
Abstract
In recent years, various types of stem cells have been characterized and their potential for cardiac regeneration has been investigated. We have previously described the isolation of broadly multipotent cells from amniotic fluid, defined as amniotic fluid stem (AFS) cells. The aim of this study was to investigate the therapeutic potential of human AFS cells (hAFS) in a model of acute myocardial infarction. Wistar rats underwent 30 min of ischemia by ligation of the left anterior descending coronary artery, followed by administration of hAFS cells and 2 h of reperfusion. Infarct size was assessed by 2,3,5-triphenyltetrazolium chloride staining and planimetry. hAFS cells were also analyzed by enzyme-linked immunosorbent assay to detect secretion of putative paracrine factors, such as the actin monomer-binding protein thymosin β4 (Tβ4). The systemic injection of hAFS cells and their conditioned medium (hAFS-CM) was cardioprotective, improving myocardial cell survival and decreasing the infarct size from 53.9%±2.3% (control animals receiving phosphate-buffered saline injection) to 40.0%±3.0% (hAFS cells) and 39.7%±2.5% (hAFS-CM, P<0.01). In addition, hAFS cells were demonstrated to secrete Tβ4, previously shown to be both cardioprotective and proangiogenic. Our results suggest that AFS cells have therapeutic potential in the setting of acute myocardial infarction, which may be mediated through paracrine effectors such as Tβ4. Therefore, AFS cells might represent a novel source for cell therapy and cell transplantation strategies in repair following ischemic heart disease, with a possible paracrine mechanism of action and a potential molecular candidate for acute cardioprotection.
Introduction
V
Many different stem cell populations have been tested in acute myocardial infarction (AMI) studies, namely unfractionated bone marrow-derived cells (BMCs), hematopoietic stem cells (HSCs), endothelial progenitor cells (EPCs), mesenchymal stem cells (MSCs), and endogenous cardiac stem cells [5]. Although their mechanism of action has been only partially elucidated, and it is likely to involve multiple pathways, these cells have been able to commonly act in an allogenic setting [6]. This is particularly important for clinical translation where, for the treatment of AMI, transplantable cells should ideally be available soon after admitting a patient to hospital. To facilitate this, stem cells should be derived from a low immunogenic source, be readily expanded in vitro, and survive cryopreservation and freeze–thaw cycles. Bone marrow-derived MSCs (BM-MSCs) have been limited by the need for bone marrow aspiration in the ischemic patient and the relative low cell numbers obtainable without expansion [7]. A possible alternative is the use of amniotic fluid stem (AFS) cells, as they retain a more immature potential than adult stem cells and seem to have wider proliferative and differentiation abilities [8]. Thus, AFS cells may provide a unique opportunity to be banked and used in regenerative cell therapy [8]. They possess unique properties, such as survival at lower oxygen tension and the ability to withstand protracted cryopreservation while maintaining their phenotype and their self-renewal potential [9].
The aim of this work was to analyze the cardioprotective effect of AFS cells in a rat model of AMI and to define whether their action could be mediated by secretion of critical paracrine factors, such as thymosin beta 4 (Tβ4), recently identified as an important cardioprotective and proangiogenic agent [10 –14].
Materials and Methods
Rat model of acute myocardial infarct
Adult male Wistar rats (Charles River, United Kingdom) weighing between 200 and 300 g were housed and maintained in a controlled environment. All surgical and pharmacological procedures were performed in accordance with the Animals (Scientific Procedures) Act 1986 (Home Office, United Kingdom). Rats were anesthetized by intraperitoneal injection of sodium thiopentone (120 mg/kg body weight; Intraval, Merial) and analgesia was provided by subcutaneous injection of buprenorphine (0.2 mg/kg body weight; Vetergesic Alstoe Animal Health). Body temperature was continuously monitored using a rectal probe and maintained at 37°C with a heating pad. Electrocardiography was acquired via subcutaneous electrodes, and the right common carotid artery was cannulated and connected to a transducer for blood pressure monitoring (PowerLab with Chart5 software; ADInstruments). Animals underwent tracheotomy and were ventilated via an endotracheal tube with oxygen (Harvard Apparatus). The right external jugular vein was cannulated to provide intravenous (i.v.) access for the treatment injection [either phosphate-buffered saline (PBS) or human AFS (hAFS) cells or hAFS cell-conditioned medium (hAFS-CM)]. A thoracotomy was performed to expose the heart and myocardial ischemia was achieved by ligation of the left anterior descending coronary artery (LAD) using a 6/0 silk suture connected to a snare. After 30 min of ligation, the snare was released, injection was performed via the right external jugular vein, and the heart was reperfused for 2 h.
hAFS cells expansion and transplantation
hAFS cells were prepared according to methods previously described [8]. Samples of amniotic fluid were collected by amniocentesis from women (mean gestational age: 14 weeks) during routine prenatal screening, after obtaining a written consent. Samples were spun at 1,200 rpm and pellets resuspended and seeded in Chang Medium [α-MEM medium (Invitrogen) containing 15% fetal bovine serum, 1% glutamine, and 1% penicillin/streptomycin (Gibco), supplemented with 18% Chang B and 2% Chang C (Irvine Scientific)] at 37°C with 5% CO2 atmosphere. After 3 days, nonadherent cells and debris were discarded and the adherent cells were cultivated to preconfluence. Adherent cells were then immunomagnetically sorted for the expression of the stem marker c-kit using a mouse monoclonal anti-c-kit (CD117) antibody (Santa Cruz) and an anti-mouse IgG CELLection Dynabeads M-450 antibody (Miltenyi Biotech). c-kit+ hAFS cells were replated at a density of 2×103 cells/cm2, cultured in Chang medium in 5% CO2 at 37°C, expanded and subsequently cloned by limiting dilution, and maintained at subconfluence.
