
Abstract
Flavonoids have key functions in the regulation of multiple cellular processes; however, their effects have been poorly examined in pluripotent stem cells. Here, we tested the hypothesis that neurogenesis induced by all-trans retinoic acid (RA) is enhanced by agathisflavone (FAB, Caesalpinia pyramidalis Tull). Mouse embryonic stem (mES) cells and induced pluripotent stem (miPS) cells growing as embryoid bodies (EBs) for 4 days were treated with FAB (60 μM) and/or RA (2 μM) for additional 4 days. FAB did not interfere with the EB mitotic rate of mES cells, as evidenced by similar percentages of mitotic figures labeled by phospho-histone H3 in control (3.4%±0.4%) and FAB-treated groups (3.5%±1.1%). Nevertheless, the biflavonoid reduced cell death in both control and RA-treated EBs from mES cells by almost 2-fold compared with untreated EBs. FAB was unable, by itself, to induce neuronal differentiation in EBs after 4 days of treatment. On the other hand, FAB enhanced neuronal differentiation induced by RA in both EBs of mES and miPS. FAB increased the percentage of nestin-labeled cells by 2.7-fold (mES) and 2.4 (miPS) and β-tubulin III–positive cells by 2-fold (mES) and 2.7 (miPS) in comparison to RA-treated EBs only. FAB increased the expression of RA receptors α and β in mES EBs, suggesting that the availability of RA receptors is limiting RA-induced neurogenesis in pluripotent stem cells. This is the first report to describe that naturally occurring biflavonoids regulate apoptosis and neuronal differentiation in pluripotent stem cells.
Introduction
T
Flavonoids are natural polyphenolic compounds present in a variety of vegetables of human intake [3]. These compounds showed several biological properties, including effects on cell differentiation [4,5]. For example, fisetin, luteolin, quercetin, and isorhamnetin induce neural differentiation in pheochromocytoma PC12 cells [6]; icariin and icaritin induce cardiac differentiation in embryonic stem (ES) cells [7,8], whereas biflavonoids such as bilobetin, sciadopitysin, and 7,4′,7′′,4′′′-O-methyl-amentoflavone induce the differentiation of osteoblasts [9].
Flavonoids are also able to reduce cell death [4,5]. Apigenin and kaempferol protect rat cortical neurons against amyloid-β–induced neurotoxicity by inhibiting caspase 2, 3, 8, and 9 [10], whereas the polyphenol epigallocatechin gallate inhibits the fibrillogenesis of both α-synuclein and amyloid-β [11], associated with Alzheimer's and Parkinson's diseases, respectively.
Here, we investigated the biological effects of agathisflavone (FAB; Supplementary Fig. S1E; Supplementary Data are available online at
Materials and Methods
Agathisflavone
Agathisflavone (FAB) (Supplementary Fig. S1E) was extracted from C. pyramidalis Tull leaves as previously described [12], stored at 50 mM in dimethyl sulfoxide (DMSO; Sigma Chemical Co), and kept out of light at 4°C until use.
Cell culture
mES cells, named as USP1, were kindly provided by Dr. Lygia Pereira (University of São Paulo-USP) and Dr. Irina Kerkis (Federal University of São Paulo-Unifesp). mES cells and miPS cells were grown in ES medium as previously described [13,14]. Briefly, cell colonies were grown on mouse embryonic fibroblast (MEF), mitotic inactivated with mitomycin C (Sigma), in 15% knockout serum replacement (KSR; Gibco)–DMEM/F12 (Gibco) supplemented with 1% nonessential amino acid solution (Gibco), 2 mM
For immunocytochemistry, mES or miPS colonies were fixed in 4% paraformaldehyde in phosphate-buffered saline (PBS) for 15 min. The expression of octamer-binding transcription factor 4 (Oct4), a transcriptional factor of pluripotent stem cells, was detected by immunofluorescence assay. Oct4 was uniformly expressed among mES colonies, but not detected on isolated nuclei corresponding to MEF cells (Supplementary Fig. S1B, C). As expected, western blot analysis showed that Oct4 was only detected in mES cells and not on MEFs (Supplementary Fig. S1D).
Retrovirus production
The human complementary DNAs of Oct4, sex-determining region Y-box 2 (SOX2), Krueppel-like factor 4 (Klf4), and cellular-myelocytomatosis viral oncogene homolog (c-Myc) inserted into moloney-based retroviral vectors (pMXs) were obtained from Addgene. Human embryonic kidney 293T cells were utilized as packaging cells to the virus production after calcium phosphate-based transfection with pMXs (20 μg), pMDM (encoding mlv GAG/POL) (10 μg), and pMD.G (encoding VSV G) plasmid (6 μg). The viral supernatants were collected at 48 and 72 h posttransfection, filtered (0.22 μm), concentrated at 32,000 g for 60 min, and incubated in a 2-round transduction on MEF (see below and Supplementary Fig. S2A).
Generation of iPS cells
For the generation of iPS cells, MEFs (Supplementary Fig. S2B) were transduced with concentrated supernatants of 4 retroviruses containing Oct4, SOX2, Klf4, and c-Myc. To improve the transduction, Polybrene (Sigma) was used at 8 μg/mL and the plate was centrifuged at 540 g for 45 min. Two rounds of transduction were performed, with a recovery period of 24 h (d-2 and d-1; see Supplementary Fig. S2A). The medium was changed at day 0, and additional 48 h of recovery time was allowed in 10% fetal bovine serum (FBS; Gibco)–DMEM/F12 (Gibco) supplemented with 2 mM
Herein, our experiments were performed with an miPS clone called i-Rio, which is very similar morphologically to mES cells (Supplementary Fig. S2C). Our miPS cells were positive for the following pluripotency markers: Oct4 (Supplementary Fig. S2D, I), SOX2 (Supplementary Fig. S2F), SSEA-1 (Supplementary Fig. S2G), and phosphatase alkaline (data not shown). As expected, they were able to form embryoid bodies (EBs) (Supplementary Fig. S2J), and as a source to further confirm pluripotenciality, when replated onto gelatin, these EBs were capable to spread out cells from the 3 germ layers: endoderm (alpha fetoprotein; Supplementary Fig. S2K), mesoderm (smooth muscle actin; Supplementary Fig. S2L), and ectoderm (nestin; Supplementary Fig. S2M).
