a self-organizing cytoarchitecture in vitro.
By transplantation of dorsally patterned progenitors into postnatal murine cerebral cortex, the production of cortical projection neurons with the correct morphology and axonal connectivity has been demonstrated [117]. The pyramidal neurons expressed Otx1, Emx1, Fezf2, and Ctip2, corresponding to deep layers neurons. After dissociation and transplantation into the murine cerebral cortex, they integrated and appropriately projected long-distance axons to subcortical targets, without forming tumors [75,117].
Regarding the protocols for ventral neuronal specification, Nkx2.1 and Gsh2-expressing progenitors gave rise to both GABA-ergic projection neurons and interneurons [118,122].
Watanabe showed that SHH treatment did not cause a substantial difference in Mash2 expression. Isl1 was found to be expressed in 25%–30% of the FoxG1-positive telencephalic cells but their subtype identity as GABA-ergic projection striatal neurons or cholinergic projection neurons was not further investigated [118].
Maroof's protocol was shown to give rise predominantly to cortical and striatal GABA-ergic interneurons. By differentiation, on day 12, streams of cells appeared to radiate from the Nkx2.1 clusters, and they expressed Lhx6. Further use of Lhx6::GFP ES cells allowed the isolation of cells with potential for developing into ventral telencephalic subpopulations and their follow-up during transplantation into the postnatal brain. Transplanted Lhx6::GFP cells demonstrated the ability to retain migratory capacity and neuronal commitment without forming tumors and exhibited cortical interneuron characteristics [122].
Telencephalic Differentiation of Human ES Cells
Regarding the differentiation of the human ES (hES) cells toward telencephalic neurons, we also review the recent published protocols that exemplify the developmental approach to the generation of specific telencephalic populations, focusing on the relevant TFs and morphogens in each stage.
Neural induction took place in hES cells similar to mES cells, but with a different timing. Under serum-free conditions, without known morphogens [118,118,124 –127], by coculture with stromal cells [128,129] or using a recently defined protocol with dual-SMAD-inhibition [130], hES cells differentiated into a synchronized population of NE cells organized into neural-tube-like rosettes within 2 weeks, a time corresponding to the development of the neural plate/tube in a human embryo. Around days 8–10, the primitive NE cells expressed PAX6, and the neural fate was consolidated by SOX1 expression around day 14 [122,124 –127].
Early A/P patterning
In the absence of exogenous morphogens, hES cells differentiated into progenitors that uniformly expressed anterior TFs, including FOXG1, OTX2, SIX3, and LHX2 but not posterior TFs [124,126]. Even when the neural induction was performed by coculturing with stromal cells, a large population of neural progenitors exhibited anterior characteristics [129]. FOXG1 appeared at approximately day 10 and it was still expressed at day 24 and even in postmitotic neurons, 1 month after differentiation. PAX6 was coexpressed in nearly all of the FOXG1 telencephalic progenitors (95% of all cells) 1 month after differentiation, in the absence of growth factors or morphogens [131].
D/V patterning
WNTs and their downstream molecules were highly expressed right after the generation of NE cells. In minimal medium and in the absence of known morphogens, hES cell-derived telencephalic progenitors exhibited a dorsal telencephalic trait, which was attributed to endogenous WNT signaling [132]. The activation of SHH and/or inhibition of WNT permitted the specification of ventral telencephalic progenitors.
At the lower dosage, SHH reduced the PAX6-expressing cell population to 61% and increased the NKX2.1 cell population to 30%. At the higher dosage, SHH almost completely eliminated the PAX6-expressing cells, while increasing the NKX2.1 ventral progenitors to 84%. The combination of DKK1 and SHH at the lower dosage significantly increased NKX2.1 and decreased PAX6 and GLI3 expression. High concentrations of SHH significantly inhibited GLI3 expression compared with low concentrations of SHH. The patterning of human neural progenitors by WNT and/or SHH was partially achieved through differentially regulating the expression of active and repressive forms of GLI3. Graded ventralization could be achieved by regulating these 2 signaling pathways. Dosing of a low concentration of SHH alone resulted in the differentiation of both LGE and MGE progenitors, whereas additional WNT inhibition (by DKK1) further ventralized the human neural progenitors, resulting in a predominant population of NKX2.1-expressing MGE progenitors [132]. Aubry et al. patterned the telencephalic progenitors by SHH and DKK1 treatment for 12 days in adherent culture and found a significant upregulation of the LGE markers GSH2 and DLX2 [128].
Neuronal specification
The regionalized dorsal and ventral human telencephalic progenitors further differentiated into functional cortical glutamatergic neurons and telencephalic GABA-ergic neurons, respectively.
