
Abstract
Chronic idiopathic neutropenia (CIN) is a granulopoiesis disorder associated with an inhibitory bone marrow (BM) microenvironment consisting of activated T-lymphocytes and pro-inflammatory mediators. In this study, we investigated the possible involvement of BM mesenchymal stem cells (MSCs) in the pathophysiology of CIN by assessing the frequency and function of BM MSCs in terms of the proliferative/clonogenic characteristics, the differentiation capacity, the potential to produce pro-inflammatory cytokines, and the ability to suppress T-cell proliferation. The frequency, differentiation capacity toward adipocytes, chondrocytes, or osteoblasts, and immunosuppressive potential to inhibit mitogen-induced T-cell proliferation did not differ significantly between patient (n = 14) and normal (n = 21) MSCs. Tumor necrosis factor-α, interleukin-1β, and interleukin-6 levels in MSC supernatants did not differ significantly between patients and controls; however, transforming growth factor (TGF)-β1 levels were significantly elevated in patients, particularly in those displaying the -509C/T TGF-β1 polymorphism. Patient MSCs displayed defective proliferative/clonogenic potential, which could not be attributed to altered cellular survival characteristics or to increased TGF-β1 production as TGF-β1 neutralization did not restore the impaired colony formation by patient MSCs. We conclude that although BM MSCs do not exert a significant role in the immune deregulation associated with CIN, they contribute to the inhibitory microenvironment by overproducing TGF-β1, at least in patients displaying the -509C/T polymorphism.
Introduction
C
Although CIN has long been considered as an “orphan” disease with unknown pathogenesis, there is now strong evidence suggesting that the impaired neutrophil production in this disease is mainly due to the inhibitory effect of the BM microenvironment consisting of activated T-lymphocytes, pro-apoptotic cytokines, and pro-inflammatory mediators such as tumor necrosis factor (TNF)-α, transforming growth factor (TGF)-β1, Fas-ligand, interferon-γ, interleukin (IL)-1β, and IL-6 [3 –5]. Interestingly, it has been shown that stromal layers from long-term BM cultures of patients with CIN, a culture system that mimics the BM microenvironment, fail to support autologous and normal hematopoiesis in cross-over experiments, [1] whereas in vitro neutralization of the inhibitory cytokine activity may restore parameters of hematopoiesis in these patients [6,7]. Therefore, CIN can be considered as a BM failure syndrome associated with a defective BM microenvironment.
The marrow microenvironment is a complex network of extracellular matrix and heterogeneous cellular components of hematopoietic and mesenchymal origin that sustain the growth, proliferation, and differentiation of hematopoietic stem cells and their progeny throughout life [8]. The mesenchymal cell population originates from a rare population of multipotential progenitor cells, currently referred to as mesenchymal stem cells (MSCs) [9,10]. Because MSCs display important immunoregulatory and immunosuppressive properties along with their hematopoiesis-supporting effects [11 –13], there is currently a particular interest in exploring the possible primary involvement of BM MSCs in clonal and immune-mediated hematologic diseases associated with BM microenvironment defects such as myelodysplastic syndromes and aplastic anemia [14 –16]. Data, however, are still limited and sometimes contradictory [17 –20].
The possible role of MSCs in BM failure associated with CIN has never been studied and the question as to whether MSCs in CIN differ from their normal counterparts remains open. In the present study, we have investigated the reserves and qualitative characteristics of BM MSCs in CIN patients assessing specifically the proliferative and clonogenic potential, the differentiation capacity, and the immunomodulatory properties in terms of their ability to suppress the T-cell proliferative responses. Identification of intrinsic alterations of MSCs might imply a primary BM microenvironment defect in CIN that could contribute to the pathogenesis/pathophysiology of the disease.
Patients and Methods
Patients
We studied 14 patients with CIN, 11 females and 3 males (age: 17–69 years; median age: 47 years), satisfying the previously defined diagnostic criteria for the disease [1]. In particular, the patients had neutrophil counts <1800 × 106 cells/L (mean: 881 ± 414 × 106 cells/L; range: 300–1400 × 106 cells/L) for a period of 18–168 months (median duration: 36 months), no clinical, serological, or ultrasonic evidence of any underlying disease known to be associated with neutropenia, no history of exposure to irradiation, no use of chemical compounds or intake of drugs to which neutropenia might be ascribed, normal BM karyotype, and negative serum leucoagglutination and immunofluorescence tests for antineutrophil antibodies. Detailed patient characteristics are presented in Table 1. As controls, 21 hematologically healthy subjects, age- and sex-matched with the patients, were studied. Institutional ethics committee approval was granted prior to the study and informed consent according to the Declaration of Helsinki was obtained from all subjects studied.
UPN, unique patient number; Hb, hemoglobin; WBCs, white blood cells; Neutro, neutrophils; Lympho, lymphocytes; Mono, monocytes; Plts, platelets.
MSC cultures
BM cells from posterior iliac crest aspirates were diluted 1:1 in Dulbecco's modified Eagle's medium-low glucose (DMEM-low glucose; Gibco Invitrogen) supplemented with 100 IU/mL penicillin–streptomycin (Gibco) and 10 IU/mL preservative-free heparin (Sigma). The BM mononuclear cells (BMMCs) were obtained following centrifugation on Histopaque-1077 (Sigma) and were cultured in DMEM-low glucose/10% fetal calf serum (FCS; Hyclone)/100 IU/mL penicillin–streptomycin (MSC medium) at a concentration of 2 × 105 cells/cm2 in 25-cm2 culture flasks at 37°C/5% CO2 fully humidified atmosphere. One to 3 days postseeding, nonadherent cells were removed, and thereafter, the medium was replaced every 3–4 days. On reaching 70%–90% confluence, the cells were detached using 0.25% trypsin–1 mM ethylenediaminetetraacetic acid (Gibco) and reseeded for a total of six passages (P). Cell-free supernatants were stored at −70°C for measurement of TNF-α, IL-1β, IL-6, and TGF-β1 by an enzyme-linked immunosorbent assay. All enzyme-linked immunosorbent assay kits were purchased from R&D Systems (Quantikine) except the TNF-α kit (Biosource International).
