
Abstract
Prenatal transplantation of genetically engineered mesenchymal stem cells (MSCs) might benefit prevention or treatment of early-onset genetic disorders due to the cells' intrinsic regenerative potential plus the acquired advantage from therapeutic transgene expression. However, a thorough assessment of the safety, accessibility, and behavior of these MSCs in the fetal environment using appropriate animal models is required before we can advance toward a clinical application. We have recently shown that fetal rabbit liver MSCs (fl-MSCs) have superior growth rate, clonogenic capability, and in vitro adherence and differentiation abilities compared with adult rabbit bone marrow MSCs. In this follow-up study, we report safe and widespread distribution of recombinant pSF-EGFP retrovirus-transduced fl-MSCs (EGFP+-fl-MSCs) in neonatal rabbit tissues at 10 days after fetal allogeneic transplantation through both intrahepatic and intra-amniotic administration. Conversely, a more restricted biodistribution pattern according to the route of administration was apparent in the young rabbits intervened at 16 weeks after fetal EGFP+-fl-MSC transplantation. Furthermore, the presence of these cells in the recipients' tissues, tracked with the reporter provirus, was inversely related to the developmental stage of the fetuses at the time of intervention. Long-term engraftment was confirmed both by fluorescence in situ hybridization analysis on touch tissue imprints using a chromosome Y-specific BAC probe, and by immunohistochemical localization of EGFP expression. Finally, there was no evidence of immune responses against the transplanted EGFP+-fl-MSCs or the EGFP transgenic product in the treated young rabbits. Thus, cell transplantation approaches using genetically engineered fetal MSCs may prove particularly valuable to frontier medical treatments for congenital birth defects in perinatology.
Introduction
W
Due to their recently described multilineage differentiation, and regenerative and immunomodulatory capabilities, mesenchymal stem cells (MSCs) are also being actively explored for the treatment of established nonhematopoietic diseases, including musculoskeletal defects [4 –7], and to improve hematopoietic stem cell engraftment [8].
Additional efforts are being put into developing preclinical MSC transplantation studies in relevant animal models to advance toward the early treatment of many genetic disorders before definitive organ damage has occurred. Thus, human MSCs from several sources have been shown to reconstitute different tissues after human–sheep [9 –11], human–mouse [12,13], or human–rat [14] xenogeneic in utero transplantation. Furthermore, proof-of-principle studies have recently stressed the therapeutic potential of human fetal MSCs for the treatment of severe osteogenesis imperfecta both after xenogeneic transplantation into the corresponding mouse model [15], and after allogeneic transplantation in an immunocompetent, HLA-incompatible patient [16].
According to several reports [17 –19] and from our own experience [20,21], the pregnant rabbit is a useful and reliable middle-sized animal model to evaluate proof-of-principle strategies based on gene transfer or cell transplantation into the fetus, which allow accurate organ-directed manipulation and delivery. Using fetal instead of adult donor cells for in utero transplantation approaches may also offer some benefit. Indeed, fetal MSCs from different tissue sources are present in higher amounts than their adult MSC counterparts, and show superior proliferative capacity once isolated [22,23].
We have recently characterized MSCs isolated from the fetal rabbit liver (fl-MSCs), which showed superior growth rate, clonogenic capability, substrate adherence, and transduction efficiency over MSCs isolated from adult rabbit bone marrow [23]. In this follow-up report, we confirm the nontumorigenic nature of fl-MSCs and analyze the potential of genetically engineered fl-MSCs for in vivo engraftment and persistence into rabbit tissues after fetal transplantation at different stages of development.
Materials and Methods
fl-MSC isolation, amplification, and characterization
fl-MSCs were isolated from the livers of 7 rabbit fetuses from a single White New Zealand pregnant rabbit at 19 days of gestation. The doe was anesthetized with a subcutaneous injection of 15 mg/kg ketamine (Ketolar® 50; Pfizer) plus 0.25 mg/kg medetomidine (Domtor®; Pfizer). The abdomen was shaved and disinfected with iodine and the uterine horns were exposed by laparotomy through a midline abdominal incision. Seven intact fetal sacs were harvested and transferred to a cell culture laminar flow cabinet, and the corresponding fetuses were euthanized by decapitation under anesthesia coming from the mother. The livers were extracted by mechanical disgregation and homogenization. The corresponding single-cell suspension was washed with phosphate-buffered saline (PBS) and resuspended in α-MEM (Invitrogen) containing 20% fetal bovine serum, 1% penicillin/streptomycin, 1% L-glutamine, and 2.5 μg/mL amphotericin (all from Invitrogen), and deposited overnight in culture flasks (Corning) (1×105 cells/cm2) at 37°C in a 5% CO2 atmosphere.
Next day, the cell monolayer was washed once more with PBS to remove nonadherent cells and cellular debris, and a fresh medium was added. Finally, the fl-MSCs were amplified for 10–12 days, replacing the culture medium every 3–4 days, until 90% confluent.
The growth, immunological properties, and differentiation capability of fl-MSCs have been recently characterized [23].
fl-MSC transduction and sorting
The gamma retroviral vector pSFb91-EGFP (pSF-EGFP; kindly provided by Christopher Baum, University of Hamburg, Germany) [24], expressing the enhanced green fluorescent protein (EGFP), was employed to generate vesicular stomatitis virus G-glycoprotein (VSV-G)-pseudotyped recombinant retrovirus particles by transient transfection, as recently described [25].