Cortical glutamatergic neurons were efficiently generated in the absence of morphogens. This indicates the intrinsic tendency of hES cells to generate cortical neural cells [126]. Neurons differentiated from dorsal progenitors in the absence of exogenous morphogens for 6 weeks exhibited a pyramidal morphology, with extensive neurite outgrowth, and expressed TBR1 and CTIP2 [132].
Eiraku et al. applied the SFEB protocol to the hES cells, showing the formation of polarized cortical tissue. On day 46 the postmitotic neurons expressed TBR1 and CTIP2 [120].
Ventral telencephalic GABA-ergic neurons, including projection striatal GABA-ergic neurons, were induced with a high concentration of SHH or a low dose of SHH together with WNT inhibitors. Culturing ventral telencephalic progenitors for 6 weeks resulted in ∼40% ISL1-positive cells. These results indicate that ventral neural progenitors generated GABA-ergic neurons, including the projection striatal GABA-ergic neurons [132].
Additionally, Aubry et al. focused to the characterization of the striatal-like progenitors, showing that more than a half of the specified neurons were DARPP32 and calretinin positive, exhibiting phenotypic features of MSN. Transplantation of the LGE-patterned progenitors in quinolinic acid–lesioned rats (a model for Huntington disease) confirmed the in vivo specification toward striatal MSN, but also graft overgrowth was observed [128].
hES cell-derived telencephalic glutamatergic and GABA-ergic neurons were reported to be electro-physiologically active [128,132,133].
TF-Related Milestones for Directed ES Cell Differentiation
The above-presented recent results suggest that mES cells and hES cells differentiate into region-specific progenitors and corresponding functional neurons, following similar developmental principles determined in vivo.
Using the above protocols, mouse and hES cell-derived neural cells undergo 3 morphologically identifiable stages, and associated TF-related milestones can be tentatively defined (Fig. 3).
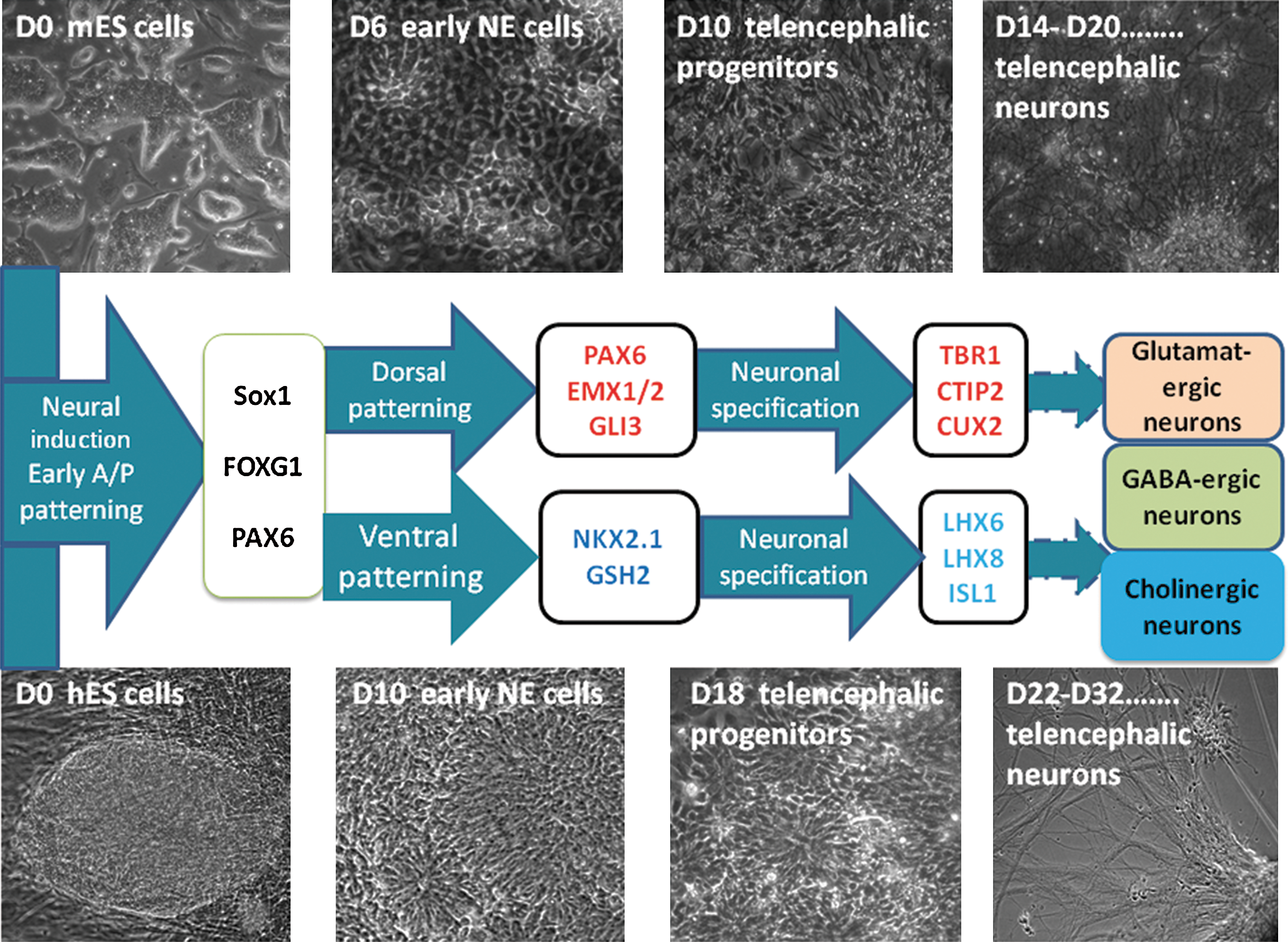
Transcription factor-related milestones achieved during telencephalic-directed differentiation of mouse and human embryonic stem (ES) cells. Three morphologically and transcription factor-related distinct steps of differentiation from mouse and human ES cells: (1) neural induction and early anterior/posterior patterning, (2) dorsal/ventral patterning, and (3) neuronal specification. Phase-contrast images showing ES cell stage (day in vitro D0), neuroepithelial (NE) cell stage (D6 in mouse cells and D10 in human cells), neural progenitor cell stage (D10 in mouse cells and D18 in human cells), and postmitotic neuronal cell stage (up to D14 in mouse cells and D22 in human cells). Transcription factor-related milestones can be defined at these time points, as shown in the middle diagram (see the text for references). Color images available online at