Flow cytometric analysis of BM
Trypsinized MSCs from P1 to P6 were immunophenotypically characterized by flow cytometry using anti-human monoclonal antibodies (mAb) against CD29 (4B4; Cyto-Stat/Beckman-Coulter), CD44 (J173; Immunotech/Coulter), CD73 (AD2; Becton Dickinson-Pharmingen), CD90 (F15.42; Immunotech/Coulter), CD105 (SN6; Caltag), anti-CD146 (P1H12; Becton Dickinson-Pharmingen), CD45 (IMMU19.2; Immunotech/Coulter), CD14 (RMO52; Immunotech/Coulter), and CD34 (QBend10; Beckman-Coulter). The expression of the Ki-67 nuclear antigen (clone Ki-67; Dako Denmark A/S, Glostrup) as an index of cell proliferation [21] was assessed in MSCs from two representative passages (P1 and P2) using the IntraPrep permeabilization reagent (Beckman-Coulter) according to the manufacturer's instructions. Finally, flow cytometry was used to study the survival/apoptotic characteristics of MSCs through P1–P6 by means of 7-aminoactinomycin D staining (7-AAD; Calbiochem-Novabiochem) as previously described [22]. Results were expressed as proportions of 7-AADneg (live), 7-AADdim (early apoptotic), and 7-AADbright (late apoptotic) cells. Flow cytometry data were processed using an Epics Elite model flow cytometer (Coulter).
MSC differentiation assays
Trypsinized MSCs from P2 were induced for adipogenic, osteogenic, and chondrogenic differentiation as previously described [23]. Adipogenic differentiation was induced in MSC medium supplemented with 10% FCS/0.5 mM 1-methyl-3-butyl isoxanthine/1 μM dexamethasone (Dex)/0.2 μM indomethacin/10 μg/mL insulin. Lipid formation was assessed by Oil Red O staining [24]. Osteogenic differentiation was induced in MSC medium supplemented with 0.1 μM Dex/0.15 mM ascorbate-2-phosphate/3 mM NaH2PO4. Differentiation was assessed by alkaline phosphatase (ALP)/von Kossa staining [24]. For chondrogenic differentiation, MSCs were pelleted in 15-mL tubes and cultured in DMEM-high glucose (Gibco) without FCS, supplemented with 6.25 μg/mL insulin, 6.25 μg/mL transferrin, 1.33 μg/mL linoleic acid, 1.25 mg/mL bovine serum albumin, 1 mM sodium pyruvate, 0.17 mM ascorbate-2-phosphate, 0.1 μM Dex, 0.35 mM
Total RNA isolated from differentiated MSCs from patients and controls (RNeasy mini kit; Qiagen GmbH) was reverse transcribed (SUPERSCRIPT II; Gibco) and amplified by polymerase chain reaction (reverse transcriptase–PCR) for the evaluation of specific, differentiation-associated gene expression: adipose fatty acid-binding protein and peroxisome proliferator activated receptor-γ for adipocytes, ALP and runt-related transcription factor 2 for osteocytes, and collagen type II (COL2A1) and aggrecan (AGC1) for chondrocytes. Products were normalized according to β2-microglobulin (β2m) expression, using the ImageJ densitometry analysis system. Primer sequences and reverse transcriptase–PCR conditions have been reported previously [25].
Quantification of MSCs in the BMMC fraction
A limiting dilution assay (LDA) was used to evaluate the frequency of MSCs within the BMMC fraction following 6-week culture of seven different concentrations of MSCs (250–10,000 cells/well) as previously described [26,27]. Wells with >50 adherent, spindle-shaped cells were considered positive and MSC frequency corresponded to the dilution that resulted in 37% negative wells [28].
Clonogenic potential of MSCs
A colony-forming unit fibroblast (CFU-F) assay was used for the evaluation of the clonogenic potential of MSCs through P1–P6. Trypsinized MSCs from P1 to P6 were seeded at three different concentrations (50–150 cells) in 60-mm plates for 14 days. CFU-Fs were quantified using Giemsa staining [24]. The number of CFU-F was estimated per 100 MSCs on the basis of the linear regression analysis obtained from the three different initial cell concentrations [27,29]. In a set of experiments (n = 5), patient MSCs from P2 were assayed for CFU-F formation in the presence or absence of two different concentrations (1 and 4 μg/mL) of a mouse anti-human TGF-β1 neutralizing mAb (50 μg/mL; R&D Systems). According to the manufacturer, approximately 0.3–1 μg/mL of the mAb neutralizes 50% of the bioactivity due to 0.25 ng/mL of recombinant human TGF-β1.
Proliferative potential of MSCs
The proliferative potential of MSCs was evaluated by a methyl triazolyl tetrazolium (MTT)-based assay at P2 and also by estimating the population doubling time through P1–P6 [27]. The formula 2 n = Nx /N 0 was used for the calculation of the population doublings (n) at each passage based on the number of cells counted in the flask after trypsinization (Nx ) and the number of cells initially plated (N 0).
MSC inhibition of T-cell proliferation
To evaluate the capacity of MSCs from CIN patients to suppress T-cell proliferative responses compared with normal subjects, T-lymphocytes were mixed with MSCs in the absence or presence of various stimuli [11,14,30]. For T-cell isolation, peripheral blood samples from healthy donors were centrifuged on Histopaque-1077 to obtain the MCs and the CD3+ cell fraction was fractionated by indirect magnetic labeling (magnetic activated cell sorting; MACS isolation kit; Miltenyi Biotec GmbH) according to the manufacturer's protocol. In each experiment, purity of CD3+ cells was greater than 96% as estimated by flow cytometry. Subsequently, 5 × 104 immunomagnetically sorted CD3+ cells were stimulated with phytohemagglutinin (PHA; 2 μg/mL) or IL-2 (500 IU/mL) in the presence or absence of 104 irradiated (30 Gy) BM MSCs from CIN or allogeneic healthy controls in V-bottomed 96-well culture plates for 7 days in 0.2 mL RPMI-1640 medium (Gibco) containing 10% FCS. T-cell proliferation was measured on day 7 following an 18-h pulse with 1 μCi/well 3H-thymidine (3H-TdR; Amersham). 3H-TdR incorporation was measured by using a liquid scintillation counter (LS1701 beta counter; Beckman) and expressed as counts per minute (cpm). Experiments were performed in triplicates.