Formation and treatment of EBs
On the first day of culture, day 0, mES and miPS cells (3×106) were passed to a 6-well plate covered with 2% gelatin (Sigma) and cultured for 48 h (d0–d2) in 15% KSR-DMEM/F12 supplemented as above (ES medium) (Supplementary Fig. S1A). At day 2, colonies were treated with TrypLE™ Express (Invitrogen) for 5 min at 37°C, transferred to nonadherent plate dishes with DMEM/F12 medium with 15% FBS (Gibco) and 1% of conditioned medium of CHO cells producing LIF, and cultured for 4 days (from d2 to d6). Spherical cell aggregates (EBs) were formed between d2 and d6 in DMEM/F12 with 15% FBS. EB treatments were performed from d6 to d10 in LIF-free medium and 15% FBS. RA (Sigma) was used to induce neuronal differentiation. At d6, EBs were divided in 4 groups: (a) 0.1% vehicle (DMSO); (b) 2 μM RA; (c) 60 μM FAB; (d) 2 μM RA+60 μM FAB. The medium was changed every other day. At day 10 (d10), EBs were fixed in 4% paraformaldehyde and processed for obtaining 10 μm of cryosections as previously described [16]. In all experiments, 0.1% vehicle (DMSO) group was determined as control.
To allow neuronal migration in some experiments, at d10, EBs were plated onto 1 μg/mL laminin (Invitrogen) and 1 μg/mL fibronectin (Invitrogen)-coated dishes and cultured in basal medium DMEM/F12 with N2 supplement and 20 ng/mL of fibroblast growth factor-2 for additional 4 days [13].
Immunofluorescence analyses
Prior immunofluorescence assay slides with EB frozen sections were heated in a boiling bath of citrate buffer (0.01 M, pH 6.0) for 10 min. Colonies were treated with 0.3% of Triton X-100/PBS (Isofar), whereas EBs were treated with 0.5% Triton X-100 in 50 mM NH4Cl/PBS for 5 min. After incubation with 5% bovine serum albumin (Invitrogen) for 30 min, samples were incubated with the primary antibody overnight. After washing, cells were incubated for at least 1 hour with the secondary antibodies: goat anti-mouse IgG conjugated to Alexa Fluor 546 (Molecular Probe; 1:400) or goat anti-rabbit IgG conjugated to Alexa Fluor 488 (Molecular Probe; 1:400). The samples were washed with PBS and nuclear staining was performed with 1.0 μg/mL solution of 4′,6-diamidino-2-phenylindole (DAPI) [13]. Images of cells were accessed with optic fluorescence microscopy (Nikon T300).
Antibodies used to assess pluripotency were anti-OCT 3/4 (Santa Cruz Biotechnology; mouse, 1:100), anti-SOX2 (Chemicon; rabbit, 1:100), and anti-SSEA1 (Chemicon; mouse, 1:100). The 3 germ layers were detected with endoderm–anti-alpha fetoprotein (Santa Cruz Biotechnology; rabbit, 1:100), mesoderm–anti-smooth muscle actin (Chemicon; mouse, 1:100), and ectoderm–anti-nestin (Chemicon; mouse, 1:100). To assess cellular mitosis rate, histological slices of EBs were incubated with polyclonal antibody anti-phospho-histone H3 (Upstate; rabbit, 1:400). Antibody anti-nestin (Chemicon; mouse, 1:100) and anti-β-tubulin III (Covance and Sigma; mouse, 1:100) were used to label, respectively, neural progenitor cells and neuronal cells in EB slices. Antibodies used to verify the expression of both RA receptors were anti-RARα and anti-RARβ (Santa Cruz Biotechnology; rabbit, 1:100).
For analysis of apoptosis in EBs, DNA fragmentation was detected by the terminal-deoxynucleotidyl-transferase dUTP nick-end labeling (TUNEL; Promega) on frozen EB sections, performed according to the manufacturer's protocol [13,17]. Cell death was also identified by cleaved caspase-3 immunofluorescence analysis using the antibody anti–cleaved caspase-3 (Cell Signalling; rabbit, 1:100).
Alkaline phosphatase staining
Alkaline phosphatase (a stem cell membrane marker associated with undifferentiated pluripotent stem cell) staining was performed in miPS colonies with the Alkaline Phosphatase Detection Kit (Millipore) to further detect pluripotency of these cells.
Quantitative analysis
The percentages of positive cells labeled with nestin, β-tubulin III, TUNEL, phopho-histone H3, or cleaved caspase-3 were calculated as follows: at each EB section, positive cells were counted, as well as the total number of cells stained with DAPI. For quantification, photomicrographs taken under 400× microscope magnification were 150% amplified on Adobe Photoshop for visualization of individual cells within the EB section. A multiplatform software [Embryonic Stem Cell Counter (ESCC)] was used for counting DAPI-positive cells from photomicrographs of EB sections. The software can be freely downloaded (
Semiquantitative analysis of the immunofluorescence of RAR alpha and beta were carried out as follows: photomicrographs were taken with the same fluorescence intensity under confocal microscope (Leica TCS SP5 AOBS). Once converted to grayscale, Image J 1.4.3U software was used to measure the area and to calculate the fluorescence intensity of each EB. The fluorescence intensity of EBs was subtracted from the background. Graphics represent the rate of fluorescence intensity per EB area. We counted about 20 EBs per group and each graphic expresses the mean of the average rates of at least 3 independent experiments. Statistical analysis was performed using unpaired Student's t-test.
Results
Agathisflavone enhances neuronal differentiation induced by RA in EBs
To test the effects of FAB on ES neuronal differentiation, EBs were treated with RA (2 μM), FAB (60 μM), or both together (RA 2 μM and FAB 60 μM) (Supplementary Fig. S1A, F). On day 10, the percentage of neural progenitors within the aggregates was determined (Fig. 1A, B). No differences in the number of nestin-positive cells were observed between control (2.1%±0.7%) and FAB-treated cells (0.4%±0.3%); however, when FAB was combined with RA (RA+FAB group), the percentage of neural progenitor cells increased by 2.7 times (9.6%±0.6% in RA alone and 26%±6.5% in the RA+FAB) (Fig. 1A, B).