Transduction of fl-MSCs (passage 1) was performed by spinoculation. Briefly, 5×105 fl-MSCs and 1×105 viral vector particles (multiplicity of infection [MOI]: 0.2) were co-cultured on a 6-well cell culture plate (Corning) in 2 mL/well α-MEM complete medium supplemented with 8 μg/mL of polybrene (Sigma), and centrifuged at 1,000 g for 90 min at 32°C. After overnight incubation, the cell culture supernatant was replaced by af resh medium and the cells were cultured for another 24 h, yielding transduction efficiency close to 1%. This ensured that most of the transduced fl-MSCs contained a single-copy provirus.
These fl-MSCs were excited at 488 nm, sorted through a 530/40 band pass filter, and recovered as a bulk population of 1.8×105 fluorescent fl-MSCs (EGFP+-fl-MSCs) using a MoFlo® jet-in-air high-speed sorter (Beckman-Coulter). These cells (mean fluorescence intensity [MFI]: 103–104) were amplified to passage 5 before their transplantation into fetal rabbits.
In vivo tumorigenic assay
The tumorigenic potential of fl-MSCs was tested in 8-week-old male SCID mice. Seven mice were administered 5×106 fl-MSCs resuspended in 40 μL PBS through intramuscular injection in their lower right limb. Two control mice were administered the same volume of PBS. All administrations were carried out using protocols approved by the Animal Care Committee.
Animals were examined twice a week and weighted weekly. After 13 weeks, all mice were sacrificed and samples from liver, kidney, gut, intestines, heart, lung, spleen, testicles, and lower left and right limb were fixed in formalin and analyzed by an expert veterinary pathologist (Histovet Histological Diagnostic Service, St. Quirze del Vallès, Barcelona, Spain).
Animals and transplantation procedures
Timed pregnant White New Zealand rabbits (13–15 days of gestation) were housed for 1 week before surgery. The housing was quiet, at constant room temperature and humidity. Laparotomies were performed under anesthesia at day 17 (E17) or day 21 (E21) of gestation (gestation=31–32 days) as described previously [20,21]. The fetus was located and fixed by external palpation of the semitransparent uterine wall, and intra-amniotic or intrahepatic injections were performed using 26-gauge needles in view of the common anatomical landmarks of the E15–22 rabbit embryos [26]. To ensure intrahepatic injection, the fetus was punctured at the lower level of the thorax. Intra-amniotic administration was made directly in the amniotic fluid of the fetal sac. The estimated accuracy rate for both administration routes was 85%–90%, assessed by Fast Green FCF staining [21].
Approximately 20 min before the intervention, the fl-MSC-EGFP+ transductants were trypsinized and resuspended in PBS. The fetuses administered at gestation day 17 received a unique 100 μL dose of 3.5×105 fl-MSC-EGFP+ cells, whereas those administered at gestation day 21 received a unique and comparable statural growth-adjusted 100 μL dose of 5×105 fl-MSC-EGFP+ cells [26]. Control fetuses were injected with an identical volume of PBS. No more than 50% of the fetuses were treated per uterine horn (the vaginal-end sacs were always excluded). Whenever possible, injection of adjacent fetuses was avoided. Once the injections were completed, the uterus was returned to the abdominal cavity and the abdomen was closed in layers. Does were kept under a warming blanket until awake and active, and received subcutaneous injections of meloxicam (Metacam; Abbott Laboratories) (0.4 mL per rabbit/24 h) over 48 h as postoperative analgesia.
All animal interventions were conducted in compliance with applicable regulations and guidelines, and were reviewed and approved by the Animal Care Committee
Tissue collection and processing
On day 31 of gestation (10 and 14 days after intervention for pups administered on E21 and E17, respectively) pregnant rabbits were sacrificed with a euthanizing dose of pentobarbital (150 mg/kg, intravenous). Samples from gonads and uterus from each intervened doe were frozen for DNA extraction.
Experimental pups were retrieved by visual identification after cesarean section, reanimated, and weighted. Those destined to the short-term study were euthanized by decapitation, and tissue samples from gonads, kidney, liver, heart, lung, trachea, esophagus, muscle, skin, thymus, brain, spleen, intestine, and placenta were washed with PBS and quickly frozen in liquid nitrogen for subsequent nucleic acids extraction. The pups destined for the long-term study were marked with a subcutaneous microchip (AVID Microchip ID Systems) for a further identification, kept in an incubator C100 (Air Shields) at 32°C for approximately 1 h, and finally allocated to foster rabbits. At 16 weeks after birth, the rabbits were sacrificed with a euthanizing dose of pentobarbital as described above. The appropriate tissue samples were harvested, including gonads, kidney, liver, heart, lung, trachea, esophagus, and placenta. A sample fraction of each tissue was quickly frozen in liquid nitrogen for nucleic acids extraction. The remaining fraction was fixed overnight in formalin and embedded in paraffin for immunohistochemical and fluorescence in situ hybridization (FISH) evaluation.
DNA isolation and polymerase chain reaction analysis
Genomic DNA was obtained from fetal tissues by standard phenol-chloroform extraction and isopropanol precipitation. Briefly, samples were washed twice in 1×saline-sodium citrate buffer (SSC) and 10 mM ethylenediaminetetraacetic acid (EDTA), and incubated overnight at 56°C in sterile lysis buffer (10 mM Tris-Cl, pH 10.5; 1 mM EDTA; 0.15 mM NaCl; 0.5% sodium dodecyl sulfate; 0.3 mg/sample proteinase K; Roche) before phenol extraction. The vector presence in the harvested organs was determined by polymerase chain reaction (PCR) amplification of a 801 bp provirus-specific DNA fragment carried out in a 25 μL volume containing 50 ng of genomic DNA, 100 μM dNTPs, 2.5 μL 10×Biotaq DNA polymerase buffer, 1.50 mM MgCl2, 0.15 μL Biotaq DNA polymerase (5 U/μL) (Bioline), and 20 pmol of the following primers: pSF-F (5′-CTC CGC CAC TGT CCG AGG GGT-3′), localized in the retroviral leader sequence, and EGFP-R (5′-TGG GGG TGT TCT GCT GGT AGT G-3′), localized in the EGFP sequence, in a GeneAmp® PCR system 2400 DNA thermocycler (Perkin-Elmer). A control PCR containing no DNA to monitor for contamination of PCR reagents and a positive control containing vector DNA were included in each PCR analysis. Amplification conditions included an initial 10 min denaturation step at 95°C, followed by 40 cycles (95°C, 45 s; 63°C, 45 s; 72°C, 1 min), and a final 7 min extension at 72°C. Rabbit skeletal muscle myosin heavy chain gene (MIO) primers, RMMHC-F (5′-AAAGAAGATGGATGTTGAGGC-3′), and RMMHC-R (5′-TCACCGTCACTTTCCCTGCT-3′), were used in parallel to amplify a 429 bp-specific DNA fragment [20], which served as a control for genomic DNA isolation and loading. All samples were evaluated through 2% agarose gel electrophoresis.