Cryopreserved hAFS cells were thawed and cultured to subconfluence and then detached from culture plastic dishes with trypsin 0.05–EDTA 0.02 (w/v) sterile solution (Biochrom AG) and centrifuged for 5 min at 1,200 rpm. The cells were then resuspended in 500 μL of PBS 1X (Gibco) sterile solution for injections.
To investigate any in vivo paracrine or cytokine-related effects mediated by the injected stem cells, hAFS cells-conditioned medium (hAFS-CM) was also used to treat the animals. Medium was obtained from hAFS cells cultured in vitro to subconfluence. hAFS cells-CM was collected after 72 h, centrifuged to exclude any debris, and then stored at −80°C prior to the in vivo administration.
According to the treatment, PBS vehicle, hAFS cells, or hAFS-CM solution–injected animals were randomly divided into 3 groups:
(a) Injection of 500 μL PBS solution as control (n=8)
(b) Injection of 5×106 hAFS cells in 500 μL of PBS (n=8)
(c) Injection of 500 μL hAFS cells-CM (n=6)
To avoid formation of any emboli, hAFS cells were delivered slowly over approximately 1 min.
Assessment of area at risk and infarct size by planimetry
To assess the infarct area, hearts were processed and 2,3,5-triphenyltetrazolium chloride (TTC) staining was performed as previously described [15]. As infarct size (IS) is a determinant of cardiac remodeling and dysfunction [16,17], assessing the proper quantification of myocardial infarction is of critical importance. Histopathology is not sensitive enough for properly evaluating the necrosis occurring during the very initial hours after coronary occlusion; thus, we have opted for the TTC staining method that has been well validated in literature [18].
After 2 h of reperfusion, the LAD was reoccluded and 2 mL of 2% Evans Blue solution (Sigma) was injected into the heart via the external jugular vein to demarcate the area at risk (AAR) in the heart, which is the region of perfusion deficit. The heart was then extracted, sliced from and perpendicular to the apex into 2-mm-thick sections, and incubated in 1% TTC solution (Sigma-Aldrich) at 37°C for 15 min to identify the area of infarct. Slices were then fixed in 4% paraformaldehyde (Sigma-Aldrich) at 4°C for 12 h and subsequently placed between glass slides and imaged using a flat-bed scanner with a resolution of 600 dpi. Images were anonymized to blind the observer to the treatment groups, and the sizes of the left ventricle (LV), AAR, and IS were measured by planimetry using ImageJ software (NIH).
Immunostaining analysis for distribution of hAFS cells and apoptosis of myocardial cells in vivo
Heart, lungs, liver, and spleen were harvested from animals as reperfusion time ended. Tissues were fixed in 4% paraformaldehyde solution, incubated in 30% sucrose–PBS solution (Sigma), embedded in OCT solution (Sigma), and snap-frozen in 2-methyl butane and liquid nitrogen. Organs were cut into 8-μm-thick frozen sections and processed for immunostaining of cardiovascular markers including α-smooth muscle actin [α-SMA; mouse monoclonal anti-α-SMA IgG-FITC-conjugated antibody (Sigma) and rabbit polyclonal anti-α-SMA IgG antibody (Abcam)] and von Willebrand Factor (vWf; rabbit polyclonal anti-vWf IgG antibody; Chemicon). Human AFS cells were detected using anti-human-specific mitochondria (Hum Mit) antibody (mouse monoclonal IgG; Abcam). Anti-rabbit Alexa Fluor 488-conjugated and anti-mouse Alexa Fluor 594-conjugated secondary antibodies were used (Molecular Probes, Invitrogen). Cell nuclei were stained with 4'-6-diamidino-2-phenylindole solution at room temperature (Vectashield; Vector Laboratories, Inc.).
To evaluate the number of apoptotic cells in the hearts of rats injected with the 3 different treatments, colorimetric terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) analysis and immunostaining for the active form of the caspase-3 protein were performed on cryosections.
TUNEL is a common method for detecting DNA fragmentation that results from apoptotic signaling cascades and it has become one of the main methods for detecting apoptotic programmed cell death. Active caspase-3 is another apoptotic cellular marker as it is a critical executioner in the apoptosis signaling pathway. Analyses were performed using the DeadEnd™ Colorimetric TUNEL System (Promega) following the manufacturer's instructions and using the anti-cleaved caspase-3 (Asp175) antibody (caspase-3, rabbit IgG; Cell Signaling Technology).
A Zeiss Axio Imager Z.1 epifluorescence microscope with AxioCam Mrm digital video camera, a Zeiss Axioplan 2 microscope with AxioCam Hrc digital video camera, and a Leica TCS SP5 confocal microscope were used to analyze cell staining and images were obtained using a Leica DC300F digital video camera.
