
Abstract
In this study we have shown that the papilla of the mouse kidney contains a population of Pax2+ cells that are detectable from the early postnatal period through to adulthood. Lineage analysis suggests that some of these Pax2+ cells are derived from the metanephric mesenchyme, a population of progenitor cells that gives rise to the nephrons during kidney organogenesis. Here we describe a method for isolating and culturing the Pax2+ population, and demonstrate that some cells within this population are multipotent stem cells, as they are clonogenic and appear to undergo unlimited self-renewal. Further, under appropriate culture conditions, these stem cells can differentiate to generate renal cell types, such as podocyte- and proximal tubule-like cells, and are also able to generate nonrenal cell types, such as adipocytes and osteocytes. The availability of a kidney-derived multipotent stem cell line with the potential to generate podocytes and proximal tubule cells in culture will expedite progress in understanding the biology of these important renal cell types, and will be a useful tool in toxicological studies and drug discovery.
Introduction
I
A characteristic of most kidney stem cell populations is that they are able to differentiate in vitro to generate various renal and nonrenal cell types [1,2,4,5,6,8,9]. It is worth noting, however, that as yet, rodent kidney stem cell populations in vitro appear unable to generate proximal tubule cells (PTCs), or cells that have the typical morphological characteristics of podocytes. This is an important goal because podocyte and PTC dysfunction is known to be a cause of renal disease [13,14]. It is now well recognized that in vitro culture presents a valuable model system with which to study the function, physiology, and pathophysiology of podocytes [15] and PTCs [16]. However, progress in this area is currently restricted by the inherent difficulties in culturing and expanding these cell types in vitro. The development of conditionally immortalized podocyte cell lines has been a step forward in facilitating the study of podocyte biology [17,18]. However, the use of such genetically modified cells can be problematic because their behavior usually differs from endogenous nontransformed cells. Likewise, although much use has been made of established PTC lines, such as the opossum kidney-derived cell line, OK, and the human tubular cell line, HK2, there is a tendency for such lines to de-differentiate and lose their specific biochemical properties [16].
The ability to generate podocytes and PTCs in vitro from kidney-derived stem cells would present a distinct advantage, for, in addition to avoiding the use of immortalized podocyte cell lines and atypical PTC lines, it would enable the mechanisms that regulate the differentiation of these important cell types to be investigated. Further, such cells could have useful applications in drug screening and regenerative medicine.
Although a number of reports have indicated that kidney stem cells can give rise to podocytes in vitro, this is usually based on the expression of 1 or 2 podocyte markers [5,6], and morphologically, these cells bear little resemblance to primary podocytes. It has recently been proposed that to define a cell in vitro as a true podocyte, apart from the expression of podocyte-specific markers, the cells should also display typical morphological characteristics, which include a high cytoplasmic-to-nuclear ratio, arborized appearance, prominent cell processes, and a high incidence of binucleation [15]. Similarly, evidence that kidney stem cells can generate PTCs in vitro is not very convincing [6] because such cells do not display the typical epithelial morphology of primary PTCs [16], nor has it been demonstrated that they express functional PTC-specific enzymes, such as alkaline phosphatase [19].
The aim of this study was to establish the origin of stem cells present in the mouse postnatal kidney, and to investigate their potential for generating podocyte and PTCs in vitro. Further, we investigated if these kidney-derived stem cells were multipotent, by assessing their ability to generate nonrenal cell types such as adipocytes and osteocytes.
Materials and Methods
Isolation and culture of kidney-derived stem cells
Kidneys were dissected from 2–6-day-old CD-1 mice (Charles River), cut into <1 mm pieces, and incubated in 1 mg/mL collagenase type I (Sigma) and 0.1 mg/mL DNase I (Sigma) in Hank's buffered saline with calcium and magnesium (Invitrogen). The cell suspension was passed twice through 21- and then 23-gauge needles, and finally through a 30 μm preseparation filter. Cells were seeded on the following substrates: (i) SIM(sandoz inbred Swiss mouse) thioguanine-resistant ouabain resistant (STO) feeder cells; (ii) STO feeder cells treated with sodium deoxycholate; (iii) fibronectin (FN). For all substrates, the culture medium comprised a 1:1 ratio of Advanced ® DMEM (Invitrogen):STO-conditioned medium, supplemented with 2 mM glutamine (Invitrogen) and 2.5% FCS (PAA laboratories). For sodium deoxycholate treatment, STO cells were incubated in 0.5% (w/v) sodium deoxycholate and 1 mM phenylmethyl sulfonylfluoride in 10 mM Tris-HCl pH 8.0 for 30 min at 4°C. FN-coated dishes were prepared by adding 1 mL of a 50 μg/mL solution of human plasma FN (Millipore) in phosphate-buffered saline (PBS) to 3.5 cm tissue culture dishes for 1 h at 37°C. The FN solution was aspirated immediately before plating the cells. To promote renal cell differentiation, cells were cultured on tissue culture plastic in high glucose DMEM (Invitrogen) supplemented with 2 mM glutamine and 10% FCS. Adipogenic and osteogenic assays were performed as previously described [5]. To generate clonal lines, the bulk population was trypsinized to give single cells, which were then transferred to 96-well plates by limited dilution. Initial attempts to generate clonal lines comprised culturing the cells on an FN substrate in STO-conditioned medium in the presence of 2.5% serum. However, under these conditions, single cells failed to survive. Therefore, to improve clonal growth, the serum content of the medium was increased to 10%. Three to 4 h after seeding, the plates were screened using an inverted microscope, and any wells containing 0 or >1 cells were excluded from further analyses. Images of single cells were taken at day 0 (day of plating), day 2, day 3, and then every 1–2 days until day 12. Single cells that were able to generate lines were expanded and characterized using immunostaining and reverse transcriptase-polymerase chain reaction (RT-PCR) between passages 8 and 10.