PCR–restriction fragment length polymorphism assay
Genomic DNA was extracted from peripheral blood samples of CIN patients using the QIAamp Blood Mini Kit (Qiagen). Genotype analysis for the detection of the -509C/T hypersecretory polymorphism of TGF-β1 was performed in peripheral blood DNA samples (QIAamp Blood Mini Kit; Qiagen) from CIN patients using a PCR-based restriction fragment length polymorphism assay as previously described [31].
Statistical analysis
Data were analyzed by means of the nonparametric Mann–Whitney test and the one- and two-way analyses of variance tests (GraphPad Software). Grouped data were expressed as means ± 1 SD.
Results
Frequency, morphologic and immunophenotypic characteristics of BM MSCs
MSC cultures were successfully expanded in all CIN patients and healthy controls until P6. Adherent cells from both groups displayed the characteristic spindle-like morphology, and immunophenotypic analysis at the end of each passage (P1–P6) demonstrated that cultures constituted of a homogeneous cell population expressing CD73, CD90, CD146, CD105, CD29, and CD44 surface antigens but lacking the hematopoietic markers CD45, CD14, and CD34 (Fig. 1A). The estimated frequency of MSCs in the BMMC fraction by means of LDA assay after a culture period of 6 weeks did not differ significantly between CIN patients (21.28 ± 3.67 per 105 BMMCs) and healthy controls (23.90 ± 15.65 per 105 BMMCs; P = 0.383) (Fig. 1B). These data suggest that the reserves and the morphologic and immunophenotypic characteristics of BM MSCs in CIN patients are similar to the normal individuals.
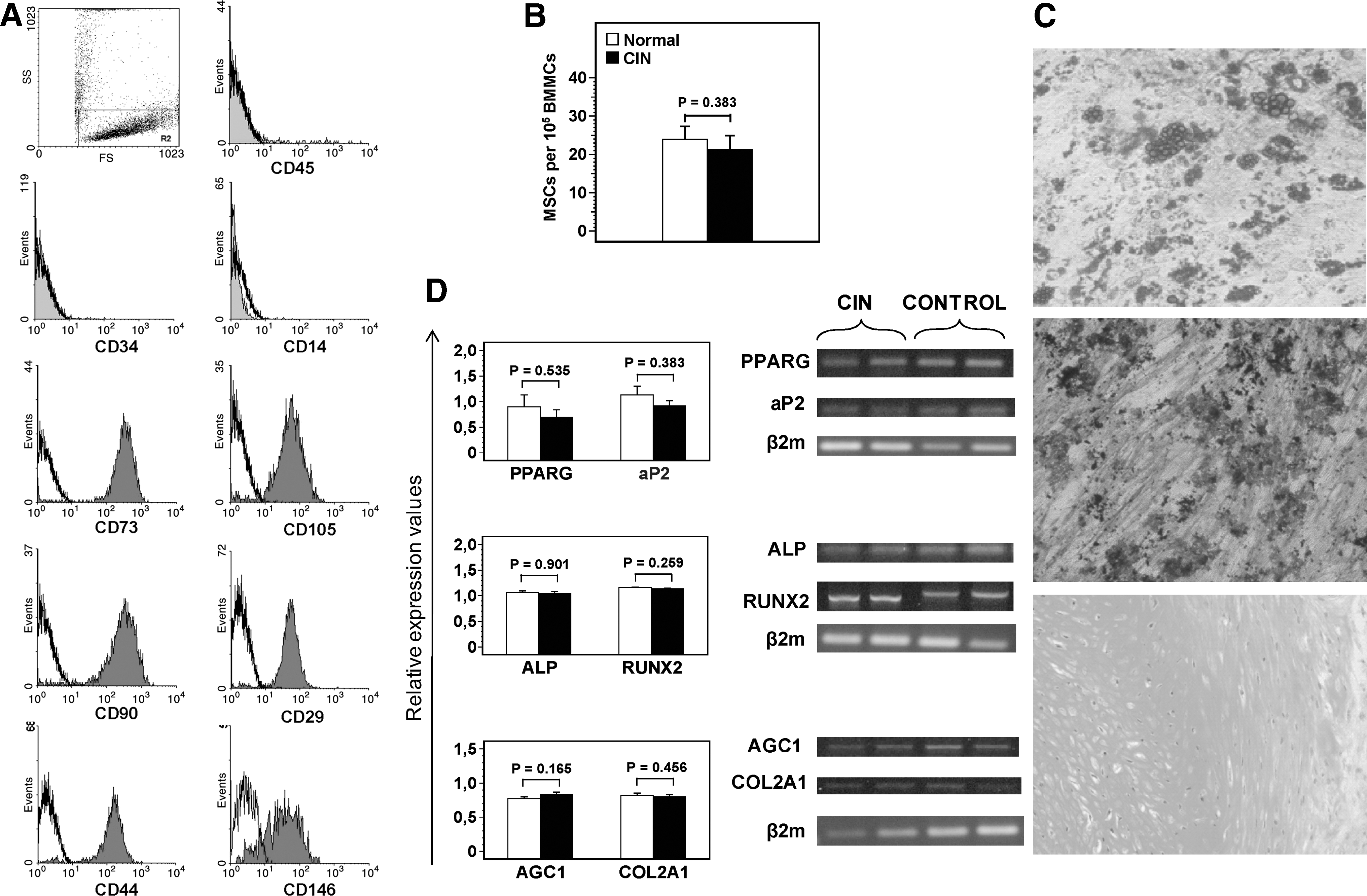
Immunophenotypic characteristics, reserves, and differentiation potential of MSCs in CIN.