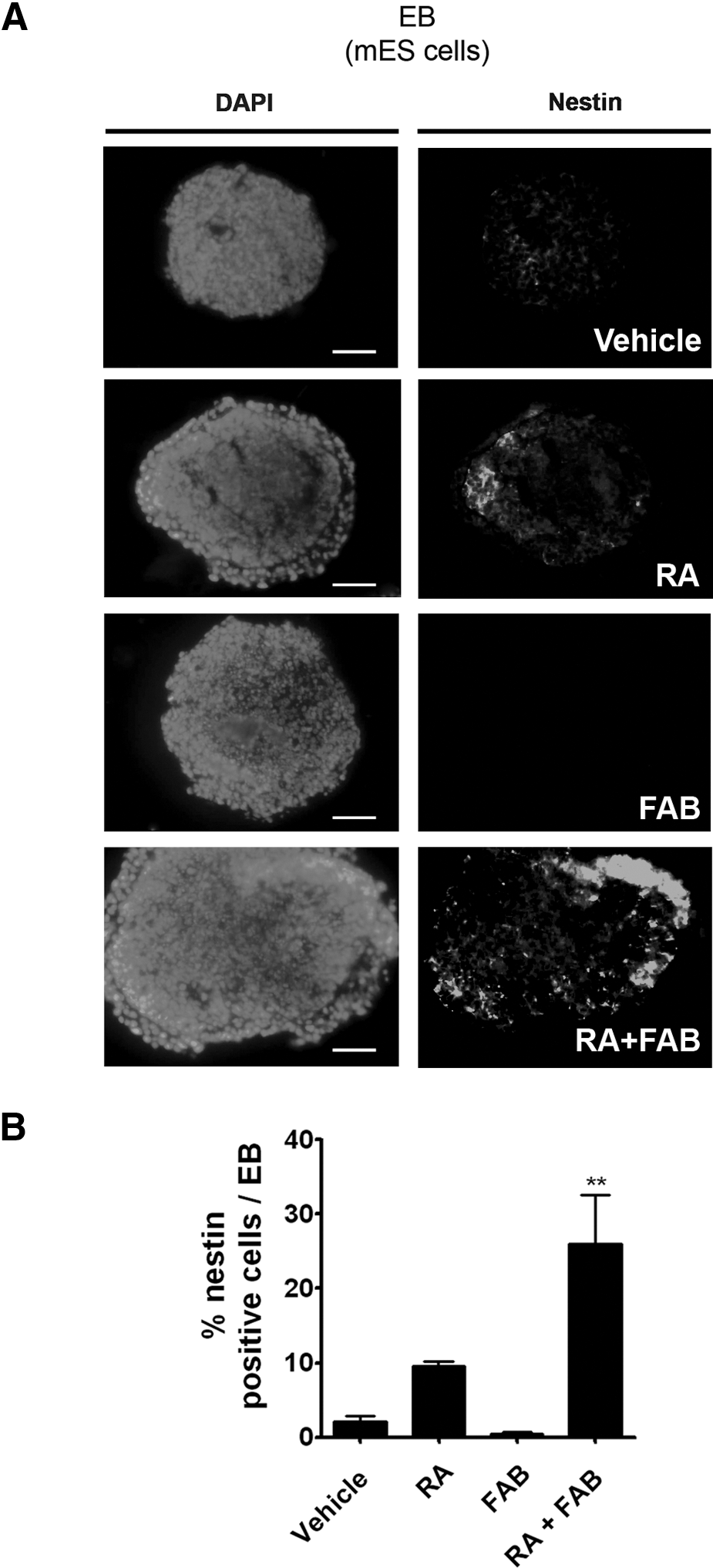
Agathisflavone increases the number of neural progenitor cells induced by RA in mES cells.
To confirm the number of young neurons, we assessed the percentage of β-tubulin III–positive cells, which were generated after RA or RA+FAB treatment. RA increased the number of β-tubulin III–positive cells (12%±2.1%; Fig. 2A, B) in comparison to controls (2.2%±0.3%; Fig. 2A, B), whereas FAB alone did not increase the number of β-tubulin III–positive cells (1.8%±0.9%). However, FAB together with RA (RA+FAB) doubled the percentage of β-tubulin III–positive cells (24.2%±4.3%) in relation to RA alone (12%±2.1%; Fig. 2A, B). To better compare the amount of neuronal cells between the treatments, EBs were plated on laminin/fibronectin-coated plates and cultured for additional 4 days in neurobasal medium free of RA or FAB. A subset of migrating β-tubulin III–positive cells with long processes emerged from EBs (Fig. 2C). Yet, the number of β-tubulin III–labeled cells migrating out of EBs after treatment with the combination of RA+FAB was visibly higher than in EBs treated with RA only (Fig. 2C). These results suggest that FAB enhances RA-induced neural differentiation in ES cells.
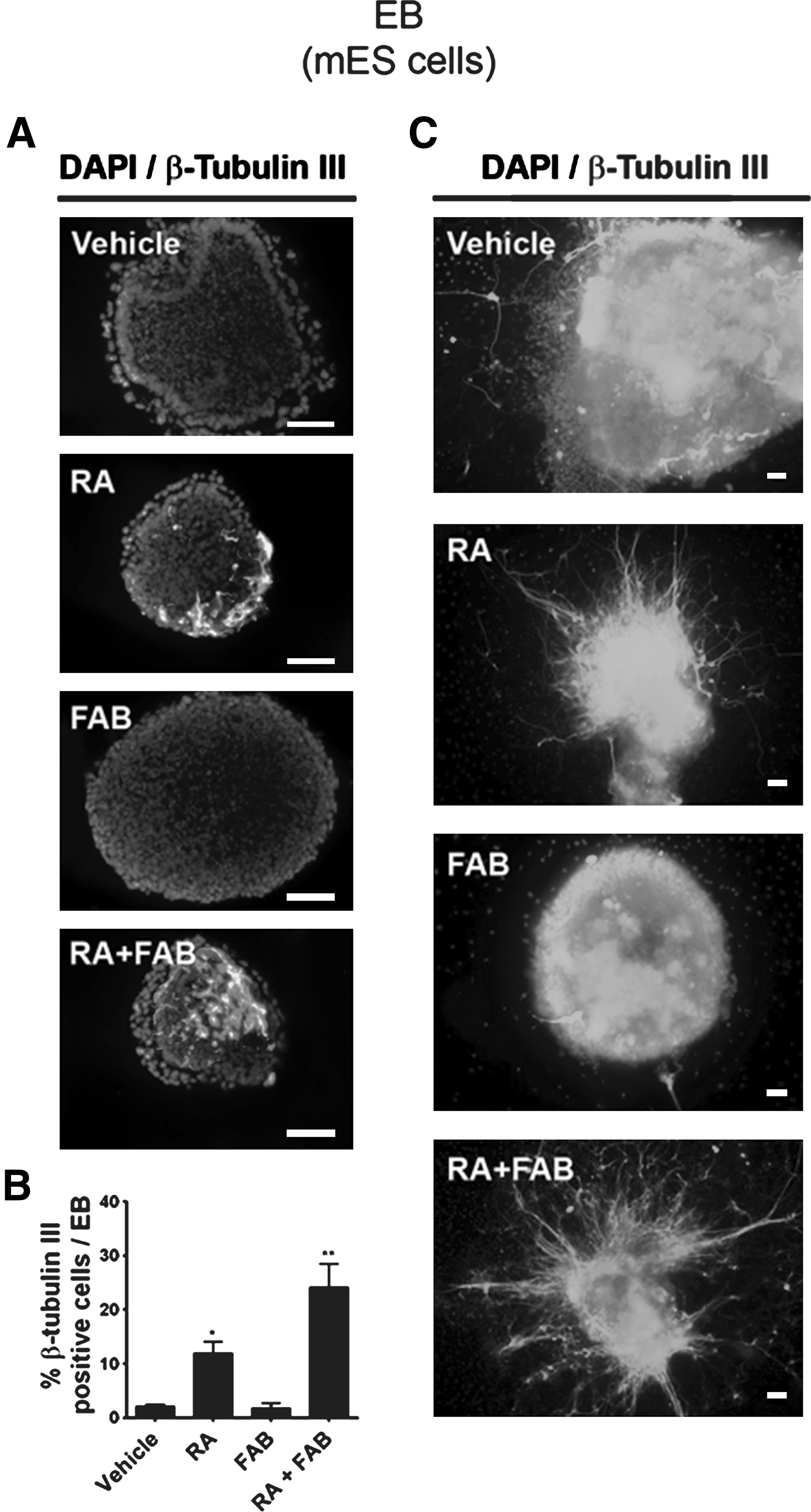
Agathisflavone increases the number of young neurons induced by RA in mES cells.