Immunofluorescence and lectin staining
Cytofluorimetric analysis, immunofluorescence, and preparation of frozen sections were performed as previously described [2,20]. Fixed cells or frozen kidney sections from wild-type and Wt1-Cre/Rosa26R mice were incubated overnight at 4°C with primary antibodies to Pax2 (Covance), Wt1 (Millipore), Sox2 (Cell Signaling), β-galactosidase (Abcam), pan-cytokeratin and αSMA (Sigma), Oct4, aquaporin1 and aquaporin2 (Santa Cruz), synaptopodin (Fitzgerald), and musashi1 (Chemicon Int.). Secondary antibodies were anti-rabbit-488 and −594, anti-mouse IgG2a-594, anti-mouse IgG1-488, anti-chicken IgG-488, and anti-goat IgG-594 (Invitrogen). For lectin and Pax2 co-staining, the sections were washed 3 times in Tris buffered saline (TBS), and incubated with PNA lectin (Vector Labs) at a concentration of 10 μg/mL (v/v) in TBS for 2 h at room temperature. Sections were then washed 3 times in TBS and blocked with 0.1% (v/v) Triton X (Sigma) and 10% (v/v) chicken serum (Sigma) in PBS for 1 h at room temperature, and immunostained for Pax2. To determine the percentage of Pax2+ cells in the papilla of 7-day-old Wt1-Cre mice that expressed β-galactosidase, the total number of Pax2+ cells within 4 different fields of view was counted, and the number co-expressing β-galactosidase was determined. Samples were analyzed using a LEICA AOBS SP2 confocal microscope or a Leica DM2500 fluorescence microscope, and images were captured using a DFC350FX camera. Antibodies for cytofluorimetric analysis were all phycoerythrin or fluorescein isothiocyanate-conjugated and were obtained from Dakoycytomation. All incubations were performed in 100 μL of PBS containing 0.1% bovine serum albumin and 0.1% sodium azide at 4°C. For each sample, 10,000 cells were analyzed on FACSCalibur cytometer (BD Biosciences Pharmingen). Gating was constructed based on negative controls and compensation controls were included in all analyses performed. Population percentages and numbers were generated for gated populations from each experiment using Cell Quest software (BD Biosciences Pharmingen).
Alkaline phosphatase activity
Alkaline phosphatase staining solution was prepared by adding 2 mg of naphthol AS-MX phosphate (Sigma) and 10 mg of Fast Red TR (Sigma) to 10 mL Tris-HCl pH 9.2. Samples were analyzed using a Leica DM1000 microscope and images were acquired using a DFC420C digital camera (Leica).
Reverse transcriptase-polymerase chain reaction
RT-PCR was performed as previously described [21]. Glyceraldehyde 3-phosphate dehydrogenase was used as a reference gene. For primer sequences see Table 1. The specificity of primers designed in-house was confirmed by sequencing of PCR products (DNA Sequencing Laboratory, Dundee, United Kingdom).