Differentiation potential of MSCs
To assess the differentiation potential of MSCs in CIN patients, we cultured P2 MSCs in standard induction media and explored subsequently their capacity to generate cells of adipogenic, osteogenic, and chondrogenic lineages. Respective cytochemical staining demonstrated that the CIN MSCs tested (n = 7) were able to differentiate into adipocytes, osteocytes, and chondrocytes and this property was not qualitatively different from that of normal MSCs (n = 7) (Fig. 1C). Similarly, the adipogenic, osteogenic, and chondroblastic capacities of CIN MSCs were not quantitatively different from that of normal MSCs, as shown by the relative mRNA expression of the adipose fatty acid-binding protein and peroxisome proliferator activated receptor-γ (adipogenic markers), ALP and runt-related transcription factor 2 (osteogenic markers), and COL2A1 and AGC1 (chondrogenic markers) (Fig. 1D). These data suggest that BM MSCs from CIN patients display normal differentiation potential, at least toward the three lineages tested.
Clonogenic and proliferative potential of MSCs
To evaluate the growth capacity of BM MSCs in CIN patients, we assessed their clonogenic and proliferative potential over time. The results are shown in Fig. 2. CFU-F recovery by culture-expanded MSCs was significantly lower in patients compared with normal controls through the P0–P6 culture period (F = 6.049, P = 0.0149), suggesting decreased clonogenic potential of BM MSCs in CIN. In keeping with the defective clonogenic capacity, patient MSCs displayed significantly increased doubling time throughout passages (F = 100.3, P < 0.0001). Specifically, the MSC doubling time ranged from 3.67 ± 1.06 (P1) to 8.73 ± 4.67 (P5) days in patients and from 2.29 ± 0.82 (P1) to 4.35 ± 0.84 (P5) days in controls. This observation was further substantiated by the MTT assay according to which the number of live cells, corresponding to the obtained optical density, at a representative passage (P2) remained significantly lower in CIN patients compared with controls (F = 29.89, P < 0.0001). The lower proportion of MSCs expressing the Ki-67 nuclear antigen in two representative passages (P1 and P2) in patients (33.78% ± 19.18% and 45.18% ± 24.83%, respectively) compared with controls (61.79% ± 18.97% and 65.84% ± 12.73%, respectively; P = 0.0006 and P = 0.011, respectively) corroborates further the option that BM MSCs in CIN display decreased cycling and proliferative potential compared with their normal counterpart.
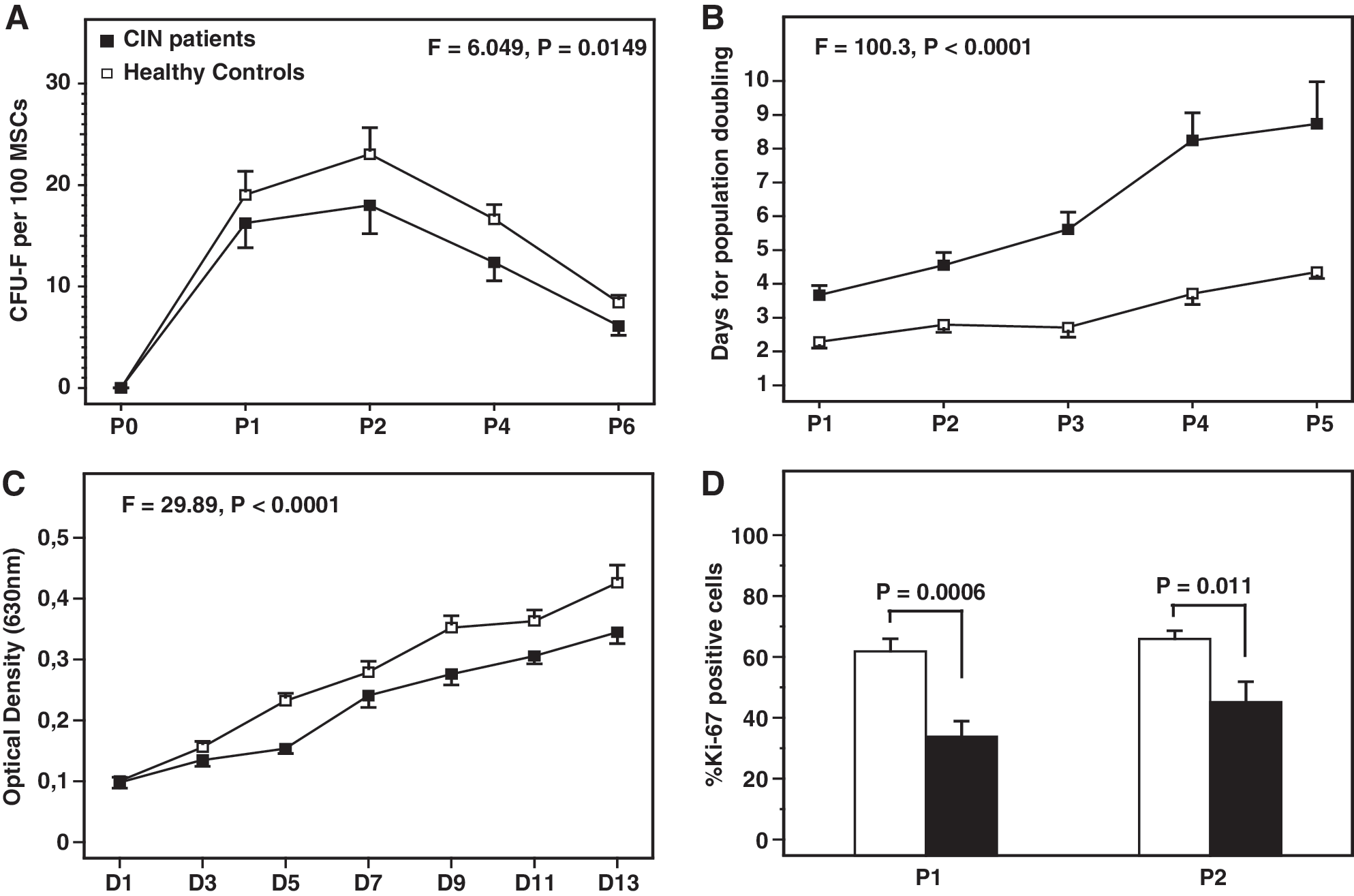
Clonogenic and proliferative potential of MSCs time-course.