Agathisflavone also enhances neural differentiation triggered by RA in iPS cells
Colonies of miPS (Supplementary Fig. S2C) were generated from MEF cells by reprogramming. Pluripotency was confirmed by immunofluorescence for specific markers of undifferentiated ES cells (OCT-4, SOX2, and SSEA1; Supplementary Fig. S2D–I) and also their ability to originate cells from ectoderm, mesoderm, and endoderm (Supplementary Fig. S2J–M). Administration of FAB on miPS EBs did not increase the percentage of cells labeled for nestin (4.9%±3.0%) or β-tubulin III (0.8%±0.4%) in comparison to the percentage of nestin (1.9%±0.2%) or β-tubulin III (5.2%±2.5%)–labeled cells in control EBs (Figs. 3 and 4). As we observed with mES EBs, FAB together with RA increased the percentage of nestin-labeled cells by 2.4 times (46.3%±1.5% RA+FAB) in comparison to the percentage nestin-positive cells in RA only (19.3%±4.8%) (Fig. 3). The amount of β-tubulin III–positive cells was also increased in 2.7-fold by FAB+RA (36.9%±11.3%) when compared to RA treatment alone (13.9%±3.1%) (Fig. 4). In conclusion, FAB increases the amount of neural progenitor cells induced by RA in both mES and miPS EBs.
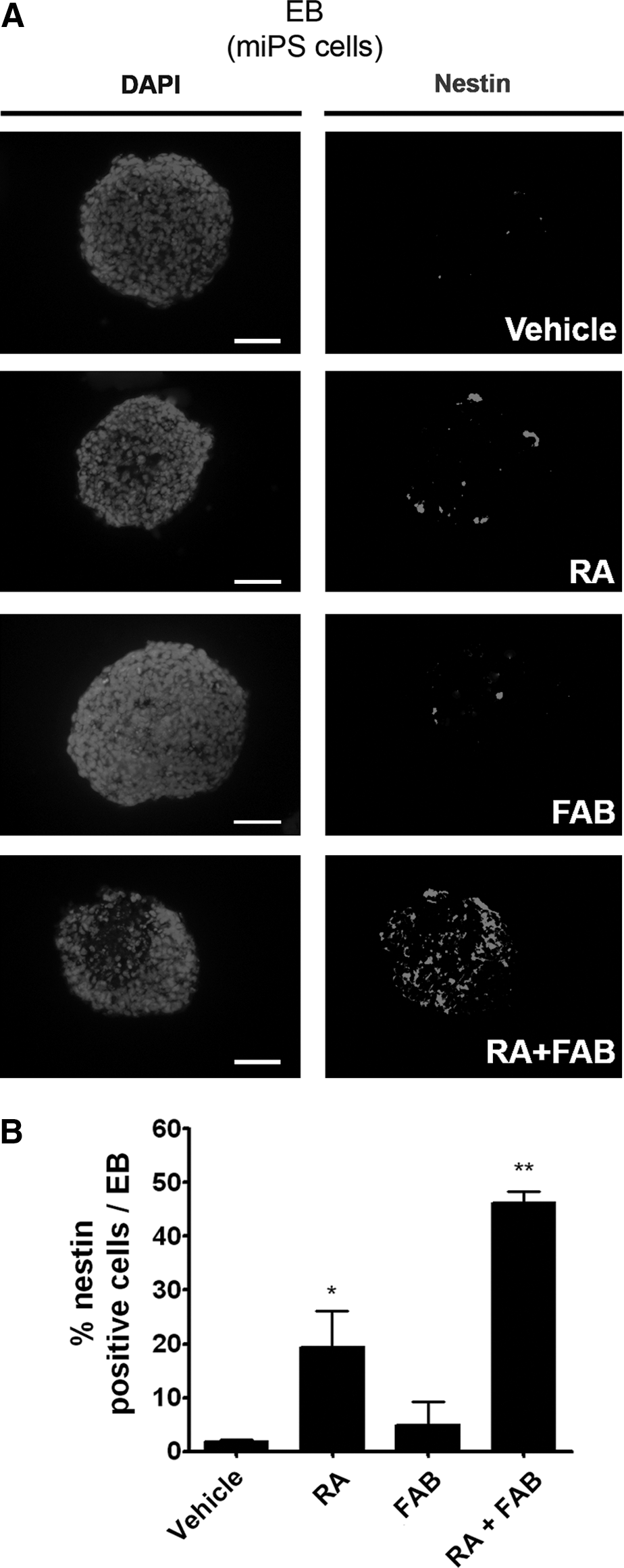
Agathisflavone increases the number of neural progenitor cells induced by RA in miPS cells.