Results
A population of Pax2+ putative kidney stem cells is present in the postnatal mouse kidney
Kidney stem cells in postnatal human and rat kidney express the MM-specific marker, Pax2 [2,4]. To investigate the prevalence and location of Pax2+ cells in postnatal mouse kidney, immunostaining was performed on kidney sections at various time points from postnatal day 1 (P1) through to adulthood. To show which structures the Pax2+ cells were localized to, co-staining was performed with PNA lectin, which binds to the proximal tubules and collecting ducts in the mouse renal cortex [22]. There are no published observations of PNA staining in the mouse renal papilla, but in rabbit, PNA stains the thin descending loops of Henle and the collecting ducts [23]. Up to the seventh postnatal day (P7), many Pax2+ cells were present in the nascent glomeruli, proximal tubules, and collecting ducts in the outer cortex, but as expected, were no longer present in this region after cessation of nephrogenesis at P14 (Fig. 1A). Pax2+ cells were also present in the papilla of neonatal mice, but unlike those in the cortex, were detected postnephrogenesis at all life stages tested, from P1 (Fig. 1A) through to adulthood (Supplementary Fig. S1A; Supplementary Data are available online at
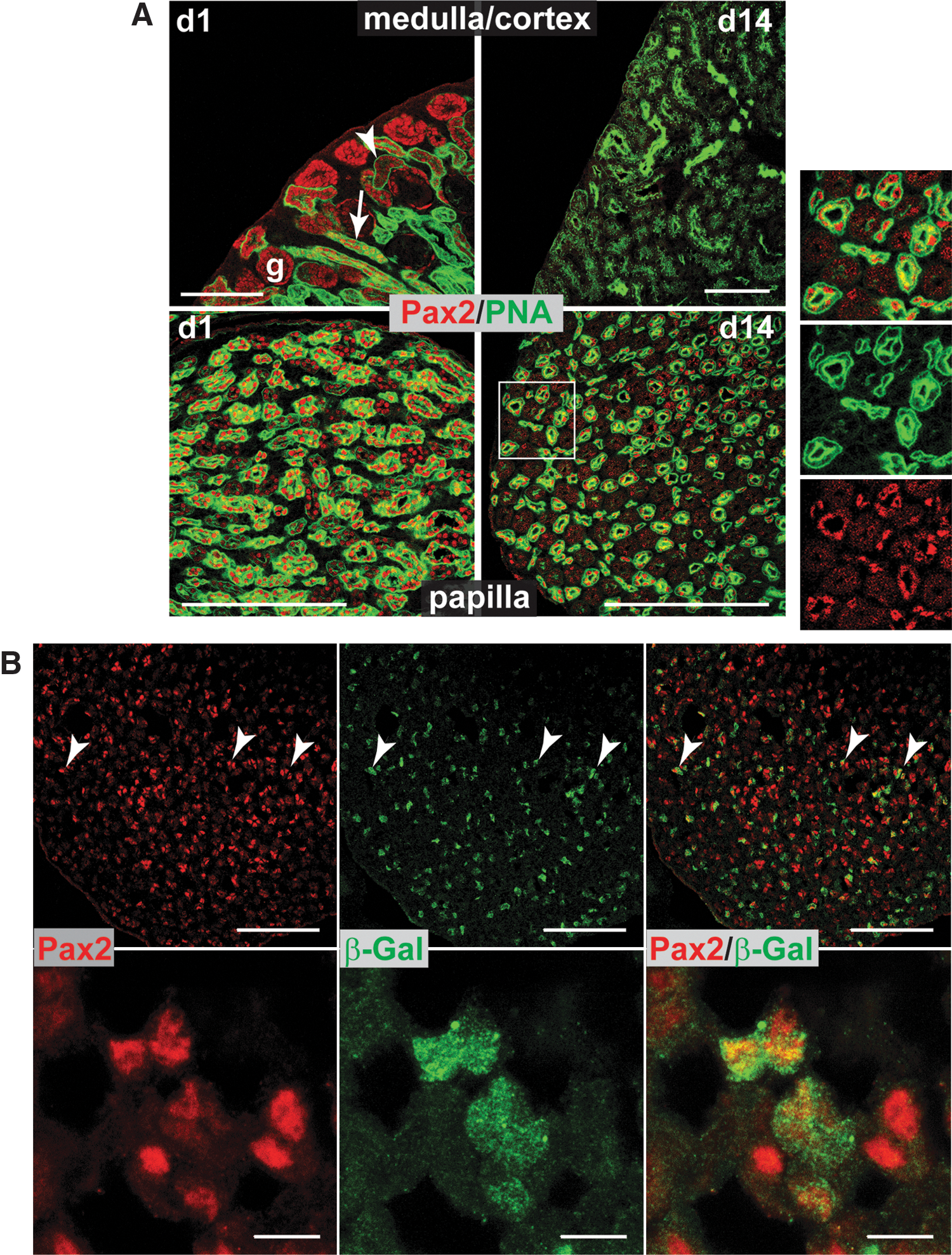
The postnatal mouse papilla contains Pax2+ cells that are derived from the metanephric mesenchyme.
During embryogenesis, kidney nephrons are derived from Wt1+/Pax2+ MM cells, whereas the collecting tubules and ureters are derived from Wt1−/Pax2+ UB cells. To confirm the origin of the papillary Pax2+ cells, kidney sections of P7 Wt1-Cre/Rosa26R mice were immunostained for Pax2 and β-galactosidase: in this line, Wt1-expressing cells and their progeny permanently express β-galactosidase [24]. To establish the percentage of Pax2+ cells that expressed β-galactosidase, the total number of Pax2+ cells within different fields of view were counted (total; 353), and the number that co-expressed β-galactosidase was determined (total; 161). The results showed that 45.6% of Pax2+ cells within the papilla expressed β-galactosidase, indicating that about half of these cells are derived from MM (Fig. 1B). β-galactosidase staining of P7 wild-type control mice showed no evidence of positive staining (Supplementary Fig. S1B).