In the first stage, NE cells are characterized by columnar epithelial morphology and correspond to early A/P patterned NE cells expressing Sox1 and FoxG1 for mouse cells and PAX6 and FOXG1 for human cells. PAX6 was recently shown to be the main fate determinant for human NE cells [134].
In the second stage, the D/V-patterned progenitors organized as neural-tube-like rosettes have different dorsal and ventral TF-related milestones: PAX6, EMX1/2, GLI3, NKX2.1, and GSH2, respectively.
During the third stage, neuronal specification, glutamatergic and GABA-ergic neurons generated from dorsal and ventral progenitors, respectively, maintained FOXG1 expression in long-term culture.
The subclasses of glutamatergic neurons generated from both mouse and hES cells were first related to deep layers, expressing TBR1, CTIP2, and FEZF2, but other subpopulations could be present later during differentiation (as upper layer TFs CUX2, BRN2, and SATB2).
The subclasses of GABA-ergic neurons were mainly interneurons positive for LHX6, but also projection neurons, positive for ISL1. LHX8 could be included as a milestone for the telencephalic cholinergic neurons generated via ES cell-directed differentiation. Additionally, other GABA-ergic projection neuron-related TFs could be considered (FOXP1/2 and MEIS2).
Telencephalic Neurons from Pluripotent Stem Cells: Perspectives for Disease Modeling and Brain Repair
Beyond its implications for our understanding of normal telencephalic development, in vitro telencephalic differentiation opens new venues to apply ES cell-based technology to telencephalic pathology.
Purification of certain neuronal subtypes derived from telencephalon represents an important goal. The prospective isolation of ES cell-derived GABA-ergic interneurons at early stages of fate commitment [122] represents a powerful paradigm that should be applicable for generating other types of telencephalic neurons.
Advances in this research field could develop the modeling of telencephalic diseases such as specific forms of mental retardation, epilepsy, Huntington disease, Alzheimer disease, and amyotrophic lateral sclerosis and the rational design of telencephalic neuron replacement therapies. Although using such a system for brain repair seems like a relatively distant perspective, it was recently shown that cortical progenitors have the remarkable ability to engraft and connect in the adult brain [117].
Another, more immediate possibility will be to use the directed telencephalic differentiation systems to induced pluripotent stem cells derived from patients with telencephalic diseases [135 –138]. This approach would provide the opportunity to model diseases of the telencephalon, and to complement the existing animal models that could be partially inadequate regarding some specific features of the normal and pathological human brain.
Footnotes
Acknowledgments
This work was supported by CNCSIS Romania (Project IDEI 626/2007). The authors acknowledge Dr. Galina Apostolova, Dr. Laura Suciu, and Dr. Alina Codita for the critical reading of the article.
Author Disclosure Statement
No competing financial interests exist.