In vitro characterization of hAFS cells' vascular potential
hAFS cells were characterized to analyze their vascular differentiation potential in control condition in vitro. The cells were cultured to subconfluence on a glass coverslip in Chang medium for 2, 4, 6, and 10 days and then processed for immunostaining for smooth muscle and endothelial markers, including anti-α-SMA [mouse IgG-FITC-conjugated antibody (Sigma) and rabbit IgG antibody (Abcam)] and anti-human vWf (rabbit IgG antibody; Abcam). hAFS cells were also stained using the anti-Hum Mit antibody (mouse monoclonal IgG; Abcam). Anti-rabbit Alexa Fluor 488-conjugated and anti-mouse Alexa Fluor 594-conjugated secondary antibodies were used (Molecular Probes, Invitrogen). Cell nuclei were stained with 4'-6-diamidino-2-phenylindole solution at room temperature (Vectashield; Vector Laboratories, Inc.). Observations were made using a Zeiss Axio Imager Z.1 epifluorescence microscope with AxioCam Mrm digital video camera.
Analysis of secretion of thymosin β4 in the culture medium of hAFS cells
Enzyme-linked immunosorbent assay (ELISA) analysis for thymosin β4 (Tβ4) expression was performed on in vitro hAFS-CM. The analysis was carried out using a Tβ4 ELISA kit (Immundiagnostik; Germany) according to the manufacturer's instructions. Briefly, hAFS cells were cultured in vitro and their conditioned medium was collected after 72 h from seeding, centrifuged to exclude any debris, and then processed for the analysis by ELISA.
Statistical analysis
All values were expressed as mean±standard error. Statistical differences between groups were evaluated by 1-way analysis of variance test followed by Dunnett's post hoc test, t-test, and Mann–Whitney U test (SPSS Statistics 15.0 and GraphPad Prism 5.0). A P value of <0.05 was considered significant.
Results
Assessment of area at risk and infarct size