Pax2+ cells can be expanded in culture and express key MM-specific markers
To establish if the Pax2+ cells in the postnatal mouse kidney were stem cells, it was first necessary to develop a culture system to enable their expansion in vitro. Disaggregated postnatal kidney cells were initially cultured on STO mouse feeder cells, which are known to support self-renewal of mouse embryonic stem cells, and in low serum-containing medium, which has been shown to support the culture of multipotent stem cells derived from mouse kidney [4]. The feeder cells were able to support expansion of the Pax2+ population during the first 7 days, but by day 28, no further expansion occurred and many of the kidney cells no longer expressed Pax2 (Fig. 2A). It has been shown previously that human kidney stem cells can be successfully expanded on FN [2]. Therefore, the performance of putative mouse kidney-derived stem cells on FN was compared to that on feeder cells. To circumvent the difficulties of performing gene expression analysis in the presence of feeder cell-derived mRNA, before seeding the kidney cells, feeder cells were treated with deoxycholate to remove all cellular material apart from extra-cellular matrix [25]. Cells on both FN and deoxycholated-STO (dSTO) substrates were cultured in the presence of STO-conditioned medium (STO-CM). Immunostaining for Pax2 and Wt1 was performed on putative kidney-derived stem cells cultured on FN and dSTO after 2 and 5 months. After 2 months, almost all cells on both substrates expressed Pax2 and Wt1 protein (Fig. 2B). However, after 5 months, while cells cultured on FN continued to express both MM-specific proteins, those on dSTO no longer expressed Pax2 (Fig. 2B). RT-PCR analysis showed no differences in MM-specific gene expression at the earlier time point (Fig. 2C), but after 5 months, while cells cultured on FN continued to express the MM-specific markers Pax2, Wt1, Gdnf, and Sall1, those on dSTO no longer expressed Sall1, and Pax2 was barely detectable (Fig. 2D). The same results were obtained with 2 independent isolations. These results show that a population of putative kidney-derived stem cells expressing key MM-specific markers can be successfully expanded in vitro if cultured on FN in STO-CM. However, although the cells expressed MM-specific markers, we could not rule out the possibility that some or all of the cells within the population, were derived from UB, given that both the MM and UB cells express Pax2. It is well established that the MM generates cells of the nephron, whereas the UB generates cells of the collecting ducts. Therefore, to investigate if MM and/or UB-like cells were present in the population, the cells were cultured under differentiating conditions and screened for the expression of various nephric and collecting duct markers (see below). Previous studies have shown that stem/progenitor cells derived from human kidney express the pluripotency marker, Oct4, both in vivo and in vitro [26]. However, using mouse embryonic stem cells as a positive control, we have found that Oct4 is expressed neither in neonatal or adult mouse kidney in vivo, nor in the Pax2+ population in vitro (results not shown).

Pax2+ cells can be expanded in culture and express key metanephric mesenchyme (MM)-specific markers.
Under differentiating conditions, putative kidney-derived stem cells give rise to podocyte- and proximal tubule-like cells
To investigate if the putative stem cells could generate renal-specific cell types, after 5 months of culture under the aforementioned expansion conditions, they were switched to differentiation-promoting conditions by withdrawing STO-CM and FN and increasing the amount of fetal bovine serum. After 4 weeks under differentiating conditions, the putative stem cell population became heterogeneous and a more diverse range of cell morphologies were observed (Supplementary Fig. S2A). Immunostaining and RT-PCR analysis showed that following the switch to differentiating conditions, Pax2 and Sall1 were no longer detectable, but Wt1 and Gdnf continued to be expressed (Supplementary Fig. S2B, C). In some cells, especially those that were binucleate, the levels of Wt1 protein were increased (Supplementary Fig. S2B). Given that podocytes express high levels of Wt1 and frequently become binucleate [15], the appearance of cells with these characteristics suggested that podocyte-like cells had spontaneously differentiated. Further, at lower plating densities, cells with a highly arborized morphology were observed, which is a typical characteristic of podocyte cells (Fig. 3A), and groups of cells with an epithelial-like morphology were also present in the population (Fig. 3B). To determine if other glomerular-specific cell types were present, immunostaining was performed to detect the mesangial marker, α-smooth muscle actin (αSMA) [5]. After culture under differentiating conditions, some cells within the population started to express αSMA (Fig. 3C). Immunostaining for cytokeratin, a marker for epithelial cells of the renal tubules [5], showed that cytokeratin-positive cells were only present after culture under differentiating conditions (Fig. 3D). This result suggested that in addition to podocyte-like cells, the cells were also able to generate renal tubular epithelial cells. To investigate if these epithelial cells displayed proximal tubule-like characteristics, the cultures were assayed for activity of alkaline phosphatase, an enzyme specific for PTCs [19]. Analysis showed that the enzyme was expressed in epithelial-like cells after culture under differentiating conditions (Fig. 3E). These results suggested that under differentiating conditions, the putative stem cell population was able to generate podocyte- and mesangial-like cells. Interestingly, although the undifferentiated cultures did not contain any cells with the typical morphological characteristics of podocytes or epithelial cells, or any cells that expressed the mesangial protein, αSMA, or the PTC proteins, cytokeratin, and alkaline phosphatase, RT-PCR analysis showed that both undifferentiated and differentiated populations expressed a number of podocyte markers, including podocalyxin and synaptopodin, the mesangial marker, desmin, and the PTC marker, megalin [27] (Fig. 3F). We failed to detect expression of the distal tubule marker, uromodulin, or the collecting duct marker, aquaporin-2 (AQP2) [28] under any culture conditions (results not shown). These data show that the putative stem cell population is able to generate podocyte-, mesangial-, and proximal tubule-like cells but does not appear to generate distal tubule or collecting duct cells.

