
Abstract
Adult neural crest related-stem cells persist in adulthood, making them an ideal and easily accessible source of multipotent cells for potential clinical use. Recently, we reported the presence of neural crest-related stem cells within adult palatal ridges, thus raising the question of their localization in their endogenous niche. Using immunocytochemistry, reverse transcription–polymerase chain reaction, and correlative fluorescence and transmission electron microscopy, we identified myelinating Schwann cells within palatal ridges as a putative neural crest stem cell source. Palatal Schwann cells expressed nestin, p75NTR, and S100. Correlative fluorescence and transmission electron microscopy revealed the exclusive nestin expression within myelinating Schwann cells. Palatal neural crest stem cells and nestin-positive Schwann cells isolated from adult sciatic nerves were able to grow under serum-free conditions as neurospheres in presence of FGF-2 and EGF. Spheres of palatal and sciatic origin showed overlapping expression pattern of neural crest stem cell and Schwann cell markers. Expression of the pluripotency factors Sox2, Klf4, c-Myc, Oct4, the NF-κB subunits p65, p50, and the NF-κB-inhibitor IκB-β were up-regulated in conventionally cultivated sciatic nerve Schwann cells and in neurosphere cultures. Finally, neurospheres of palatal and sciatic origin were able to differentiate into ectodermal, mesodermal, and endodermal cell types emphasizing their multipotency. Taken together, we show that nestin-positive myelinating Schwann cells can be reprogrammed into multipotent adult neural crest stem cells under appropriate culture conditions.
Introduction
D
Although strong evidences exist for a glial identity of adult stem cells within the central nervous system (as reviewed in ref. [2]), this issue remained unanswered in detail for the peripheral nervous system.
Recently, we described a novel population of palatal NC-SCs (pNC-SCs) within palatal rugae (or rugae palatinae) of the palatal mucosa, which are transversal ridges developing on the secondary palate.
These nestin-positive NC-SC can be efficiently isolated, expanded, expressed Klf4, Sox2, c-myc, Oct4, and differentiated into different neural cell types [3].
Recently, it has been demonstrated that Schwann cells are highly plastic cells that are able to differentiate into melanocytes and myofibroblasts, undergo cellular reprogramming, dedifferentiate, and re-enter the cell cycle [4,5]. A hallmark of reprogramming of an adult cell to a stem cell is the alteration of its pattern in gene expression [6]. Here, the phenotype of the nestin-positive cells within the adult palate in their endogenous niche and their potential link to Schwann cells were investigated using immunohistochemistry, reverse transcription–polymerase chain reaction (PCR), correlative fluorescence, and transmission electron microscopy. Moreover, the expression pattern of cultivated neurospheres, isolated from nestin-positive Schwann cells from sciatic nerve (SN), was investigated in comparison to pNC-SCs. Finally, we analyzed the expression of the pluripotency factors Oct4, Klf4, c-Myc, and Sox2 and NF-κB protein family members within SN Schwann cells cultured in a conventional approach and as neurospheres in comparison to directly isolated SN Schwann cells and investigated the multilineage differentiation potential of Schwann cell spheres of palatal and sciatic origin as proof of their multipotency.
Materials and Methods
Preparation of cryosections
Wistar rats were killed and decapitated when older than 23 days (adult). Briefly, the whole mucoperiosteum was removed using a dental scraper, embedded in OTC compound (Miles-Bayer), and quick frozen in −40°C cold isopentane. Cryostat sections (10 μm) were cut and mounted on poly-lysine coated glass slides.
Immunohistochemistry and immunocytochemistry
Palatal section (see above) or neurospheres were harvested on microscope slides by cytospin centrifugation (40 g, 10 min; Shandon, Thermo) and fixed in phosphate-buffered 4% paraformaldehyde (pH 7.4) (4% w/v paraformaldehyde, 100 mM NaH2PO4, 0.4 mM CaCl2) for 60 min at 4°C followed by 3 wash steps in 1× PBS for 5 min. Blocking was done in 5% appropriate serum for 30 min at 23°C followed by incubation with primary antibodies for 1 h at 23°C at the following dilutions: mouse anti-nestin 1:100 (Milipore), mouse anti-β-III-tubulin 1:100 (Promega), mouse antineurofilament M 1:100 (Milipore), rabbit anti-S100 1:100 (Dako), goat anti-p75NTR 1:50 (Santa Cruz), mouse anti-Ki67 1:100 (Milipore), mouse anti-αSMA 1:50 (Sigma-Aldrich), rabbit anti-AFP 1:100 (Sigma-Aldrich), and mouse anti-Synaptophysin 1:100 (Milipore). The secondary fluorochrome-conjugated antibodies were diluted 1:300 (Alexa 555, Alexa 488 or ATTO 655, 1 h at 23°C). Nuclear counterstaining was performed with SYTOX green (1:20000; Molecular Probes). Fluorescence imaging was performed using a confocal laser scanning microscopy (LSM 510; Carl Zeiss) and analyzed using ZEN software (Carl Zeiss). The percentage of marker-positive cells was determined using ImageJ Software (National Institute of Health). Antibody controls were performed for each staining using the detection antibodies without primary antibodies to exclude unspecific fluorescence. All experiments were performed as triplicate. Representative experiments are shown.