In Figure 1A–C, representative planimetric images of Evan's blue/TTC heart sections from each group (PBS, hAFS cells, and hAFS-CM treatment) are shown. The region of perfusion deficit or AAR of infarction (expressed as a percentage to the LV, %AAR/LV, in red) was similar across all groups and no significant differences were detected (PBS control: 56.8%±2.1%; 5×106 hAFS cells: 55.7%±2.2%; hAFS-CM: 54.7%±5.4%; P>0.05; Fig. 1D, top). The small variability of %AAR/LV between the groups suggests high reproducibility of LAD occlusion. The IS (expressed as the ratio between the volume of infarct and the AAR, % In/AAR) of cell- and conditioned medium-treated animals was compared with the PBS control group (PBS: 53.9%±2.3%; Fig. 1D, bottom). Animals treated with 5×106 hAFS cells and with their conditioned medium demonstrated a significant reduction in the IS, compared with the untreated animals (5×106 hAFS cells: 40.0%±3.0%, **P<0.01; and hAFS-CM: 39.7±2.5%, *P<0.05; Fig. 1D, bottom). IS between the hAFS and hAFS-CM groups was not statistically different (P>0.05).
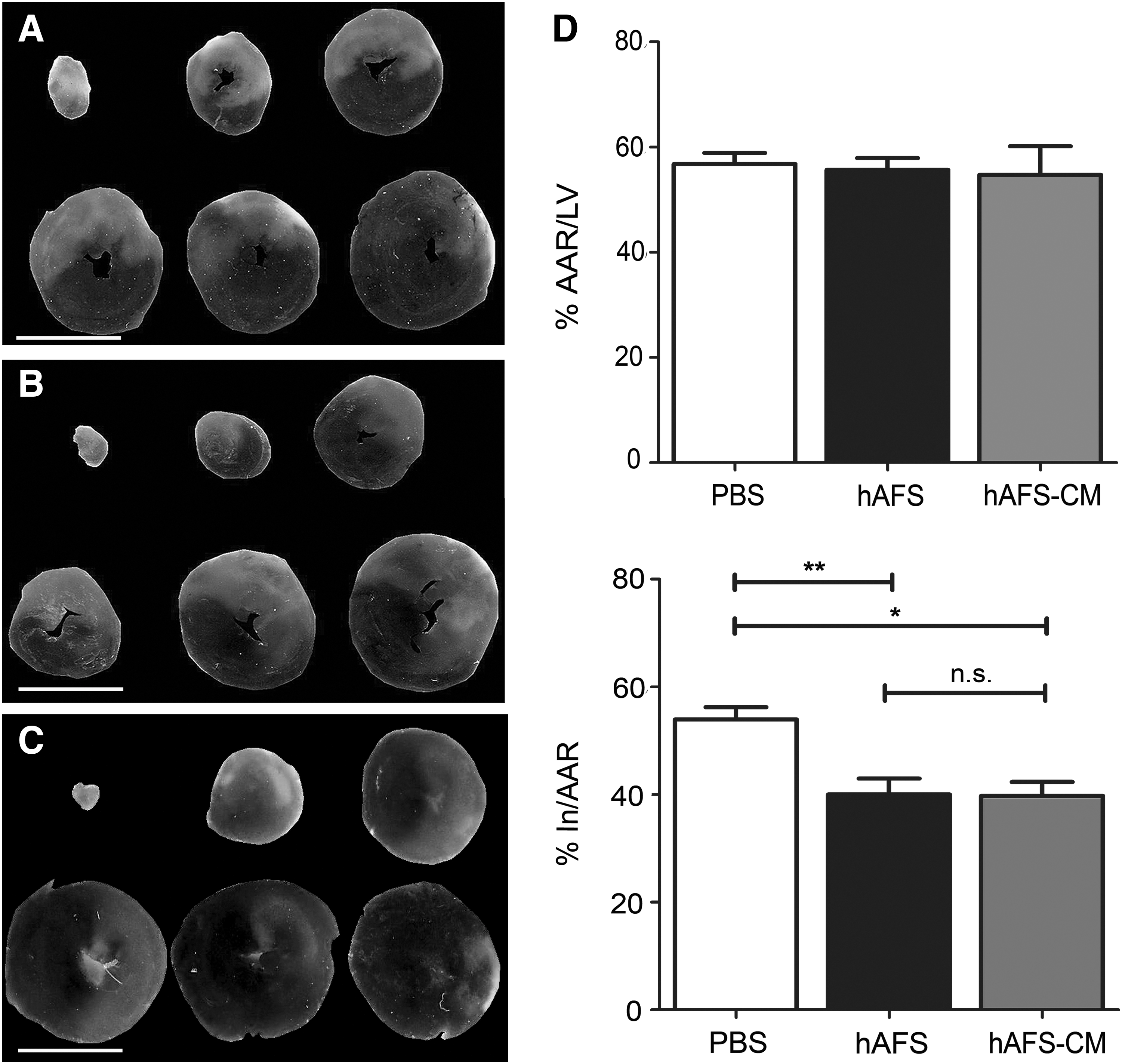
Assessment of AAR of infarction and infarct size. 2,3,5-Triphenyltetrazolium chloride staining on heart of control rat injected with 500 μL PBS
hAFS cell transplantation and distribution in vivo
Injection of hAFS cells and hAFS-CM was not associated with any difference in the rate of arrhythmias. We readily detected ischemia/infarction-specific electrocardiography changes such as ST elevation following LAD ligation, whereas arrhythmias were only occasionally observed and were almost always transient (eg, <1 min), non lethal, and self-limiting. The rate of arrhythmias was similar amongst all groups.
Serial heart, lungs, spleen, and liver sections from animals injected with hAFS cells were stained with the anti-Hum Mit antibody to evaluate hAFS cell distribution and localization (Fig. 2). hAFS cells were found in the host myocardium (Fig. 2A–C); notably, most hAFS cells detected in the cardiac tissue were viable, as fewer than 10% were found to express the apoptotic protein caspase-3 (Fig. 2C). About 32%±9% of the hAFS detected in the host heart costained for the endothelial marker vWf and almost all of them (82%±10%) were positive for the expression of α-SMA (Fig. 2D–I), indicative of potential commitment of the donor hAFS cells to the endothelial and smooth muscle cell (vascular) lineages in vivo.