Survival characteristics of MSCs
To probe the underlying cause for the impaired growth potential of BM MSCs in CIN, we studied the survival characteristics of cells over the P1–P6 culture period (Fig. 3). No statistically significant differences were found between patients and controls in the proportion of early (F = 0.6869, P = 0.4087) or late apoptotic (F = 0.127, P = 0.7222) cells throughout the P1–P6 culture period, suggesting that the impaired clonogenic and proliferative potential of MSCs in CIN is not due to accelerated cell apoptosis.
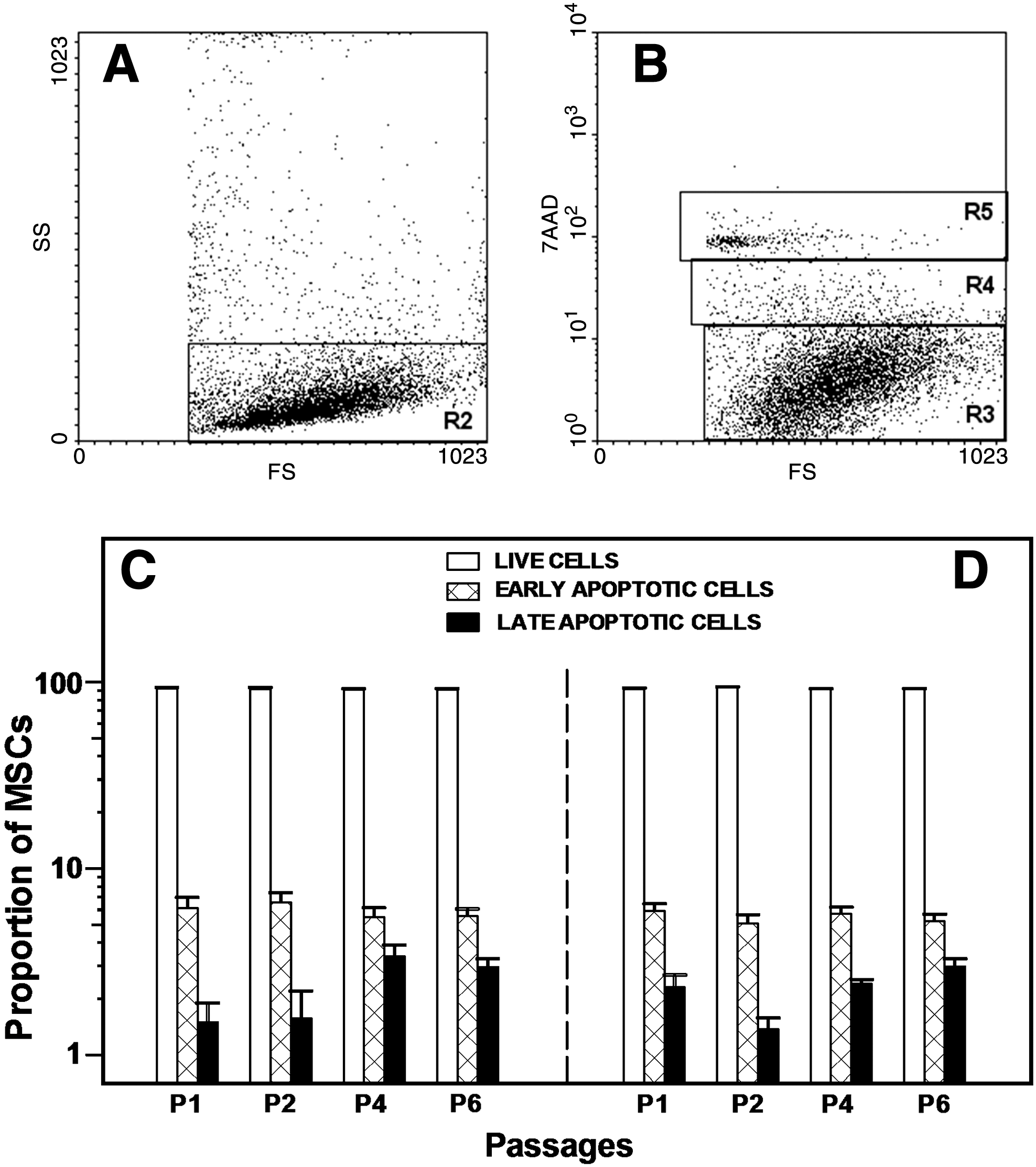
Survival characteristics of MSCs. MSCs from P1 to P6 were stained with 7-AAD and analyzed by flow cytometry for the study of apoptosis. Scattergram
Immunosuppressive properties of MSCs
The effect of patient MSCs on T-cell proliferation was evaluated by mixing normal or patient MSCs with allogeneic normal T-lymphocytes, stimulated to proliferate with PHA or IL-2. T-cell proliferation was measured by 3H-TdR incorporation following a 7-day culture and the results are depicted in Fig. 4. As anticipated, T-cell proliferation was significantly reduced when PHA– or IL-2–stimulated T-lymphocytes (4414 ± 1829 and 30,118 ± 26,379 cpm, respectively) were cocultured with MSCs from normal donors (644 ± 440 and 3296 ± 3141 cpm, respectively; P = 0.0022 and P = 0.0087, respectively). Similarly, T-cell proliferation following PHA or IL-2 stimulation (5845 ± 3747 and 20,390 ± 24,747 cpm, respectively) was significantly suppressed in the presence of MSCs from CIN patients (710 ± 297 and 2158 ± 3373 cpm, respectively; P = 0.0022 and P = 0.0152, respectively). The percentage of inhibition of PHA– or IL-2–induced T-cell proliferation by MSCs did not differ significantly between patients (84.61% ± 9.57% and 85.26% ± 10.24%, respectively) and controls (88.55% ± 3.83% and 89.05% ± 2.14%, respectively; P = 0.485 and P = 0.309, respectively). These findings suggest that MSCs from CIN patients display normal immunosuppressive properties in terms of the capacity to suppress T-cell proliferative responses.