Putative stem cell cultures give rise to podocyte, mesangial, and proximal tubule-like cells.
Putative kidney-derived stem cells are clonogenic
The previous results suggested that the putative stem cell population either contained multipotent stem cells with the potential to become podocytes, mesangial cells, and PTCs, or contained unipotent progenitor cells that were committed to become 1 of these 3 cell types. To determine which of these scenarios was correct, single cells were plated in 96-well plates in order to establish clonal lines (Supplementary Fig. S3). Although some of the clonal lines, such as clones H9 and E5, were completely homogeneous and did not express Pax2 (Fig. 4A, D) or generate proximal tubule- or podocyte-like cells, after expansion, other lines, such as H6, generated a heterogeneous population of cells, some of which had the characteristic podocyte- and epithelial-like morphologies that were present in the bulk culture (Fig. 4A). Unlike human-derived kidney stem/progenitor cell lines that require morphogens, such as retinoic acid, to direct their differentiation to podocytes and proximal tubule-like cells [6], some cells within the H6 population readily underwent spontaneous differentiation to these cell types in the absence of exogenous morphogens. Further, apart from their ability to undergo spontaneous differentiation, the H6 cells also expressed the MM markers, Pax2 and Six2 (Fig. 4C), and were therefore selected for further characterisation.
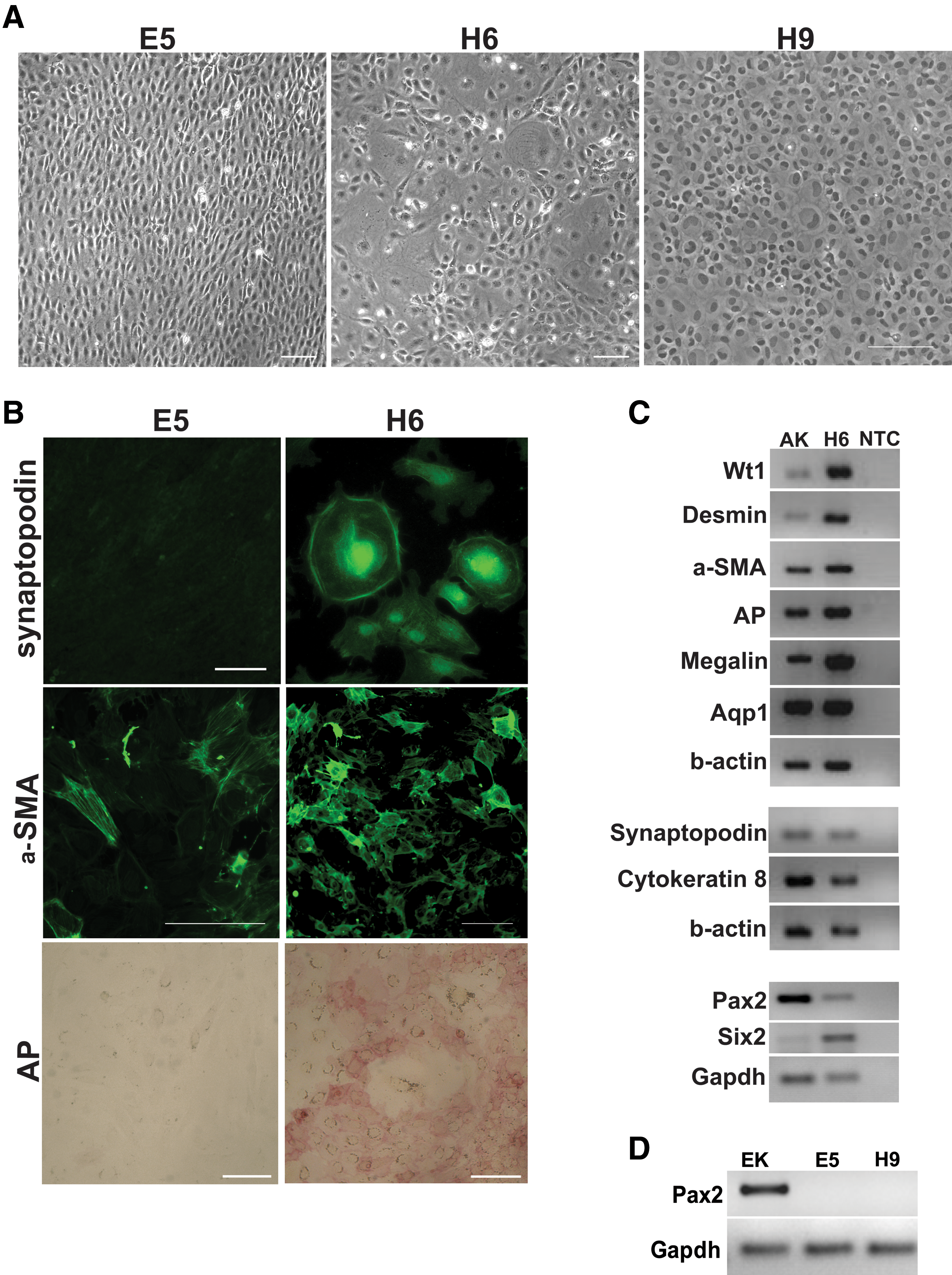
The kidney-derived cells are clonogenic and the H6 clonal line can generate podocyte-, mesangial-, and proximal tubular-like cells.
Characterization of the H6 clonal line
To investigate if the H6 line expressed podocyte-specific markers, immunostaining was performed for synaptopodin, a marker for mature podocytes [29]. For comparison, immunostaining was also performed on the homogeneous line, E5. To promote differentiation, both lines were cultured under the previously described differentiation promoting conditions. Cells with intense staining for synaptopodin were only observed in the H6 population. Likewise, cells with alkaline phosphatase activity