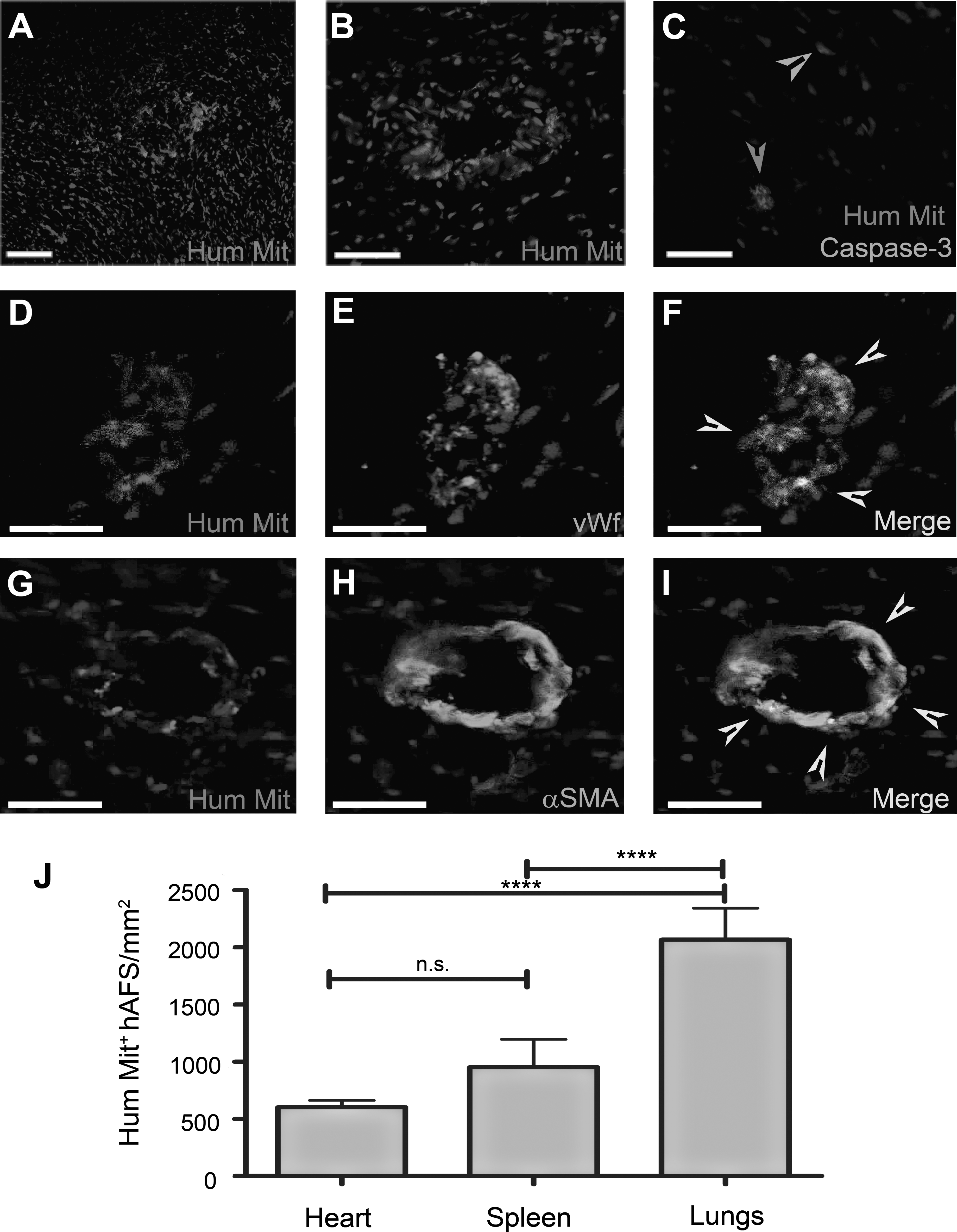
Immunostaining analysis of the heart of animals treated with hAFS cell injection and hAFS cell distribution in vivo. hAFS cells were found in the myocardium at 2 h after LAD ischemia/reperfusion and intravenous injection (as labeled by the Hum Mit antibody).
The hAFS cells found in the host myocardium after 2 h of reperfusion were 601.5±60 cells/mm2, hAFS cells found in the spleen were 951.4±242.6 cells/mm2, and human cells in the rat lungs were 2066±275.4 cells/mm2 (Fig. 2J). No cells were found in the liver.
Assessment of myocardial apoptosis
TUNEL-positive apoptotic cardiac cells in the rats receiving PBS (Fig. 3A), 5×106 hAFS cell injection (Fig. 3B), and hAFS cells-CM (Fig. 3C) were 3840±397.6 cells/mm2 (PBS; Fig. 3G, top), 723.7±223.6 cells/mm2 (hAFS; Fig. 3G, top), and 1500±207.1 cells/mm2 (hAFS-CM; Fig. 3G, top), respectively.
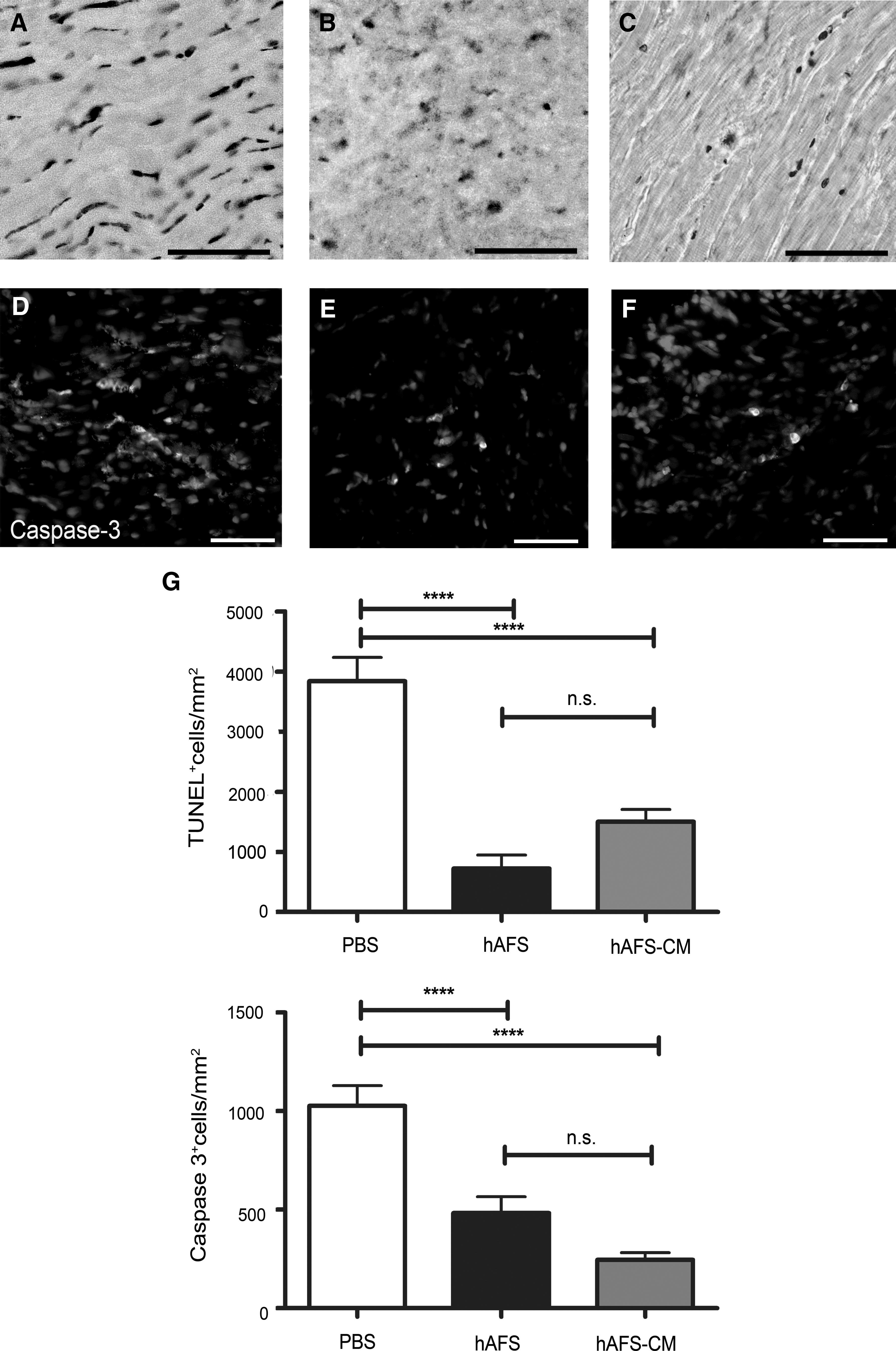
Analysis of myocardial apoptosis after 2 h of reperfusion following LAD occlusion. In the rats injected with PBS, TUNEL-positive apoptotic cardiac cells
In the rats injected with the PBS solution, active caspase-3-positive myocardial cells (Fig. 3D) were 1027±102.9 cells/mm2 (PBS; Fig. 3G, bottom), whereas in animals injected with the hAFS cells (Fig. 3E), the apoptotic active caspase-3-positive cells in the myocardium were 483.2±82 cells/mm2 (hAFS; Fig. 3G, bottom) and in the group receiving the hAFS-CM (Fig. 3F) it was 245±37 cells/mm2 (hAFS-CM; Fig. 3G, bottom). Rats treated with either hAFS cells or the hAFS-CM upon reperfusion showed a significantly reduced level of cell death in the acute phase following injury, compared with the PBS control group (****P<0.001). The 2 treatments were equally effective in improving myocardial cell survival (P>0.05).
In vitro vascular potential of hAFS cells
In vitro cultured hAFS cells (Fig. 4A) did not express any endothelial markers (Fig. 4B, C). A proportion of them were positive for the expression of α-SMA, as detected by immunostaining (about 61%±13%; Fig. 4D–F), showing a partial commitment to the smooth muscle lineage.