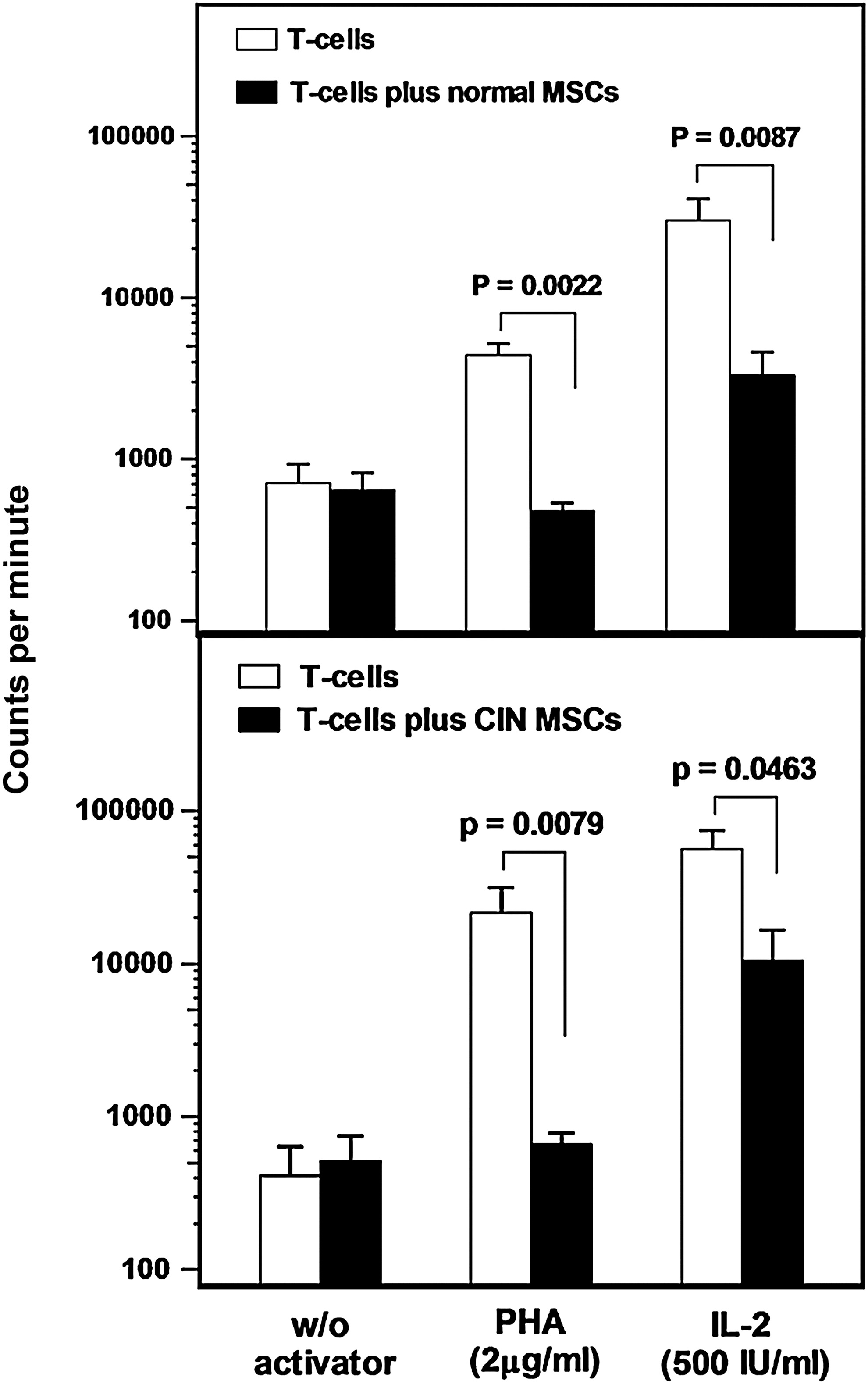
Inhibitory effect of MSCs on T-lymphocyte proliferation. The bars represent the proliferative responses (expressed in counts per minute upon 3H-TdR incorporation) of unstimulated and activator-induced T-lymphocytes in the presence or absence of BM MSCs from allogeneic normal (upper panel) or patient (lower panel) MSCs. Data are expressed as mean (±SEM) of triplicates of six separate experiments, respectively. Comparison of counts per minute in the presence or absence of MSCs was performed by means of the nonparametric Mann–Whitney test.
Cytokine production by MSCs
To investigate whether BM MSCs are the primary source of the pro-inflammatory cytokines known to be associated with the pathophysiology of CIN, we assessed TNF-α, IL-1β, IL-6, and TGF-β1 levels in culture supernatants from P2, P4, and P6 (Table 2). No statistically significant difference was found between patients and control MSCs in the production of TNF-α, IL-1β, and IL-6 time-course (P = 0.7853, P = 0.1391, and P = 0.4228, respectively) (Table 2). Interestingly, however, TGF-β1 levels were found to be significantly elevated in MSC cultures from CIN patients compared with healthy individuals throughout the culture period (F = 19.52, P < 0.0001).
Values are expressed as means ± SD. Median and range are indicated in parentheses.
Comparison between patients and controls regarding the cytokine levels in culture supernatants through the P2–P6 culture period has been performed by means of the two-way ANOVA.
TNF, tumor necrosis factor; CIN, chronic idiopathic neutropenia; TGF, transforming growth factor.