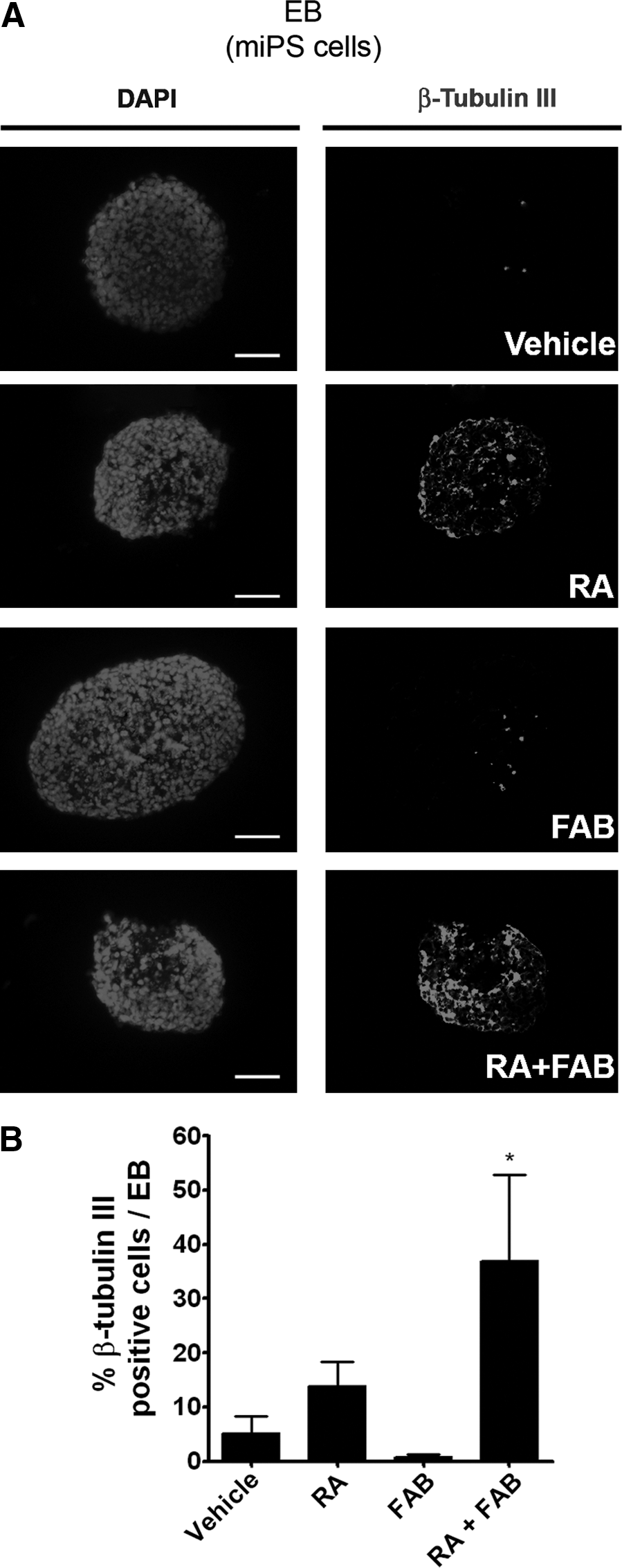
Agathisflavone increases the number of young neurons induced by RA in mouse iPS cells.
Agathisflavone does not affect the mitotic rate, but decreases cell death in EBs
To investigate whether the number of neuronal progenitors was increased by an effect of FAB on proliferation, we evaluated the effects of FAB in mitosis of pluripotent stem cells growing as EBs. Frozen sections of EBs were processed for immunofluorescence to identify phosphorylated histone H3 (Fig. 5A), a G2/mitosis specific marker [18,19]. The amount of positive cells for phosphorylated histone H3 was similar in the experimental groups (Fig. 5B), suggesting that FAB has no effect on the mitotic rate of both mES and miPS cells.
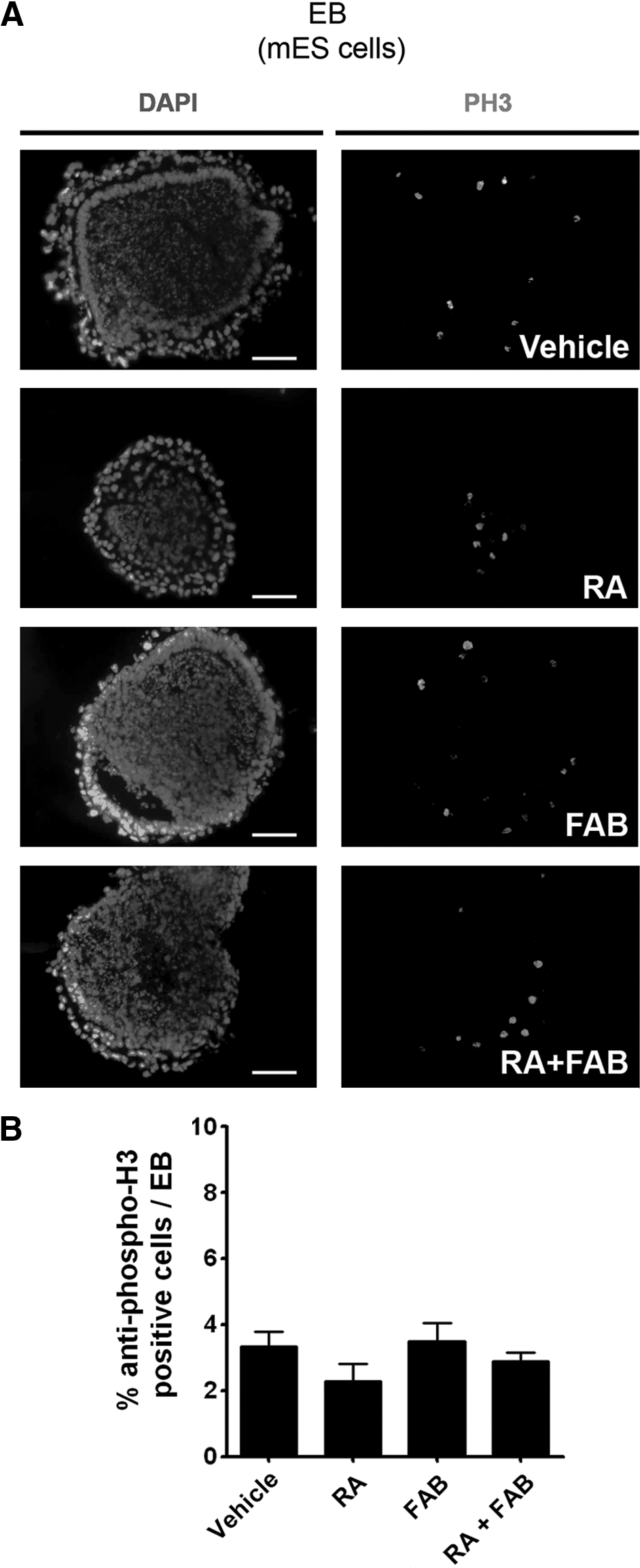
Agathisflavone does not alter mitosis in EBs.