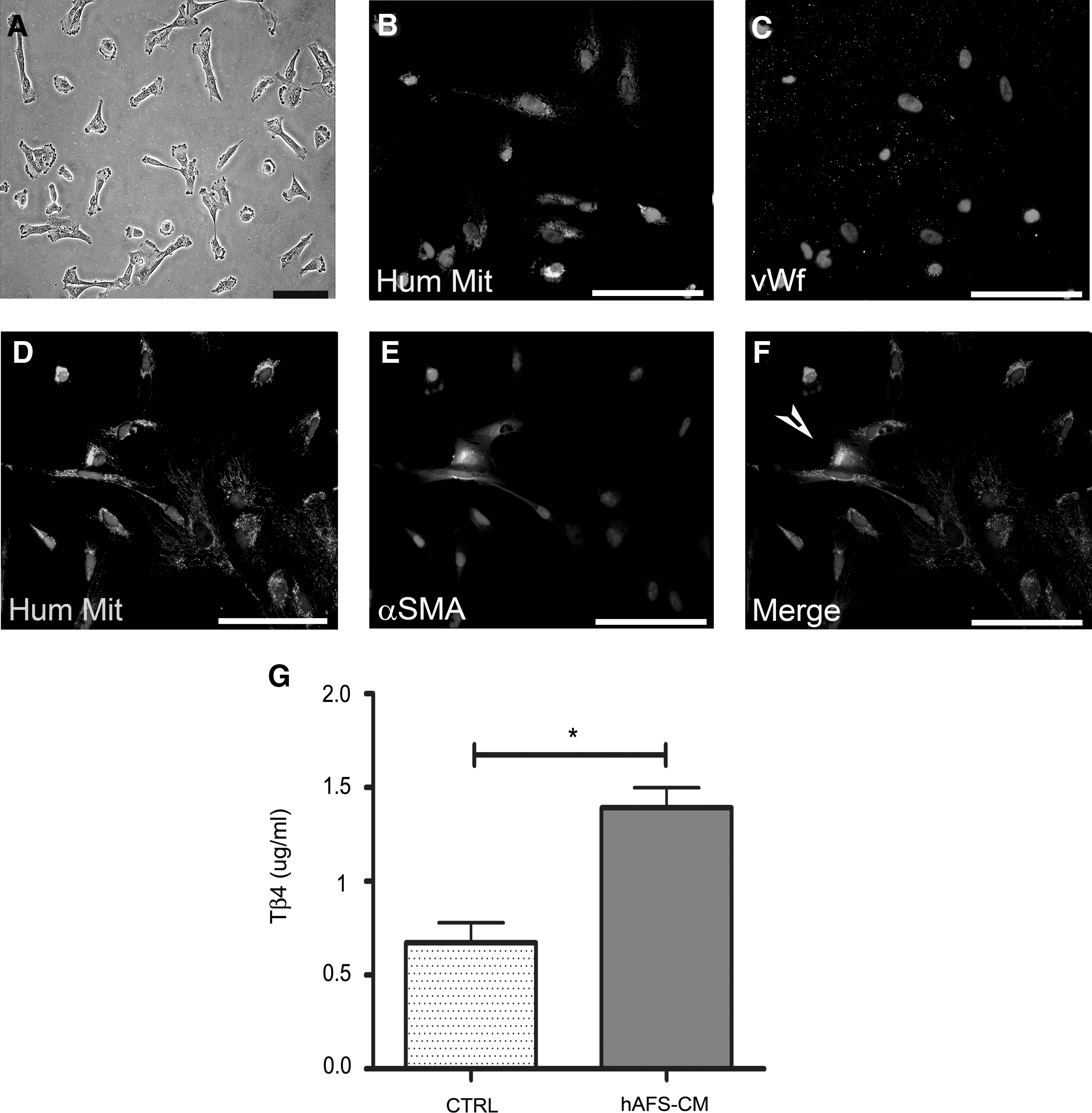
In vitro characterization of hAFS cells and analysis of Tβ4 production in the culture medium. hAFS cells
Tβ4 secretion in the culture medium of hAFS cells
hAFS cells were shown to secrete Tβ4 into the medium. The untreated culture medium alone contained a basal level of Tβ4 (control, 0.67±0.11 μg/mL; Fig. 4G), and hAFS cells-CM showed a significant increase in the concentration of Tβ4 (hAFS-CM, 1.39±0.10 μg/mL, *P<0.05; Fig. 4G).
Discussion
In this study, we demonstrate for the first time that, in an acute myocardial ischemia/reperfusion injury model, the i.v. systemic administration of hAFS cells, at the beginning of reperfusion, results in a significant decrease in the IS and a reduced number of apoptotic cardiomyocytes.
Considering the small quantity of hAFS cells engrafted in vivo, the rapidity of their beneficial effect (over a period of 2 h), and given the finding that hAFS cells secrete the cardioprotective factor Tβ4, we hypothesize that this result may be explained primarily via an in situ paracrine effect.
Reperfusion is a critical time for cardiomyocyte survival as well as the release of growth factors that can locally act to benefit the injured cells [19 –22]. Although the transplanted cells may work through different mechanisms [3], the demonstration that effects on ventricular function and myocardial perfusion acutely occur and do not require cell differentiation supports the idea of a paracrine action [20,23]. These mechanisms may include proangiogenic stimulation, stem cell mobilization, stabilization of the extracellular matrix, and activation of resident cardiac stem and progenitor cells [24]. In this regard, BM-MSCs have been shown to improve heart function, decrease collagen volume fraction, and increase expression of adrenomedullin, an antifibrotic factor, in the myocardium [25]. This protective mechanism has been also shown during ischemia in other organs, such as brain [26] and kidney [27]. More recently, BM-MSCs-conditioned medium (CM) was also shown to exert protective effects on endogenous cardiac progenitor cells [28].