Our previous studies have shown that the -509C/T polymorphism of TGF-β1 is identified at a higher frequency in CIN patients compared with normal individuals [32]. We have also shown that CIN patients with this particular polymorphism display increased levels of the cytokine in serum and long-term bone marrow culture (LTBMC) supernatants [32]. To investigate whether the -509C/T polymorphism has also a role in the increased TGF-β1 production by patient MSCs, we analyzed cytokine levels on the basis of -509C/T TGF-β1 genotype and the results are shown in Fig. 5A. Genotype data of this group of CIN patients have been reported previously [32]. We found that the group of CIN patients with the mutant CT (n = 7) or TT (n = 3) genotype displayed significantly increased levels of TGF-β1 in P2, P4, and P6 MSC culture supernatants (1946 ± 1085, 1195 ± 1024, and 652 ± 277 pg/mL, respectively) compared with patients with the wild-type CC genotype (n = 4) (678 ± 227, 327 ± 79, and 233 ± 51 pg/mL, respectively; P = 0.002, P = 0.002, and P = 0.008, respectively) or the healthy controls (858 ± 273, 422 ± 324, and 266 ± 206 pg/mL, respectively; P = 0.0004, P = 0.0029, and P = 0.0005, respectively). Notably, no statistically significant difference was found between CIN patients displaying the wild-type CC genotype and healthy individuals in the TGF-β1 levels detected in MSC culture supernatants through P2–P6.
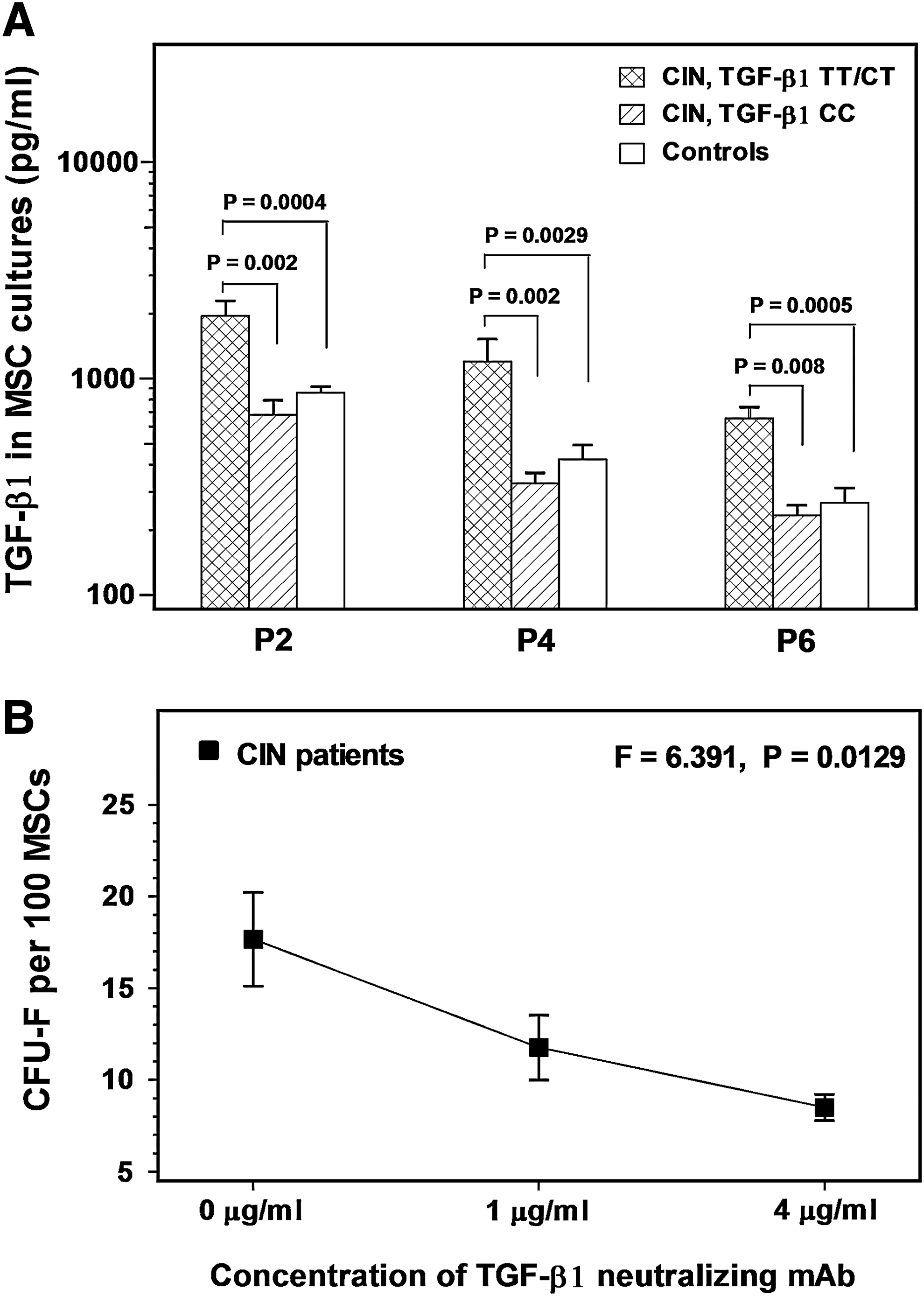
TGF-β1 levels in MSC culture supernatants and the effect of TGF-β1 neutralization on the clonogenic potential of BM MSCs.
The effect of TGF-β1 neutralization on the clonogenic potential of MSCs
To investigate whether the increased endogenous production of TGF-β1 might account for the impaired clonogenic potential of patient MSCs, we assessed the CFU-F formation by P2 MSCs in the presence of 1 and 4 μg/mL of a TGF-β1 neutralizing mAb (Fig. 5B). Interestingly, we found a dose-dependent decrease in the number of CFU-F upon TGF-β1 neutralization (11.76 ± 3.96 and 8.50 ± 1.58 CFU-F per 100 MSCs, respectively) compared with baseline untreated cultures (17.67 ± 5.70 CFU-F per 100 MSCs; F = 6.391, P = 0.0129), suggesting a stimulatory rather that an inhibitory effect of TGF-β1 on the clonogenic potential of BM MSCs.
Discussion
The impaired neutrophil production in patients with CIN has been clearly associated with an abnormal BM microenvironment exerting a growth inhibitory effect on the granulocytic progenitor cells [1,4,5]. Whether this abnormality originates or affects the BM MSCs has not been investigated thus far. The present study aimed therefore at assessing the reserves and the phenotypic and functional characteristics of culture-expanded BM MSCs in CIN patients.