Correlative fluorescence and transmission electron microscopy
Rats aged 3 weeks were anesthetized with Nembutal and transcardially perfused after ligation of the aorta descendens with 4% paraformaldehyde, 0.2% glutaraldehyde, and 0.5% picric acid in 0.1 M sodium phosphate buffer, pH 7.2 for 10 min. The hard palate was dissected, postfixed for additional 2 h at 4°C in 4% paraformaldehyde, infiltrated in phosphate-buffered 30% saccharose overnight, and was sectioned longitudinally into 45 μm thick sections at −22° with a cryostat. Sections were incubated free-floating in anti-nestin monoclonal antibody (Milipore) overnight at 4°, incubated in anti-mouse-biotin antibody (1.5 h), and avidin-coated quantum-dots (2 h; Qdot655; Invitrogen).
Sections in phosphate buffer were imaged using a confocal lazer scanning microscope (LSM710; Zeiss) with 633 nm excitation. Individual positive rugae were photographed, and their position was documented.
Sections were postfixed in buffered 4% glutaraldehyde (30 min), washed intensively in aqua dest, and quantum dots were silver intensified in 2 steps (each 5 min at 36°) by the method following the protocol published in ref. [7].
Sections were postfixed in buffered 0.5% osmium tetroxide (15 min, 4°C) and flat-embedded in Araldite. Selected and documented rugae were cut out with a scalpel blade, glued to the tip of a resin block, and serially sectioned into semi- and ultrathin sections. For light microscopical identification of target region, sections of 1.5 μm thickness were stained with Richardson's blue (1% w/v methylene blue, 1% w/v Azur II) for 1 min, 80°C. For electron microscopy, 60–80 nm thick sections (stained for 40 min in uranyl acetate and 7 min in lead citrate) were inspected at a Zeiss EM 109.
Cell isolation and culture of SN Schwann cells as neurospheres
All tissue was extracted from adult Wistar rats according to local guidelines (Bezirksregierung Düsseldorf). After careful removal of the connective tissue, SNs from adult rats were dissected as described by [8] with modifications. Nerves from opposite site were fixed in 4% PFA and processed for immunohistochemical staining against nestin as just described.
In particular, SNs were cut in 2 cm fragments, which were cropped with microsurgical scissors and plated on poly-L-lysine coated 100 mm cell culture dishes. After 4 h of incubation in a low volume of medium (2 mL) to allow adhesion of the nerves, the volume was increased to 10 mL. After staining and microscopical verification of the nestin expression, the initial expansion of Schwann cells was performed in DMEM containing 10% FCS until the cells reached 80% confluence. Thereafter, SNs were passaged to a fresh cell culture dish, and the adherent fraction was discarded. After 2 further expansion steps until a confluency of 80% was reached, the medium was switched to serum-free medium (DMEM/F12; Gibco) containing basic fibroblast growth factor (FGF-2; 20 ng/mL; Chemicon), epidermal growth factor (EGF; 20 ng/mL; R&D Systems), and B27 supplement. The cells were further cultivated for at least 14 days as neurosphere culture.
Cell isolation and culture of pNC-SCs
pNC-SCs were isolated and cultivated as described in ref. [3]. pNC-SCs at passage 6 were used in all experiments.
Limited dilution assay
To prove whether the neurosphere formation is due to aggregation or clonal growth, a limited dilution assay was used. After enzymatic and mechanical dissociation, the cells were diluted in cultivation media to obtain theoretically 1 cell per 100 μL and subsequently placed into 96-well flat-bottom microtiter plates. A total of 288 wells were analyzed for the presence of single cells 4 h after plating. At 96 h, the wells were examined for new-formed neurospheres using an inverse microscope equipped with phase contrast.