As some flavonoids, such as casticin, are able to increase the survival of neuronal progenitor cells in vitro [20], we investigated whether FAB could reduce cell death in EBs undergoing neurogenesis. FAB was able to decrease the amount of TUNEL-positive cells in all experimental groups (Fig. 6A, B). Similar antiapoptotic effect of FAB was also observed by immunofluorescence analyses of cleaved caspase-3 within EBs (Supplementary Fig. S3).
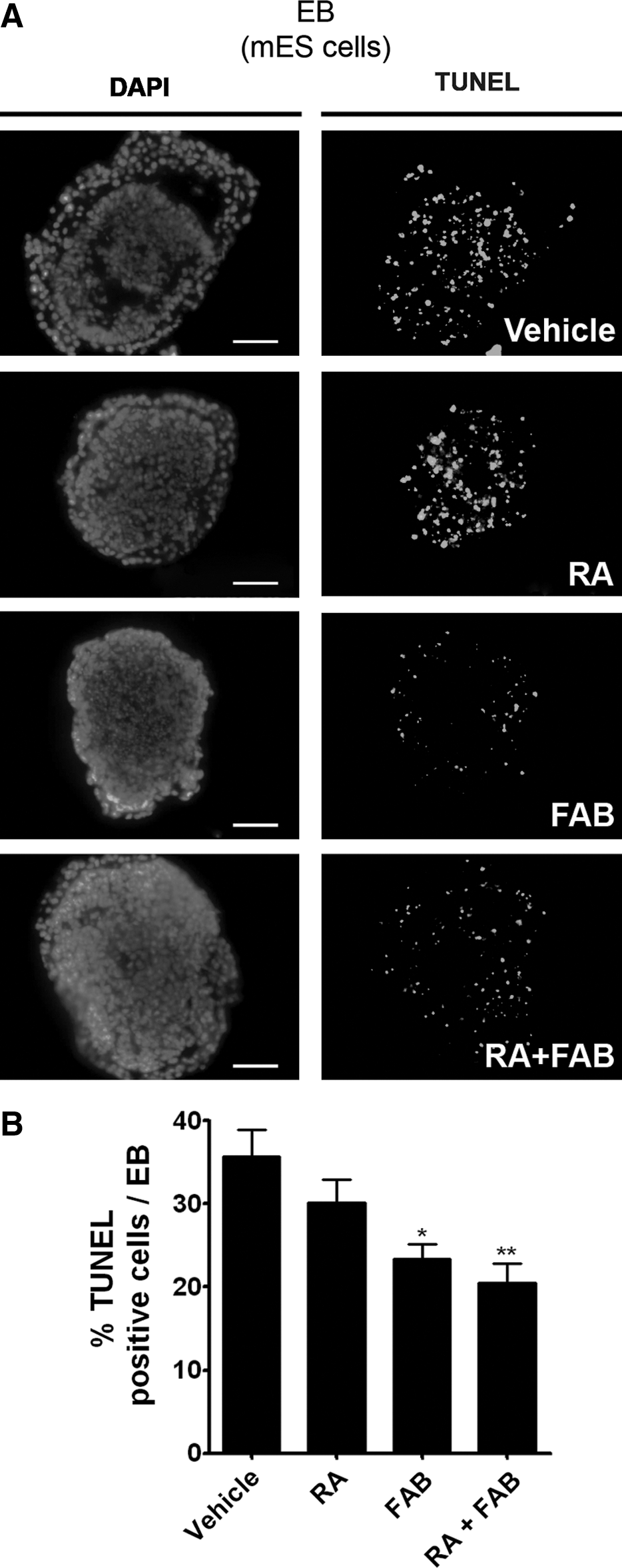
Agathisflavone increases cell survival in EBs.
To evaluate whether FAB was protecting specifically neuronal progenitor cells, we analyzed EBs with 2-color fluorescent staining for TUNEL and neuronal markers (nestin and β-tubulin III) by confocal microscope (Fig. 7A, B). The percentage of nestin– and β-tubulin III–positive cells undergoing cell death was statistically higher in RA-treated cells (nestin: 50.4%±3.1%; β-tubulin III: 47.1%±1.6%) than within the RA+FAB treated group (both below 1%) (Fig. 7C, D). These results suggested that the effect of enhancing RA-induced neurogenesis is, at least in part, due to FAB promoting the specific survival of neuroprogenitor cells derived from pluripotent stem cells.