The frequency of MSCs assessed by an LDA in the BMMC fraction did not differ significantly between patients and normal subjects, suggesting that CIN patients exhibit normal BM MSC reserves. In addition, culture-expanded MSCs from CIN patients displayed normal morphologic and immunophenotypic characteristics and exhibited normal differentiation potential toward the adipogenic, osteogenic, and chondrogenic lineages as evidenced by the respective cytochemical staining and the quantitative mRNA expression of differentiation-specific markers.
MSCs have been shown to possess diverse immunomodulatory properties such as inhibition of T-cell proliferation in response to alloantigens and nonspecific mitogens, thereby providing an immunoprotective environment to hematopoietic progenitor cells. [11] Interestingly, an impaired capacity of BM MSCs to suppress T-cell priming and proliferation has been reported in patients with aplastic anemia, suggesting a possible (primary?) role of BM MSCs in the pathogenesis of the disease [17]. Because an accumulation of activated T-lymphocytes in the BM has been involved in the pathophysiology of CIN, we investigated whether this abnormality could be explained by a defect in the ability of patient MSCs to effectively suppress T-cell activation. However, we found that MSCs from CIN patients sufficiently inhibited mitogen-induced T-cell proliferations in vitro, suggesting that deficient MSC immunosuppressive properties are improbable to account for the aberrant T-cell responses associated with the disease process. The option that BM MSCs do not exert an important role in the immune-mediated pathogenesis of CIN was further corroborated by the normal levels of TNF-α, IL-1β, and IL-6 in patient cultures, a finding indicating that MSCs are unlikely to represent the main cellular source for the aberrant pro-inflammatory cytokine production previously reported in CIN BM [1,4,5].
The levels of TGF-β1, however, were found to be markedly increased in MSC cultures of CIN patients compared with normal controls throughout the culture period. Previous studies have also shown elevated TGF-β1 levels in patient sera and LTBMCs and this cytokine has been critically involved in the pathophysiology of CIN by affecting several parameters of hematopoiesis [4,5,7,32,33]. It has also been proposed that TGF-β1 overproduction in CIN may be genetically determined because CIN patients display at a high frequency the TGF-β1-509C/T polymorphism, which in turn is associated with the elevated cytokine concentration in peripheral blood and LTBMC supernatants. We had no clues until now, however, regarding the cellular population(s) responsible for the overproduction of TGF-β1 in CIN. The data presented herein allow us, for the first time, to assume that BM MSCs may be part of the cellular origin of TGF-β1 in CIN. Further, in accordance with the serum and LTBMC data, patients with the −509C/T polymorphism displayed higher levels of the cytokine in MSC culture supernatants compared with patients with the wild-type genotype and the healthy individuals, suggesting that overproduction of TGF-β1 by BM MSCs in CIN may be also genetically controlled.
Despite the normal morphologic and immunophenotypic characteristics and the normal differentiation and immunosuppressive properties, patient MSCs displayed reduced rate of proliferation time-course as estimated by the population doubling time through passages. This finding was further substantiated by the results of the MTT assay and also by the lower intracellular expression of the Ki-67 nuclear antigen by patient MSCs in representative passages, suggesting lower proportion of cells in active cycle in patient compared with normal cultures [21]. In accordance with the low proliferative potential was the defective clonogenic capacity of patient MSCs compared with normal subjects as shown by the CFU-F clonogenic assay through passages. To probe the underlying cause for these defects, we evaluated the rate of MSC apoptosis in patient and normal cultures time-course. Patient MSCs, however, displayed normal survival characteristics, suggesting that an accelerated apoptotic cell death is unlikely to account for the defective clonogenic and proliferative potential of MSCs in CIN. Also, the elevated TGF-β1 levels in patient cultures did not display an autocrine inhibitory effect on MSC growth as was indicated by the absence of correlation between TGF-β1 levels and CFU-F numbers in all passages tested (data not shown) and by the absence of any improvement in the frequency of CFU-F upon TGF-β1 neutralization. Instead, CFU-F numbers were decreased significantly following TGF-β1 neutralization, suggesting that the cytokine exerts a stimulatory rather than an inhibitory effect on MSC growth. Indeed, it has been shown that although TGF-β1 is a negative regulator of hematopoietic stem cell proliferation [34], it may stimulate the proliferation and expansion of human BM MSCs [35,36]. The significantly reduced clonogenic and proliferative potential of patient MSCs compared with normal controls despite the overproduction of TGF-β1 in patient cultures might be explained by the fact that the stimulatory effect of TGF-β1 on MSC growth has been observed following the exogenous cytokine administration in experimental conditions. Yet, it is unknown how the co-operation between the endogenous TGF-β1 and the complex autocrine cytokine network may act on MSC growth. Nevertheless, the underlying cause for the defective proliferative and clonogenic potential of MSCs in CIN remains unclear. However, consistent with previously reported data from patients with long-standing inflammation within the BM such as rheumatoid arthritis and myelodysplastic syndromes [14,27], we might speculate a premature cell senescence secondary to the long-term inflammatory process present in CIN BM.
In conclusion, we have shown that the reserves, phenotypic characteristics, and differentiation potential of BM MSCs in patients with CIN are normal despite some defects in the cellular growth. Patient MSCs do not seem to contribute to the aberrant T-cell activation associated with CIN as they display normal ability to suppress T-cell proliferative responses and do not display a major role in the pro-inflammatory cytokine production constantly present in CIN BM. We have demonstrated, however, that BM MSCs in CIN may aberrantly produce TGF-β1 due to an intrinsic defect. This is the first study providing evidence that BM MSCs are important cellular elements for the abnormal TGF-β1 production in CIN BM, suggesting that these cells may be primarily involved in the pathophysiology of the disease, at least in patients displaying the -509C/T polymorphism.
Footnotes
Acknowledgment
This work was supported by a grant from the Hellenic General Secretary of Research and Technology (PENED 2003 03ED072).
Author Disclosure Statement
The authors declare no competing financial interests.