Reverse transcription PCR
Total RNA from cultivated pNC-SCs at passage 6 and spheres from SN were isolated using the Quiagen RNeasy Mini Kit according to manufacturer's guidelines. cDNA was synthesized in a 20 μL reaction, containing 40 U M-MuLV Reverse Transcriptase, 20U RiboLock Ribonuclease Inhibitor, 0.2 μg random hexamer primer, 5× reaction buffer, and dNTP mix. PCR was performed in a 25 μL reaction using the KAPA2G Robust PCR Kit, containing 0.5 U/0.1 μL KAPA2G Robust Polymerase (Peqlab) and 0.5 μM primers (Metabion). The cycling conditions comprised an initial denaturation of 30 sec at 95°C and 38 cycles of 5 sec 95°C, 20 sec at the appropriate temperature, and 30 sec at 72°C followed by the final elongation for 2 min at 72°C. The primer sequences are provided in Supplementary Table S1 (Supplementary Data are available online at
Multilineage differentiation of SN Schwann cells, SN spheres, and pNC-SCs
Schwann cells from SN were cultivated on poly-L-lysine coated 100 mm cell culture dishes or as neurospheres (SN spheres) in serum-free medium in the presence of FGF-2 and EGF for at least 14 days as just described. pNC-SCs were cultivated as recently described [3]. After dissociation of the spheres, or trypsinisation of Schwann cells, 1×105 cells were plated in medium containing 10% FCS for mesodermal and endodermal or 1% FCS for neuronal differentiation for at least 7 days. This cultivation method leads to spontaneous differentiation of several neural crest-derived stem cell types precultivated in serum-free medium containing growth factors [9 –11]. After 7 or 21 days of differentiation, cells were fixed and processed for immunocytochemical analysis. The percentage of marker-positive cells was determined using ImageJ Software (National Institute of Health). As a control, undifferentiated pNC-SCs and SN spheres were stained with antibodies against β-III-tubulin, αSMA, and AFP. All differentiation assays were performed as triplicate/condition. Representative experiments are shown. Statistical significance was determined by ANOVA with Bonferroni correction, using GraphPad's Prism. P≤0.05 was considered as significant.
Results
Localization of nestin-positive cells within adult rat palate
To examine the localization of neural crest-related stem cells, cryostat sections of the whole mucoperiosteum were prepared from adult rats and, subsequently, immunostained for the intermediate filament nestin.
Richardson-Blue staining showed the morphological structure of nestin-positive palatal regions (see schematic drawing in Fig. 1A and Richardson-Blue staining in Fig. 1B). In addition to axons, nerve fibres and Schwann cells were observed.

Immunoreactivity for nestin was observed in 63.2%+15.8% of the cells within the investigated structures (Fig. 1C).
Nerve fibers and synapses are present within the palatal regions expressing nestin
To verify the presence of nerve fibers and synapses adjacent to nestin-positive cells, immunohistochemical analysis of the expression of the immature neuron marker β-III-tubulin, the neuronal intermediate filament neurofilament M (NF-M), and synaptophysin was performed (Fig. 1D). Besides β-III-tubulin fibers, a branched network of neurofilament M has been detected.
Since Schwann cells occasionally have been described to express neurofilaments under certain conditions [12,13], additional staining against synaptophysin was performed.
Expression of p75NTR, S100, and Ki67
For further characterization of the endogenous niche of nestin-positive cells, immunohistochemical stainings against p75NTR, S100, and Ki67 were performed. p75NTR immunoreactivity was detected within numerous cells (Fig. 2A). Importantly, not all cells within the analyzed structures showed expression of p75NTR (52.5%+3.6%). In contrast, S100 was detected in 98.8%+0.5% of the cells. Proliferating cells, indicated by immunoreactivity for the proliferation marker Ki67, were located mainly in the epidermal layer (Fig. 2A, arrowheads) with single Ki67-positive cells in dermis (Fig. 2A, arrow).

Nestin and p75NTR are mostly coexpressed
To investigate a potential overlap in expression of nestin and p75NTR, sections were coincubated with antibodies against nestin and p75NTR. Confocal microscopy revealed a coexpression of both markers in 97.1%+1.2% of the analyzed cells (see Fig. 2B, arrows).
Nestin and S100 are coexpressed by a subpopulation of cells
Palatal sections were costained with antibodies against nestin and S100. In consistence with the results of the single stainings, nearly all cells were positive for S100 (Fig. 2C). 34.4%+4.7% of the cells coexpressed nestin and S100 (Fig. 2C, arrows), whereby 65.6% of the cells were found to be single-positive for S100 (Fig. 2C, arrowheads).
Triple staining for nestin, S100, and p75 NTR verified frequently coexpressed p75 NTR and nestin and S100 single-positive cells
The results of double labeling of frozen palatal sections with antibodies against nestin/p75NTR and nestin/S100 were verified by triple stainings for nestin, S100, and p75NTR. Confocal imaging of triple labeled sections demonstrated coexpression of nestin and p75NTR. In addition, all nestin and p75NTR double-positive cells expressed S100 (Fig. 3, arrows). In a good agreement with the results of single and double stainings, cells only positive for S100 were detected (Fig. 3, arrowheads).