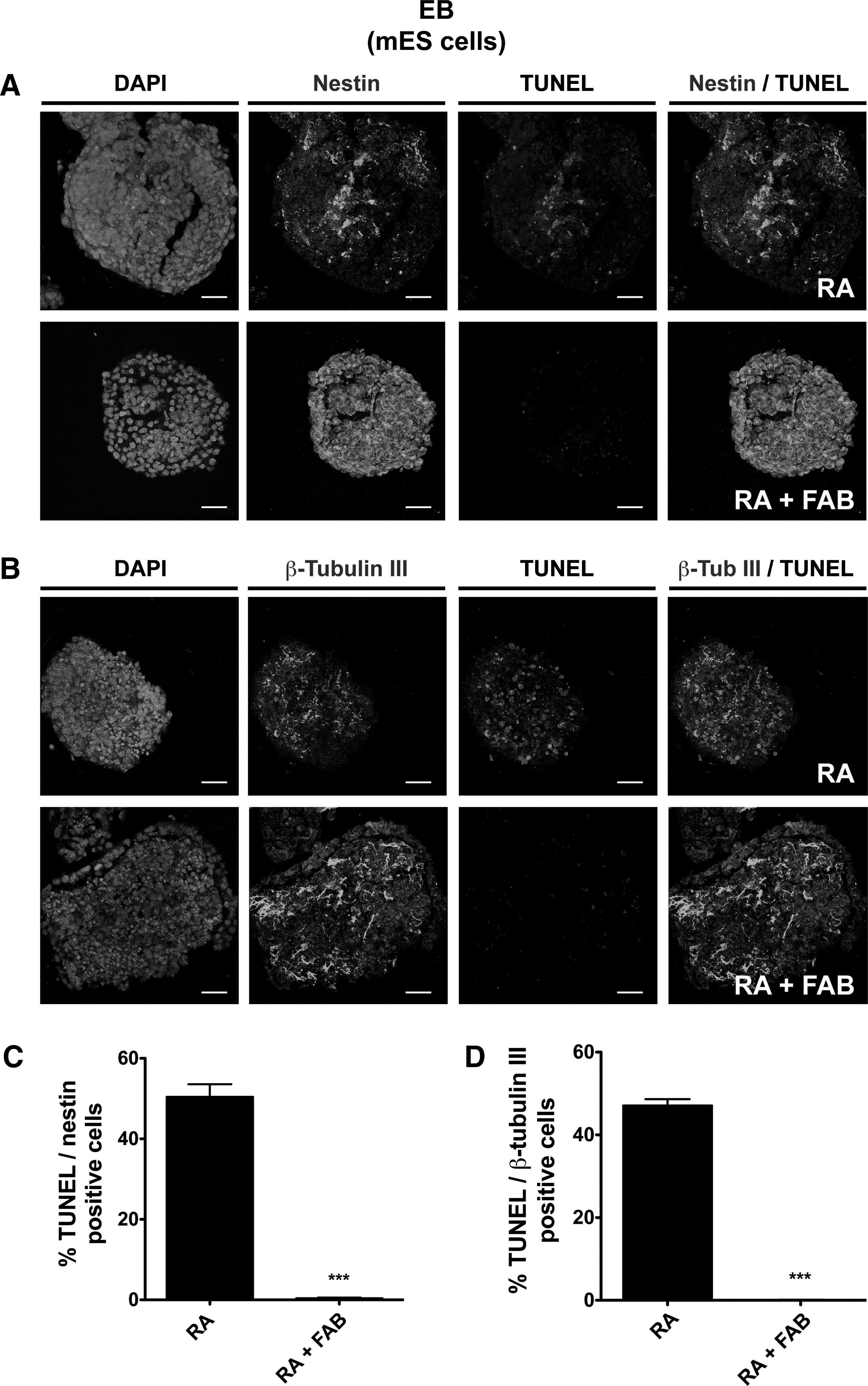
Agathisflavone promotes the survival of neuroprogenitor cells in EBs. EBs derived from mES cells were treated with 2 μM RA or the combination of 2 μM RA plus 60 μM FAB (RA+FAB).
Agathisflavone increases the expression of RAR in EBs treated with RA
It is already known that RARs, such as RARβ, are important to neuronal differentiation of mES cells [21] and that sequential activation of both RARβ and RARα are able to induce neuronal differentiation in cultured neural progenitor cells [22]. In order to characterize a possible mechanism by which FAB enhances neurogenesis induced by RA in pluripotent stem cells, we examined whether FAB increase the expression of RAR in mES after RA treatment. Immunofluorescence assays for RARα and RARβ in EBs treated with RA alone or with of RA and FAB (Fig. 8A, C) reveal that FAB increases about 3.7 times the expression of both RARα and RARβ (Fig. 8B, D). These results suggest that FAB enhances neural differentiation induced by RA in mouse pluripotent stem cells by increasing the expression of RAR α and β.
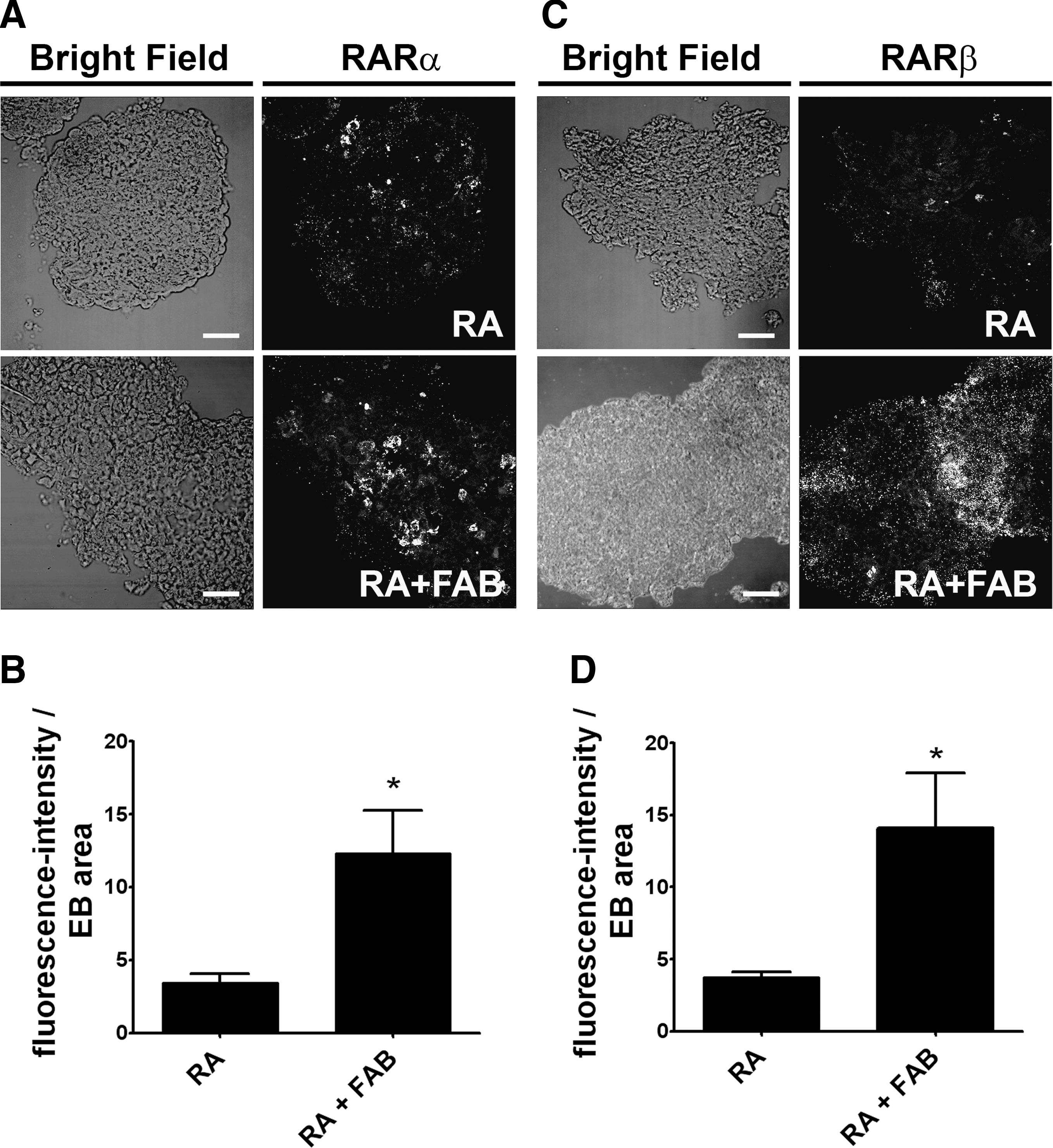
Agathisflavone increases the expression of RARs α and β in EBs treated with RA.