Furthermore, to corroborate EGFP+-fl-MSC-derived retroviral vector biodistribution in the treated rabbit's tissues, all above samples were re-analyzed by PCR amplification of a 422 bp fragment using primers: PSF IN 5′ (5′-TCTGCAC CACCGGCAAGCTG-3′), and the previously described EGFP-R, both within the EGFP sequence.
Additionally, genomic DNA from the in utero EGFP+-fl-MSC-transplanted young rabbit tissues (100 ng/each) was analyzed by real-time quantitative TaqMan polymerase chain reaction (TaqMan qPCR; PE Applied Biosystems) to estimate the provirus copy number within each tissue. Specific primers F1EGFP (5′-GTAAACGGCCACAAGTTCAGC-3′) and R1EGFP (5′-TGGTGCAGATGAACTTCAGGG-3′), and TaqMan MGB probe PEGFP (5′-CTTGCCGTAGGT GGC-3′) amplified and detected the retroviral transgenic EGFP sequence. All PCRs were performed in a 20 μL reaction mixture containing 10 μL of TaqMan Universal PCR master Mix (PE Applied Biosystems), 45 nM of each primer, and 12.5 nM fluorogenic probe. Amplification conditions involved an initial 10 min denaturation step at 95°C, followed by 40 cycles (95°C for 30 s; 58°C for 30 s). Amplifications were carried out in 96-well reaction plates (PE Applied Biosystems) in a spectrofluorimetric thermal cycler (ABI PRISM 7300 Sequence Detector; PE Applied Biosystems). Provirus copy number was assessed using a standard curve generated by mixing the pSF-EGFP plasmid with nontransduced rabbit genomic DNA at different ratios, ranging from 106 to 1 provirus genome copies/100 ng of rabbit genomic DNA. As positive controls, we employed genomic DNA from K562 cell clones containing 1 or 2 provirus copies of pSF-EGFP [25]. A control PCR containing no DNA to monitor contamination of PCR reagents was included in each PCR. All samples were run in triplicate.
Fluorescent in situ hibridization
Previous to the in utero administration of EGFP+-fl-MSCs, we estimated the percentage of male and female cells within the sorted bulk population. To this end, we performed a FISH analysis using 2 BAC clones as specific probes, either for the rabbit X chromosome (530F8 at Xq26.1) or for the rabbit Y chromosome (828D7 at Yp12) [27]. BAC 530F8 was labeled with Spectrum Green, and BAC 828D7 with Spectrum Red (all from Vysis) by a standard nick translation reaction. FISH was performed as described elsewhere [28]. Briefly, 400 ng of labeled probes was co-hybridized overnight on metaphase chromosomes obtained from a confluent plate of EGFP+-fl-MSCs. Post hybridization washes were performed through one change of 0.4×SSC/0.1% Tween 20 at 74°C, and one change of 2×SSC/0.1% Tween 20 at room temperature. Slides were mounted in Vectashield solution (Vector Laboratories) containing 150 ng/mL of 4′,6-diamidino-2-phenylindole dihydrochloride counterstain. FISH analysis was completed under an epifluorescence microscope (Olympus BX50) equipped with the appropriate filter set.
The presence of transplanted, EGFP+-fl-MSCs cells within the different in utero-treated rabbit tissues was assessed through FISH analysis using the rabbit Y chromosome probe (828D7 at Yp12) on paraffin-embedded tissues from 16-week female rabbits that were PCR positive for the presence of the pSF-EGFP provirus sequence.
Immunohistochemistry
Formalin-fixed tissues were dehydrated with an ethanol gradient, cut, and embedded in paraffin. Sliced 5-μm sections were mounted on poly-
Enzyme-linked immunosorbent assay
Serum for the long-term study was obtained from the young treated rabbits at 6 and 16 weeks after fl-MSC-EGFP+ administration. Blood was taken from the lateral saphenous vein and left coagulating for approximately 30 min. Samples were centrifuged twice at 3000 g for 10 min and kept at −80°C until their use.