were only detected within the H6 population. However, both cell lines expressed the mesangial marker, αSMA (Fig. 4B). RT-PCR analysis showed that the H6 line expressed the podocyte markers, Wt1 and synaptopodin; the mesangial marker, desmin; the renal tubule marker, cytokeratin; and the proximal tubule markers, megalin [30] and aquaporin-1 (AQP1) [28] (Fig. 4C), whereas distal tubule and collecting duct markers were not detected (results not shown).
To investigate the expression of stemness markers in the H6 cells, immunostaining was performed for the pluripotency markers, Oct4 and Sox2, and for the adult stem cell marker, musashi1. Expression of the pluripotency gene, nanog, was investigated using RT-PCR. The results showed that while the H6 line did not express Oct4, some cells within the population expressed Sox2 and musashi1. Further, compared to adult kidney, the H6 line expressed high levels of nanog (Supplementary Fig. S4).
Previous studies have shown that stem cells isolated from rodent and human kidney express sca-1 and CD24, which are also expressed by many mesenchymal stem cell (MSC) populations, but express only low levels of hemopoietic stem cell markers [5,9]. Cytofluorimetric analysis showed that the stem cell markers sca-1 and CD24 were expressed by ∼30% of the H6 kidney-derived stem cell (KSC) population (Fig. 5A, B). The MSC markers β1-integrin (CD29) and CD44 were expressed in >90% of H6 cells (Fig. 5C, D), whereas the hemopoietic stem cell markers CD45 and CD117 were expressed by <20% of cells within the population (Fig. 5E, F). To investigate if the H6 line could generate nonrenal cell types, the cells were cultured in adipogenic and osteogenic media and stained for oil red-O and alizarin red, respectively. We found that although there was no evidence of spontaneous differentiation of nonrenal cell types, after culture in the appropriate inductive medium, the H6 line readily underwent adipo- and osteogenesis (Fig. 6). The H6 clonal line was cultured continuously for >12 months (>200 population doublings), during which time it continued to spontaneously generate podocyte- and proximal tubule-like cells and did not undergo senescence.