Although it is likely that the paracrine effect plays the most relevant role in any donor cell-based protection during ischemia-reperfusion, it is still an open question as to the specific identity of the key secreted factor(s).
In this study, we mainly focused on Tβ4—a potent stimulator of coronary vasculogenesis and angiogenesis and of cardiomyocyte survival following cardiac injury [10,12 –14]—as the most interesting paracrine factor to explain the associated IS reduction following hAFS cells and hAFS-CM injection. Indeed, Tβ4 has been demonstrated to be one of the most upregulated paracrine-acting agents present in the conditioned medium of BM-MSCs overexpressing Akt, capable of restoring cardiac function and inhibiting ventricular remodeling in a rat myocardial infarct model [29,30]. Regarding the detailed mechanism behind the cardioprotective effect mediated by Tβ4, despite the extensive identification of its multiple biological roles, little progress has been made in identifying the comprehensive signaling beyond its beneficial effects on the ischemic heart. Bock-Marquette et al. reported the significant role of intracardiac and intraperitoneal administration of Tβ4 in promoting survival of cardiomyocytes after myocardial infarct in mice, possibly involving the integrin-linked kinase and Akt pathways [10]; Hinkel et al. showed that the cardioprotective potential of embryonic endothelial progenitor cells in a model of cardiac ischemia-reperfusion can be attributed, at least in part, to Tβ4 [13]. In addition, Zhao et al. recently reported that ILK/PI3K/Akt, as well as MAPK signaling, seems to be the dominant mechanism through which Tβ4 promotes endothelial precursor cell survival under in vitro serum deprivation conditions [31]. In our experimental setting, i.v. administration of hAFS cells diminished the area of infarct by maintaining myocardial cell survival and reducing apoptosis in the acute phase following injury (Fig. 3G), possibly through a paracrine mechanism. We envisage that this may be mediated by the secretion of Tβ4 and, indeed, this fits with an early role for Tβ4 after injury [10].