The generation of a rabbit humoral immune response against the EGFP reporter protein was determined by enzyme-linked immunosorbent assay (ELISA). Maxisorb ELISA 96-well plates (Nunc) were coated overnight at 4°C with 0.1 μg/well of recombinant EGFP (BioVision) diluted in 50 μL of 50 mM carbonate buffer (pH 9.6). After several washes with PBS-T (PBS plus 0.05% Tween 20), plates were blocked with 300 μL of 3% fish gelatin (Sigma-Aldrich) diluted in PBS for 2 h at room temperature. Serum samples diluted 1:500 with diluting buffer (PBS-T+3% bovine serum albumin) were incubated in triplicate wells for 2 h at room temperature. The plates were then washed again with PBS-T and 100 μL of secondary antibody (biotin-conjugated goat anti-rabbit IgG, 1:200,000 dilution) (Sigma-Aldrich) was further incubated for 2 h at 37°C. Subsequently, plates were washed with PBS, incubated for 30 min with biotinylated horseradish peroxidase–avidin complex (ImmunoPure ABC Peroxidase Staining Kit; Pierce), and developed using 1-Step Slow TMB (Pierce) for 30 min. Finally, the reaction was stopped with 2 M H2SO4 and the corresponding absorbance was measured at 450 nm on an ELISA plate reader. A standard curve was prepared from serial dilutions of a rabbit polyclonal (IgG) anti-GFP antibody (MBL) in rabbit serum (prediluted 1: 500 with diluting buffer) to establish the range of sensitivity.
Mixed lymphocyte culture
Lymphocytes were isolated from the rabbit's spleens. After mechanical disgregation on a culture dish, spleens were homogenized by gentle flushing with a 21G needle. The cellular suspension was filtered through a 70 μm nylon filter to remove cellular debris, incubated for 30 min with lysis buffer (0.15 M NH4Cl, 10 mM KHCO3, 0.1 mM EDTA) at 4°C, and centrifuged at 300 g for 5 min. The lymphocyte pellets were washed twice with PBS and finally cultured in complete DMEM at 37°C and 5% CO2 atmosphere. Cell viability was assessed by trypan blue exclusion.
A 2-way mixed lymphocyte culture (MLC) was performed to study the effect of fl-MSCs on lymphocyte reactivity. Lymphocytes were seeded in triplicate at a concentration of 2×105 cells/well in 96-well U-bottom microtiter cell plates (Corning) in the presence or absence of 1×105 fl-MSCs (passage 2), preplated overnight before the experiment. After a 5-day incubation period, cell proliferation was determined using the Amersham cell proliferation biotrack ELISA system (GE Healthcare Amersham Biosciences).
Statistical analysis
Fetal weight comparisons were performed using the Student's t-test. The logarithm transformation was used to warrant normality. The percentages of E17- and E21-treated rabbits with short- and/or long-term presence of EGFP+-fl-MSC-derived provirus, and/or according to their route of administration, were compared using Fisher's exact test. Data are expressed as mean values±standard deviation. Statistically significant differences among fl-MSC-transplanted and control PBS-treated rabbits in both the ELISA assays and the lymphocyte proliferation assays were estimated using the nonparametric Mann–Whitney U test.
Results
Overall safety of fl-MSCs for fetal transplantation in the pregnant rabbit model
Rabbit fl-MSCs showed both a superior clonogenic capability and a higher proliferation rate compared with their adult bone marrow-derived counterparts [23]. Thus, before undertaking the planned in utero fl-MSC rabbit transplantation assays, we assessed the potential tumorigenicity of these cells in SCID mice. According to the outcome of several parameters (typical mouse behavior and growth evolution, absence of localized tumor formation and normal histology from tissue biopsies at the end of the 13-week follow-up period), no differences were observed between the fl-MSC-treated and the control PBS-treated SCID mice (data not shown). Hence, the isolated fl-MSC population did not exhibit tumorigenic potential.
Regarding EGFP+-fl-MSC fetal rabbit delivery, in terms of survival and regardless of the route and period of administration employed, a total of 52 fetuses from 11 pregnant does were intervened (41 were injected with EGFP+-fl-MSCs and 11 were injected with PBS) and only 2 abortions were documented among the EGFP+-fl-MSC-treated fetuses. Thus, the overall survival rate at birth was 95%. Furthermore, no anatomical defects were detected in any of the treated fetuses. They maintained a correct weight at the termination of pregnancy according to a normal developmental pattern (median weight of the EGFP+-fl-MSC-treated fetuses: 48.0+1.7 g, n=39; median weight of the control PBS-treated fetuses: 44.2+3.7 g, n=11) (P=0.3298; Student's t-test).
Therefore, neither the intrinsic features of the EGFP+-fl-MSCs nor the delivery procedures became noxious to the in utero-treated rabbit fetuses beyond the standard risk from the surgical interventions.
Efficiency and persistence of EGFP+-fl-MSC transplantation
We wished to evaluate the biodistribution and potential engraftment of fl-MSCs into allogeneic fetal tissues, and to analyze the influence of the developmental stage on fl-MSC homing. Therefore, we administered EGFP+-fl-MSCs into fetal rabbits at both E17 (intra-amniotically) and E21 (intra-amniotically and intrahepatically) of gestation. Their behavior was assessed at birth (for the E21-administered rabbits) and at 4 months after transplantation (for both the E17- and E21-administered rabbits). The presence of EGFP+-fl-MSCs in both the fetal and the adult tissues from the treated rabbits was reported by PCR-mediated amplification of integrated pSF-EGFP provirus sequences using genomic DNA isolated from the harvested tissues (Fig. 1A and Table 1).

Long-term assessment of allogeneic EGFP+-fl-MSC engraftment into young rabbit tissues after fetal transplantation (Part I): PCR-derived distribution assessment and FISH-mediated localization.
IH, intrahepatic; IA, intra-amniotic; N, number of EGFP+-fl-MSCs transplanted fetuses.
The presence of EGFP+-fl-MSC-derived proviral DNA in the different tissues from the in utero-treated fetuses were analyzed after genomic DNA extraction and PCR amplification using provirus-specific primers. For each tissue, frequencies are displayed as number of provirus-positive animals/total number EGFP+-fl-MSC-transplanted animals analyzed.
EGFP, enhanced green fluorescent protein; fl-MSC, fetal rabbit liver mesenchymal stem cell; ND, not determined.