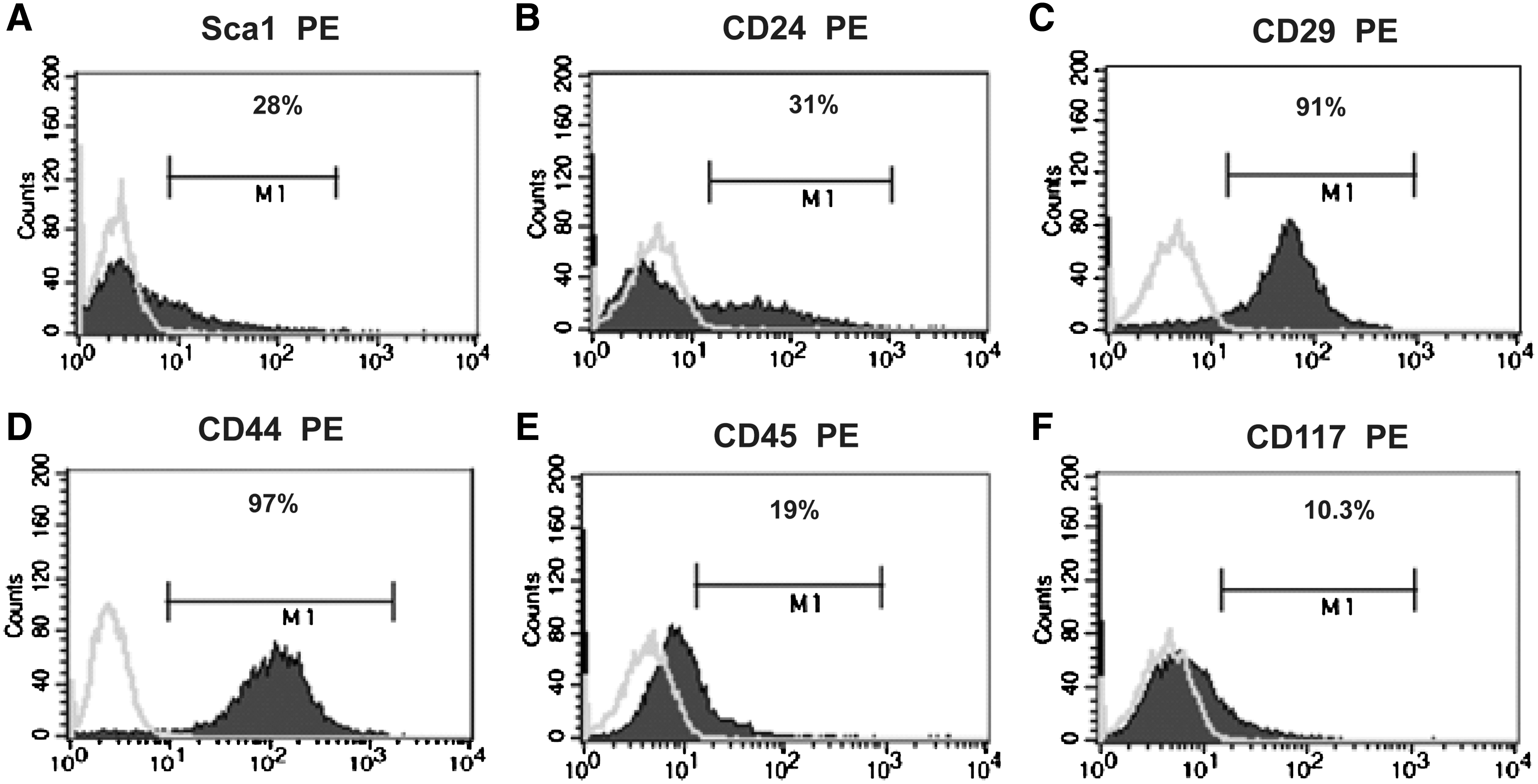
FACS analysis of surface stem cell markers in the H6 line.

The H6 clonal line can undergo adipogenesis and osteogenesis.
Discussion
Our results show that the renal papilla of the postnatal mouse kidney contains a population of MM-derived Pax2+ cells that, after expansion in culture, expressed several MM- and stem cell-specific markers. Under differentiating conditions, some cells within the population spontaneously differentiated and gave rise to mesangial-, proximal tubule-, and podocyte-like cells. Importantly, the morphology of these podocyte-like cells was very similar to that of primary podocytes in culture [15]. The cells were clonogenic, and the H6 clonal line expressed MM genes, the kidney stem cell markers, sca-1, CD24, and musashi1, and the pluripotency markers, Sox2 and nanog. Further, H6 cells had limitless proliferative capacity, and displayed the same differentiation potential as the original bulk culture, including the ability to generate nonrenal cell types. Taken together, these characteristics indicate that the H6 kidney-derived stem cell line comprises a true stem cell population. To our knowledge, this is the first time that a kidney-derived stem cell line has been found to generate cells with the typical marker expression profile and morphological characteristics of podocytes and PTCs in culture.