Our assumption seems to be also confirmed by the fact that the treatment with hAFS-CM alone—that is enriched with Tβ4 as demonstrated by the in vitro analysis—provided equivalent results in decreasing the IS in the short term. Indeed, the 2 treatments appear to be comparable in terms of improving the survival of the cardiac tissue within the reperfusion time, as supported by our analysis of apoptosis. Hence, hAFS cells seem to be able to mediate a significant paracrine effect on the ischemic heart by releasing Tβ4, which might locally act in the cardiac tissue potentially supporting the activation of resident progenitors [12,28]. Although hAFS cells can secrete considerable amounts of Tβ4 in their medium, we did not detect any increase of the protein concentration in the blood plasma at the end of reperfusion, either in the animals injected with cells or in those treated with the conditioned medium (data not shown). Indeed this may not be entirely unexpected, given the very short length of time between the treatment and the analysis, which might not be sufficient to develop an effect detectable systemically.
Interestingly, our findings demonstrating the paracrine potential of hAFS cells seem to be in line with the current trend in regenerative medicine, as human amniotic fluid-derived stem cells have been recently shown to secrete angiogenic factors that may account for the neoarteriogenic effect in a hind-limb ischemic model. Indeed, in that work, similarly to ours, AFS cells-CM was shown to be enriched in soluble mediators, such as MCP-1, IL-8, SDF-1, and VEGF, and its administration improved the outcome of the ischemic injury [32].
In our cell transplantation model we used in vitro expanded hAFS cells, which were previously cryopreserved. AFS cells are easy to isolate, expand, and preserve, maintaining their proliferative and differentiation potential, following long-term culture and freeze–thawing cycles [8,33]. Conversely, BM-MSCs, although they similarly demonstrated comparable remarkable cardioprotective effects in the short term (as shown in a model of myocardial I/R injury in a comparable study [34]), when utilized after prolonged culture, failed to invoke any improvement in cardiac function up to 6 weeks after cell transplantation [35]. Therefore, from a clinical translation perspective, AFS cells can be considered a novel and attractive source for both cell and protein therapies (the latter considering the paracrine soluble factors they release). Specifically, hAFS cells produce high levels of Tβ4, a significant paracrine cardioprotective agent and activator of resident neovascular progenitors. Further, hAFS cells could have regenerative potential in the long term, as they have been shown to possess broad differentiation potentials. We and others have demonstrated that hAFS cells can differentiate into cardiovascular lineages, including endothelial, smooth muscle cells, and cardiomyocytes in vitro [8,33,36 –38] and in vivo over the long term [38], and can be harvested under defined conditions [9]. Moreover, amniotic membrane-derived hMSCs have been also shown to possess a remarkable cardiomyogenic potential in vivo [39], thus confirming how amniotic fluid and amnion tissue may represent a new font for allograftable stem cells for cardiac regenerative medicine.
In this work, our hypothesis is that early injection of hAFS cells might provide a significant benefit after ischemia-reperfusion injury via the release of paracrine factors, including Tβ4. Here, we focused on evaluating their acute cardioprotective influence in the very short term (2 h after the coronary artery occlusion), rather than their regenerative effect in terms of providing de novo cardiomyogenesis with creation of new functional cells, a process that may require days.
In our work, hAFS cells were systemically injected to assess their ability of homing to the target tissue. Because of the route of administration, it was not surprising that the injected hAFS cells were also found in tissues other than the heart, including the spleen and lungs. hAFS cells were shown to partially engraft in the heart, expressing markers of the endothelial and smooth muscle lineages. Moreover, in our study, undifferentiated hAFS cells have been shown to express smooth muscle markers, in accordance with what was previously reported [36]; therefore, hAFS cells seem to possess a (cardio)vascular immature potential in the undifferentiated state, indicative of a partial commitment that can be driven to more complete maturation by the environmental stimuli received once injected and engrafted in the specialized host tissue. At the same time, as the hAFS cells costained for endothelial and smooth muscle markers were found in close relationship with the endothelial and smooth muscle cells from the host, we cannot exclude that they might have fused with the some of the resident cells, generating chimeric vessels.
Further, although in our acute model we could not investigate the effect of the hAFS cells on the host immune system, these cells have been shown to express proteins and genes of major histocompatibility complex (MHC) class I, but not those of MHC class II [40]; they have also demonstrated an immunosuppressive potential on induced T-cell proliferation in vitro and to be resistant to rejection, expressing immunomodulatory factors such as CD59 and HLA-G [41]. Our group has demonstrated that, although AFS cells have the propensity to acquire a cardiomyogenic phenotype in vitro [36,38] and preserve cardiac function in vivo, their potential may be limited by poor survival in the allogeneic setting in the long term [38]. Hence, the significance of AFS cells in cardiac regenerative medicine might be more relevant in the autologous setting. Indeed, from a clinical translation perspective, AFS cells might represent an attractive source of autologous stem cells to be isolated during pregnancy, cryopreserved, and banked for use when needed.
In conclusion, the data presented here support the idea that hAFS cells potentially are a very promising and useful source for the treatment of AMI as they represent a very appealing stem resource and a font of cardioprotective soluble factors.
Footnotes
Acknowledgments
The authors acknowledge the support of the British Heart Foundation, the Engineering and Physical Sciences Research Council, and the Biotechnology and Biological Sciences Research Council. S.B. was supported by Città Della Speranza, Malo (VI), Italy, and P.D.C. was funded by Great Ormond Street Hospital Children's Charity, United Kingdom.
Author Disclosure Statement
No competing financial interests exist.