Short-term study
Eighteen fetuses (7 EGFP+-fl-MSC-treated and 2 control PBS-treated, for each of the administration routes) were sacrificed at birth (10 days postadministration). All EGFP+-fl-MSC-transplanted fetuses except one, intra-amniotically-transplanted, contained pSF-EGFP-derived transgenic sequences in at least one of the tissues/organs analyzed. In contrast, tissues from the control PBS-treated fetuses were all negative for the presence of vector sequences (data not shown). Regardless of the route of administration employed, either intra-amniotic or intrahepatic, there was a widespread distribution pattern of EGFP+-fl-MSC-derived provirus sequences through all the analyzed tissues. The transplantation efficiency after intrahepatic administration was significantly higher than after intra-amniotic administration (P<0.0001; Fisher's exact test), although particular tissues seemed to show the opposite trend (eg, brain, intestine, and muscle). It should be noted that EGFP+-fl-MSC-derived provirus sequences were present in gonadal tissue from some EGFP+-fl-MSC-transplanted rabbits (frequency: 4/12, 33%), and from some individual placentas (3/8, 37.5%). In contrast, the does' tissues analyzed (uterus and gonads) were all negative for the presence of EGFP+-fl-MSC-derived pSF-EGFP provirus sequences (data not shown).
Long-term study
Rabbit pups were recovered by cesarean extraction 1 day before birth. After reanimation, 19 pups treated at E21 were placed in the care of a foster mother and 15 survived to adulthood: 12 were EGFP+-fl-MSC-transplanted (6 through intra-amniotic administration and 6 through intrahepatic administration), and 3 were control PBS-treated (2 through intra-amniotic administration and 1 through intrahepatic administration). Necropsies from the remaining 4 rabbit pups (1 intrahepatically EGFP+-fl-MSC-transplanted, 2 intra-amniotically EGFP+-fl-MSC-transplanted, and 1 intrahepatically control PBS-treated) discarded malformations or disease as the basis of nurturer rejection. Furthermore, all 13 recovered E17 intra-amniotically treated pups (10 were EGFP+-fl-MSC-transplanted, and 3 were control PBS-treated) survived to adulthood.
To assess the persistence of EGFP+-fl-MSCs after fetal transplantation, treated young rabbits were sacrificed at 16 weeks postintervention and freshly extracted tissues were examined for the presence of EGFP+-fl-MSC-derived provirus (Fig. 1A and Table 1). As in the short-term study, intrahepatic administration appeared more efficient than intra-amniotic administration (P=0.02; Fisher's exact test). Interestingly, close to 50% reduction in the percentage of animals with evidence of EGFP+-fl-MSC-derived provirus, independently of the administration route, was apparent when comparing both the short-term and the long-term studies (P<0.0001; Fisher's exact test). In consequence, low-level pSF-EGFP-derived provirus present in the EGFP+-fl-MSC-transplanted animals at long term reflected a progressive extinction of the delivered EGFP+-fl-MSCs.
Importantly, among the intra-amniotically administered animals, 60% of those transplanted at E17 and only 33% of those transplanted at E21 had detectable EGFP+-fl-MSC-derived provirus at 16 weeks after EGFP+-fl-MSC delivery. Therefore, the stage of development at the time of transplantation was inversely related to the efficiency of EGFP+-fl-MSC homing/engraftment (P=0.0002; Fisher's exact test). Furthermore, the young rabbits analyzed long-term showed a more restricted EGFP+-fl-MSC biodistribution pattern than the pups analyzed short-term (eg, regarding EGFP+-fl-MSC-derived provirus presence 16 weeks after transplantation at E21, intra-amniotic administration was limited to lung, trachea, and esophagus, whereas after intrahepatic administration it was present primarily in the liver). Again all PBS-treated control animals were completely negative for the presence of EGFP+-fl-MSC-derived provirus sequences (data not shown).
Finally, when evaluated by real-time quantitative PCR, the number of transplanted EGFP+-fl-MSCs remaining in all of the tissues/organs from the recipient young rabbits at 16 weeks post-transplantation was low (below the sensitivity of the assay: 1 provirus copy/ng genomic DNA).
Engraftment of EGFP+-fl-MSCs in fetal organs
Fetal homing and engraftment of EGFP+-fl-MSCs into the different rabbit tissues persisting to adulthood was monitored both by FISH analysis and by transgenic EGFP expression of tissue sections positive for the presence of pSF-EGFP provirus sequences from the young EGFP+-fl-MSC-treated rabbits.
We first estimated the proportion of male (47%) and female (53%) cells from the sorted, EGFP+-fl-MSC pool to be used for allogeneic fetal transplantation using rabbit chromosome X- and chromosome Y-specific BAC probes. The FISH analysis was further performed over EGFP+-fl-MSC-transplanted female rabbit tissues employing the Y-specific probe. The presence of engrafted EGFP+-fl-MSCs was evident as fluorescent dots seen in interphase nuclei from several tissues harvested from the in utero fl-MSC-treated young rabbits irrespective of their developmental stage at the time of fetal transplantation (Fig. 1B).
We also confirmed the presence of engrafted, allogeneic EGFP+-fl-MSCs in post-natal, in utero-treated rabbit tissues assessing transgenic EGFP expression through immunohistochemistry. The reporter EGFP protein was detected using an anti-EGFP antibody to enhance EGFP observation while avoiding the high background fluorescence present in some of the analyzed tissues [21]. The evaluation was performed in tissues from all PCR-positive animals containing the integrated pSF-EGFP provirus at 16 weeks post-treatment. Additionally, both PCR-negative tissues from EGFP+-fl-MSC-treated rabbits and from PBS-treated rabbit tissues were processed for control purposes. We should note that the presence of EGFP+-fl-MSCs detected by immunohistochemistry could underestimate the true proportion of engrafted cells owing to a degree of long-term transgene expression extinction perceived in pSF-EGFP-transduced MSCs in previous in vitro assays ([28], and data not shown).