Over recent years, several stem cell populations have been identified in rodent and human kidney. These stem cells have been identified in various locations within the kidney, suggesting that they might represent distinct cell populations that are perhaps derived from different sources. For instance, in rat, stem cells identified by BrdU retention have been identified both in the papilla [1,7] and in proximal tubules within the region of the cortico-medullary junction (CMJ) [3,8]. However, despite their different locations, both papillary and CMJ-derived cells could engraft into renal tubules after ischemia-reperfusion injury (IRI) [1,3], suggesting that the apparent differences in intrarenal distribution might have simply been due to variations in the protocols of the 2 groups; in particular, in the age of the experimental animals that were used. In support of this, a recent study has shown that BrdU-retaining cells are present in both the papilla and CMJ of the mouse kidney [31]. Apart from BrdU-retention, Sca-1 expression has also been used to identify stem cells in the mouse kidney [9]: in agreement with our findings and the studies of Oliver et al. [1,7], it was found that the majority of Sca-1+ stem cells were present in the papilla, and could also populate the renal tubules after IRI [9]. Using a different approach based on the use of restrictive culture conditions, Gupta et al. were able to establish a stem cell population that could also populate the tubules after IRI [4]. The ability of all the aforementioned kidney stem cell lines to populate renal tubules in vivo suggests that they are indeed resident stem cells, rather than MSCs that have localized to the kidney, for MSCs display a limited ability to generate renal tubule cells after transplantation into rodents with induced renal injury [32]. In the current study, we show that a Pax2+ cell population is present in the mouse renal papilla. Burg and co-workers have also shown that Pax2+ cells are located in the mouse renal papilla, where they are present in both the thin descending loop of Henle, and in the collecting tubules [33]. During development, the loop of Henle, glomerulus, and proximal and distal tubules are generated from the MM, whereas the collecting tubules are generated from the UB. Given that UB cells are unable to generate nephric cell types [34], it was important in this study to determine if any of the Pax2+ cells present in the papilla were of MM origin. Using a Wt1-Cre/Rosa26R mouse line, we show here that ∼50% of the Pax2+ cells in the papilla are derived from Wt1+ cells, confirming that they are derived from the MM, and therefore could have the potential to generate nephric cell types.
The most striking result of our study was that the H6 clonal line can spontaneously generate podocyte- and PTC-like cells. Although most of the rodent kidney stem cells isolated to date can generate renal tubular cells in vivo [1,3,4,7,9], and some can generate nonrenal cells in vitro [1,4,9], as yet, it has not been demonstrated that these cells can generate PTCs in vitro. Further, it has not previously been demonstrated that rodent kidney stem cells can generate podocytes in vivo or in vitro. The lack of podocyte differentiation in vivo might be due to the particular renal injury model that is commonly used, which is usually the warm IRI model. In warm IRI, it is mainly the PTCs that are damaged and the podocytes are left intact [35]: it is therefore not surprising that podocyte differentiation was not observed in these studies. Previous studies have shown that human kidney stem cell lines upregulate podocyte-specific markers in vitro after induction with retinoic acid, but it is important to note that morphologically, these cells do not resemble true primary podocytes [5,6]. However, it has recently been demonstrated that CD133+ CD24+ kidney stem cells isolated from human Bowman's capsule are bipotent stem cells that can generate podocytes and tubular cells in a mouse model of adriamycin-induced nephropathy [6], an injury model that principally affects podocytes but also causes tubular damage: this supports the view that previous difficulties in generating podocytes from kidney stem cells in vitro has probably been due to deficiencies in the culture environment.
Characterization of the surface markers expressed by the H6 cells in vitro showed that most cells within the population expressed the MSC markers CD29 and CD44. Further, the H6 cells shared some of the differentiation potential of MSC, as they were able to generate osteocytes and adipocytes under appropriate culture conditions. However, unlike MSCs, 10%–20% of cells within the H6 population expressed the haemopoietic markers, CD45 and CD117. Interestingly, when kidney rudiment cells derived from early stage rat fetuses are transplanted into adult mouse kidneys, they give rise to various nonrenal cell types, including chondrocytes and osteocytes—cell types that are more commonly derived from MSCs [36]. Moreover, it has been shown that if MSCs are transplanted to the site of the presumptive kidney in the early stage rat embryo, they are able to generate a range of kidney-specific cell types [37]. The similarities in the developmental potential of H6 cells, early stage kidney rudiment cells, and MSCs might reflect the fact that they originate from the same subpopulation of mesoderm during embryogenesis, for the kidney and the aorta-gonad-mesonephros region, where MSCs first originate [38], are both derived from the intermediate mesoderm.
In conclusion, we have shown that the postnatal mouse kidney contains a population of multipotent stem cells that are able to spontaneously differentiate in vitro to cells that have the typical characteristics of PTCs and podocytes. Further studies are needed to evaluate if these cells have potential for drug screening, and for elucidating the mechanisms underlying PTC and podocyte differentiation during embryogenesis.
Footnotes
Acknowledgments
This work was supported by the Alder Hey Children's Kidney Fund and the EC 6th Framework program grant KIDSTEM under contract number 036097-2.
Author Disclosure Statement
The authors declare that they have no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.