To investigate whether upregulation of RARs were relevant to induce RA-neurogenesis, EBs of mES cells were preconditioned with FAB during EB formation (from d4 to d6; Supplementary Fig. S4). After washing-out, RA incubation was carried out for 4 days (d6 to d10). The expectation was that the preconditioning with FAB would increase the expression of RAR and even after the FAB was washed-out, it would provide an increase on neuronal differentiation induced by RA. The percentage of β-tubulin III-positive cells detected in the group preconditioned with FAB (FAB/RA) was significantly higher (24.0%±1.0%) than RA treated group (12.1%±0.5%) or vehicle (0.3%±0.1%) (Fig. 9), suggesting that preconditioning with FAB increases neuronal differentiation by upregulating RARs even before RA treatment.

Pretreatment with agathisflavone increases the number of young neurons differentiated after RA treatment in EBs.
Discussion
The main feature of pluripotent stem cells is their ability to differentiate into any other cell type. Nevertheless, to design efficient protocols for amplifying differentiated cell populations is still a challenge that demands new strategies and the combination of multiple compounds. In this sense, small molecules have been intensely investigated as inducers of specific differentiation in pluripotent stem cells [23].
Flavonoids are natural polyphenolic compounds present in a variety of vegetables and Chinese herbal medicines [3], which show biological activities including antioxidative [24], anti-inflammatory [25], and antitumoral effects [26 –28]. The variety and relatively low toxicity of flavonoids compared with many active plant compounds highlights their potential for therapeutic use. As an example, the flavonoid epicatechin was able to decrease motor neuron specific death in a drug-screening model for amyotrophic lateral sclerosis by reducing reactive oxygen species (ROS) production in mutated astrocytes [29]. Further, polyphenol epigallocatechin gallate and tocotrienol were included as candidates for the treatment of familial dysautonomia disease using familial dysautonomia patient-specific iPS cells [30].
The influence of FAB on proliferation, cell death, and neural differentiation has never been explored in mES and miPS cells. This biflavonoid did not change proliferation or neural differentiation rates in those cells, but it was able to reduce cell death, which was about 35% in 0.1% of DMSO (vehicle) group. The reduction of cell death by FAB is consistent with previous results that show that flavonoids can reduce cell death [10,20]. The exact mechanism by which flavonoids protect against cell death may be vast, as those compounds are widely known not only as antioxidant, but also as activators of PI3 kinase, tyrosine kinase and MAP kinase pathway [4,31,32], enhancers of proteasome activity [33], and modulators of protein assembly and oligomerization [11]. Since DMSO is the solvent most suitable for maintaining the stability of RA and FAB [5,6,9,20,23,28], side effects induced by the vehicle might be an inevitable feature of the protocol. In this context, the association of FAB to the protocol becomes applicable in order to reduce inevitably cell death induced by the DMSO and RA. Nevertheless, the ability to reduce cell death per se opens an opportunity to use flavonoids to increase the survival of differentiated stem cells in vitro. As our data suggest that FAB is promoting the survival of neuroprogenitor cells derived from pluripotent stem cells.
Agathisflavone enhanced the effectiveness of RA as a neural morphogen in both mES and miPS cells, whereas the percentage of neural progenitor cells and young neurons increased more than 12-folds. These observations were associated with an increase in the expression of RARα and RARβ in ES cells and even the preconditioning of EBs with FAB was able to increase neuronal differentiation once it is presumably upregulating RAR before RA treatment.
Both receptors have shown to be relevant to neurogenesis, since: (i) they are expressed in the subventricular zone of the adult mice [34]; (ii) the sequential activation of RARβ and RARα induces neuronal differentiation in cultured neural progenitor cells [22]; (iii) RARβ are necessary for RA-induced differentiation, as RARβ mutant in mES cells fail to differentiate, while remained pluripotent [21].
In the present study, we show that agathisflavone (FAB) increases the expression of RARα and RARβ in ES cells, which probably improves the ability of RA to induce neural differentiation in pluripotent stem cells. It is worth mentioning that flavonoids, such as apigenin, are able to activate estrogen receptors [35], which promote amplification of the RA signaling pathway [36] by increasing the expression of their receptors. As FAB is an association of 2 apigenins, it is possible that FAB effects as an enhancer of RA-induced neurogenesis is associated to an increase of the amount of RARs in pluripotent stem cells.
Recently, the flavonoid icaritin had been reported to induce neuronal differentiation in mES cells growing as EBs [23]. The authors described that icaritin was able to induce neuronal differentiation by itself when EBs were cultured for at least 10 days in neurobasal medium supplemented with B27. The expression of nestin and β-tubulin III was similar in the icaritin-treated cells and RA alone, suggesting that this flavonoid can act as a substitute of RA rather than an enhancer of RA neurogenic effects.
Taken together, our results indicate that FAB increases cell survival and RA-induced neuronal differentiation in pluripotent stem cells. To our knowledge, this is the first report to describe that naturally occurring biflavonoids regulate apoptosis and neuronal differentiation in both mES and miPS cells. These results may benefit the development of novel protocols to enrich the number of differentiated neurons derived from pluripotent stem cells in vitro.
Footnotes
Acknowledgments
This work was supported by Instituto Nacional de Ciência e Tecnologia em Terapia Celular (INCTC); Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ); Conselho Nacional para o Desenvolvimento Científico e Tecnológico (CNPq), Instituto do Milênio de Terapia Gênica, Ministério da Saúde/DECIT, Fundação do Câncer; and Instituto Nacional do Câncer (INCa). The authors thank Ismael Gomes and Severino Galdino for technical assistance.
Author Disclosure Statement
The authors report no conflicts of interest, including personal or financial.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.