In the animals treated through intrahepatic administration, long-term EGFP+-fl-MSC-derived cytoplasmic EGFP expression could only be detected in liver and kidney, and the signal was limited in both cases to a few patchy areas from the analyzed sections (Fig. 2). In the animals treated through intra-amniotic administration and regardless of the developmental stage, fetal breathing and swallowing of EGFP+-fl-MSCs led to concomitant long-term EGFP expression exclusively, although at low level, in some lung samples within arterioles and the bronchiolar tree (Fig. 2).
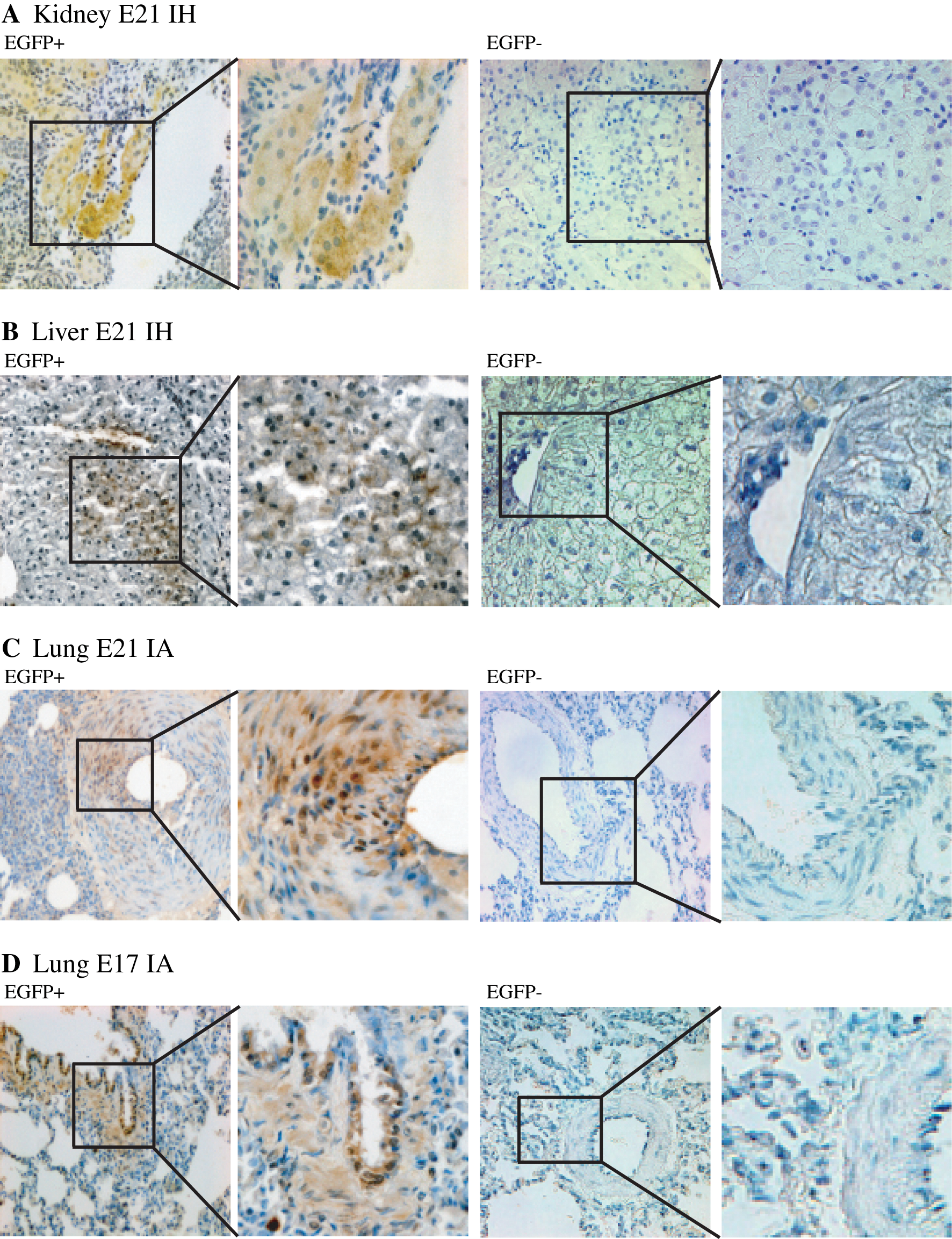
Long-term assessment of allogeneic EGFP+-fl-MSC engraftment into young rabbit tissues after fetal transplantation (Part II): immunohistochemical evaluation of transgenic EGFP expression. Immunostaining for EGFP-expressing cells was performed on PCR-positive tissues from EGFP+-fl-MSC-transplanted animals at 16 weeks postadministration [left paired images (EGFP+);×20 and×40, respectively]. The corresponding PCR-negative tissues both from EGFP+-fl-MSC-transplanted animals at 16 weeks postadministration [right paired images (EGFP−);×20 and×40, respectively], and from PBS-treated animals (not shown) were used for control purposes. Microscopic analysis of selected tissue sections according to the EGFP+-fl-MSC delivery route. EGFP+-fl-MSC-derived cells appear brown. Positive kidney staining of tubular epithelial-like cells
As previously reported [21], additional examination of the different tissue sections did not reveal the presence of cellular infiltrates in any of the analyzed samples, suggesting absence of immune-inflammatory activity.
Absence of immunity against allogeneic EGFP+-fl-MSCs and/or EGFP+-fl-MSC-derived transgenic EGFP after fetal transplantation
MSCs have been endowed with immunomodulatory properties. We have recently shown that fl-MSCs expressed both MHC class I-II and co-stimulatory molecules after short-term in vitro culture and induced proliferation of co-cultured allogeneic lymphocytes [23]. Therefore, we wished to assess whether young rabbits undergoing prenatal transplantation of allogeneic EGFP+-fl-MSCs would be able to develop a humoral immune response against the EGFP+-fl-MSC-derived EGFP transgenic product and/or to induce a cellular immune response against the fl-MSCs.
A specific ELISA to detect the presence of antibodies against EGFP was performed on sera harvested from the young rabbits at 6 and 16 weeks post-in utero EGFP+-fl-MSC transplantation. A single rabbit showing pSF-EGFP provirus integration in the lung developed anti-EGFP antibodies at 6 weeks after EGFP+-fl-MSC intra-amniotic administration, although there was no evidence of anti-EGFP antibodies in any of the animals analyzed at 16 weeks post-in utero transplantation (Fig. 3A).

Immunoregulatory capability of EGFP+-fl-MSCs.
Moreover, mononuclear cells isolated from the spleen of all but one of the in utero-transplanted young rabbits were unable to proliferate when exposed to allogeneic fl-MSCs (Fig. 3B). In this E21 intrahepatically treated rabbit, we could not verify the presence of the EGFP+-fl-MSC-derived pSF-EGFP provirus in any of the tissues analyzed at 16 weeks post-transplantation.
Discussion
Fundamental issues regarding the overall safety and therapeutic efficacy of ex vivo-manipulated allogeneic stem cell transplantation into the fetus mean that it requires rigorous testing in suitable animal models before its clinical application.
In the present study, we assessed the overall safety, biodistribution, and engraftment capacity of EGFP-expressing, genetically engineered fl-MSCs after allogeneic fetal transplantation into the pregnant rabbit model. We have shown the usefulness and relevance of this model in previous fetal gene transfer studies [20,21]. Indeed, there are several inherited disorders in which a rabbit model is already accessible, such as familial hypercholesterolemia [29], Pelizaeus-Merzbacher disease [30], and osteopetrosis [31]. All of them could represent good candidates for engineered allogeneic fl-MSC-based in utero therapeutic approaches.
Regarding the donor cells employed for fetal transplantation, we have recently assessed the optimal features of fl-MSCs for that purpose [23]. Besides sharing superior growth rate and ex vivo expansion, clonogenic efficiency and in vitro adherence over adult, bone marrow-derived MSCs, we speculated that fl-MSCs might take advantage of their ontogeny. Thus, the normal sequential developmental migrations of stem cells in the fetus might increase the homing and engraftment capability of fl-MSCs after cell transfer into fetal tissues (eg, hematopoiesis starts at the yolk sac or aorto-gonado-mesonephric region, migrates to the fetal liver, and finally resides in the bone marrow [32]). We [23] and others [22] have previously reported that fl-MSC possess at least some of the immunomodulatory properties found in adult-derived MSCs. In addition MSCs and, particularly, fetal MSCs are efficiently transduced with integrating vectors and express the corresponding transgene without losing stemness and differentiation capabilities [12,23,33].
We have confirmed that allogeneic in utero transplantation of retroviral vector-transduced rabbit fl-MSCs is safe both for the recipient dams and for the intervened offspring. Rabbit fl-MSCs have proven nontumorigenic when injected into immunodeficient mice. Numerous reports suggest that MSCs consistently pass into senescence and do not undergo transformation in culture, in contrast to immortal cells such as embryonic stem cells or induced pluripotent stem cells [34]. Additionally, none of the intervened rabbits showed any evidence of developmental abnormalities either in the short term or in the long term.
On the other hand, we have detected the presence of allogeneic EGFP+-fl-MSCs in the gonads of some treated rabbits at 10 days postfetal transplantation. However, none of the EGFP+-fl-MSC-transplanted young rabbits had evidence of donor cells in their reproductive organs when analyzed at long term, which implies absence of fl-MSC-derived germ line alterations in these animals. Finally, during the timeframe of our study, none of our in utero EGFP+-fl-MSC-transplanted rabbits showed any evidence of genotoxicity, although we cannot exclude the potential development of tumors at later time points. This important biosafety issue should warrant a careful comparative analysis of retroviral integration sites in a comprehensive genotoxicity study once the rabbit genome, scheduled for deep coverage by the mammalian genome project (
Concerning the overall biodistribution of allogeneic EGFP+-fl-MSCs into the transplanted fetuses, at 10 days post-transplantation we detected close to 100% cell transfer frequency and a widespread presence of EGFP+-fl-MSC-derived pSF-EGFP provirus sequences into most of the organs analyzed regardless of the administration route employed. The use of more accurate in utero targeted administration technologies, such as ultrasound biomicroscopy [35], should reduce delivery inconsistencies (cell misdelivery and/or spread) in diseases with focused affectation.
In comparison, the frequency of EGFP+-fl-MSC transfer was significantly reduced at 16 weeks after transplantation. What is more, at this time point, the EGFP+-fl-MSC biodistribution pattern appeared to be essentially restricted to the primary target organs according to the route of administration (liver for intrahepatic administration; lung, trachea, and esophagus for intra-amniotic administration). Several xenogeneic fetal transplantation studies have also reported short-term widespread MSC tissue biodistribution, although long-term analysis showed a drastic reduction or even complete loss of donor cell engraftment in the analyzed tissues [10,12,13]. Analogously, in utero transplantation of allogeneic fl-MSCs into sheep fetuses resulted in multiorgan, although low level, engraftment at 14 days post-transplantation [36]. It seems that competition against the developmentally active resident fetal MSC niches might constitute a significant barrier to successful allogeneic fl-MSC homing and/or engraftment, leading to their further elimination by physiological turnover, analogous to that described for the fetal hematopoietic system [37,38]. Alternatively, the allogeneic fetal microenvironment might support the viability and early differentiation of fl-MSCs, but not their replication, which would dilute the fl-MSC presence in adult rabbit organs at the time of analysis. Furthermore, the heterogeneity of the transplanted fl-MSC population with respect to differentiation potential or replication capacity and longevity might also influence the loss of tissue engraftment.
In injury models, MSCs can be recruited from remote storage sites to areas of wound healing [10,39]. Indeed, tissue injury seems to release soluble factors (TNF-alpha, SDF-1, etc.) capable of attracting and facilitating the homing of MSCs, which would enhance their repair/regeneration capabilities [40,41], and there are already a variety of preclinical studies and clinical trials exploring the repair/regeneration potential of MSCs in injured tissues (eg, sepsis, acute myocardial infarction, graft-versus-host disease, Crohn's disease, and acute lung injury). However, fetal intervention into uninjured fetal organs seeks to prevent diseases before the onset of irreversible damage. In this sense, genetically engineered, conditioned MSCs endowed with enhanced homing/engraftment capabilities (through over expression of an integrin, chemokine or cytokine) [42], should improve the intrinsic therapeutic benefit of naked MSC delivery. Additionally, the delivery of fl-MSCs from an identical developmental stage to the target fetal rabbit tissues might perhaps facilitate their incorporation into the developmental program of the transplanted fetal rabbits. This could assist in fl-MSC expansion and/or differentiation into tissue niches.
We documented persistent EGFP+-fl-MSC-derived mesodermal and endodermal tissue microchimerism using PCR, FISH, and EGFP reporter tracking by immunohistochemistry in the young EGFP+-fl-MSC-transplanted rabbits. Further work will be required to ascertain whether the allogeneic EGFP+-fl-MSCs underwent specific cell lineage differentiation, cellular fusion, or adaptation upon engraftment into the host tissues.
The developmental stage of the fetus could constitute another aspect able to influence the efficiency of MSC homing/engraftment. We have found a reverse relationship between the gestational age of the fetus at the time of intra-amniotic EGFP+-fl-MSC transplantation and the degree of long-term allogeneic EGFP+-fl-MSC engraftment. In contrast, it has been reported that intraperitoneal administration of human MSCs into sheep fetuses at E65 and E85 showed similar engraftment rates [10]. This suggests that the homing/engraftment efficiency through a given route of cell delivery might depend on the accessibility of the tissues according to the stage of development of the host [43].
On the other hand, the immunological naivety of the fetus should provide an environment facilitating allogeneic stem cell homing and engraftment. Nevertheless, the significantly different EGFP+-fl-MSC engraftment efficiencies found when comparing neonatal versus young adult transplanted rabbits or E17- versus E21-transplanted young rabbits might imply the development of host allospecific cellular and/or humoral immune responses. We have found no substantial evidence for allorecognition between splenocytes isolated from EGFP+-fl-MSC-transplanted rabbits and allogeneic fl-MSCs. In addition, no persistent immune-inflammatory infiltrates were present in any of the EGFP+-fl-MSC-positive young rabbit tissues analyzed by immunohistochemistry, although we did not perform such analysis in EGFP+ tissues from EGFP+-fl-MSC-transplanted neonates. This suggests absence of an adaptive immune barrier to engraftment after fetal allogeneic EGFP+-fl-MSC transplantation.
We recently reported the induction of humoral immunity in young rabbits after lentiviral vector-mediated fetal EGFP gene transfer [21]. In contrast, we account here lack of host-mediated production of specific antibodies against the overexpressed EGFP transgenic product. Taken together, these results suggest the induction of host tolerance to allogeneic, genetically engineered EGFP+-fl-MSCs after fetal rabbit transplantation, in line with the extensively reported immunoregulatory features of both adult and fetal MSCs [44 –46].
In summary, we have shown that the use of a safe fetal-to-fetal approach to allogeneic MSC transplantation yields low-level but persistent engraftment into different organs such as liver, kidney, and lung. Thus, the therapeutic possibilities intrinsic to the fl-MSCs may support fetal tissue regeneration or dampen chronic inflammation. Furthermore, their proper genetic engineering using safe vectors capable of yielding high-level, persistent transgene expression [28,47] may assist in the prevention or correction of a variety of inherited disorders diagnosed in the fetus such as immunodeficiencies, enzyme storage diseases, or cystic fibrosis, or support the delivery of biological agents to overcome different pathological settings. Nevertheless, further work is needed to understand the molecular mechanisms underlying MSC homing/recruitment, their in vivo growth, and/or multi-lineage differentiation potential under specific microenvironmental and developmental conditions [42,48], and the paracrine factors (chemokines, growth factors, etc.) capable of regulating their final engraftment or accommodation into relevant target tissues before a clinical application can be envisaged.
Footnotes
Acknowledgments
This work was supported by grants 01/1417 from the Fondo de Investigaciones Sanitarias (FIS-ISCIII) to E.G., from 2009SGR1490 (Generalitat de Catalunya) and from “Fundación Sira Carrasco para ayuda a la fibrosis quística” to J.M.A., and by a fellowship from “Associació Catalana de Fibrosi Quística” to I.M. J.M.A. is sponsored by the “Researchers Stabilization Program” from the SNS-Dpt. Salut Generalitat de Catalunya (Exp. CES06/012). We wish to thank Christopher Baum (University of Hamburg, Germany) for the pSFb91-EGFP vector, and Didier Trono (Geneve, Switzerland) for the pMD2.G vector encoding the pantropic VSV-G envelope glycoprotein.
Author Disclosure Statement
The authors declare no potential conflicts of interest.