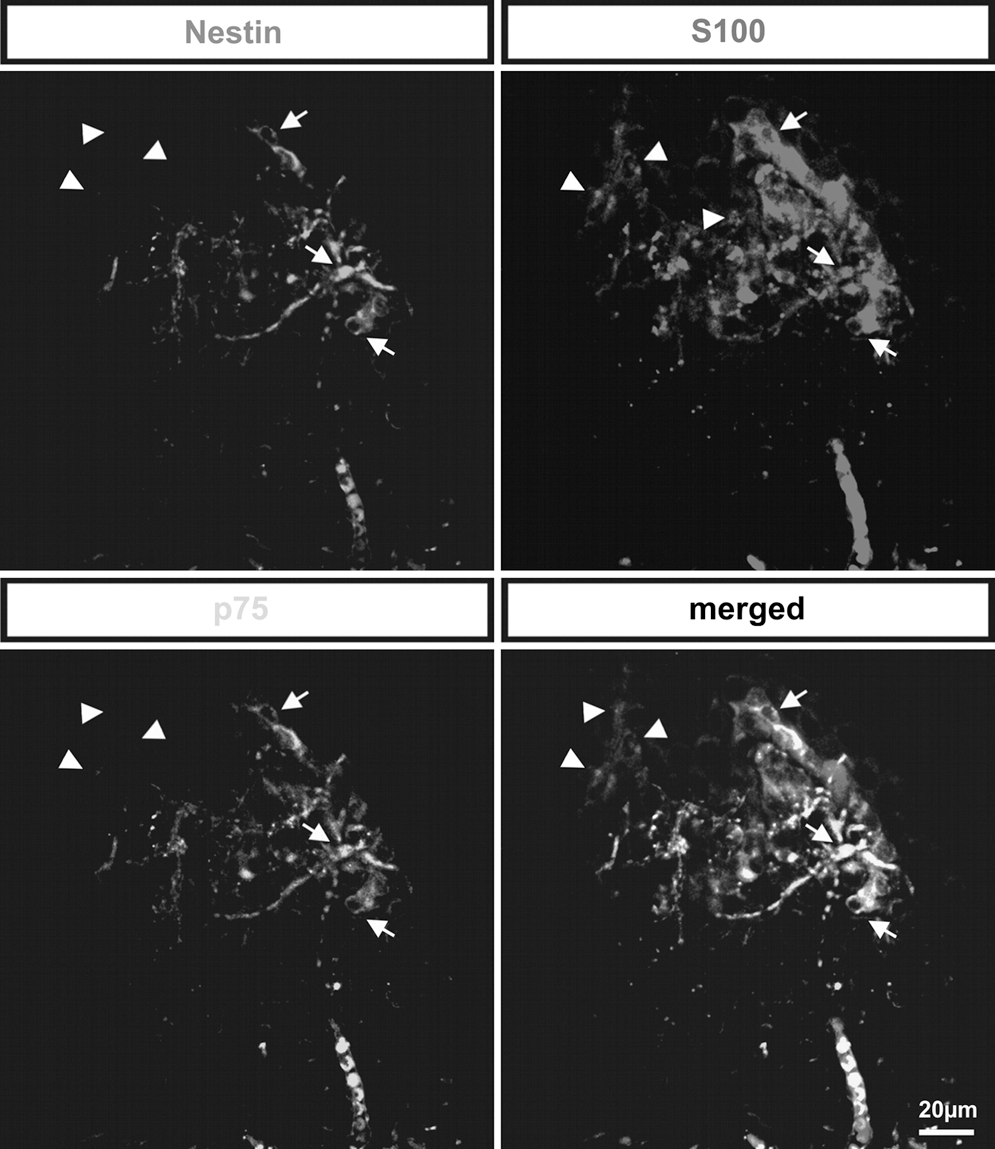
Triple stainings for nestin, S100, and p75NTR revealed coexpression of p75NTR and nestin. Triple-labeled sections indicated coexpression of nestin and p75NTR. All nestin and p75NTR double-positive cells expressed S100 (arrows). Further, several cells were positive only for S100 and not for p75NTR or nestin (arrowheads).
Correlative fluorescence and transmission electron microscopy revealed exclusive expression of nestin within myelinating Schwann cells
To investigate the expression of nestin at cellular and ultrastructural level, correlative fluorescence and transmission electron microscopy was performed on sections of perfused rats, using antibody against nestin, linker antibodies, and detection antibodies coupled to quantum dots (Fig. 4). After labeling with quantum dots, the sections were processed for confocal imaging, and nestin expression matching the distribution pattern as by labeling with conventional fluorochromes was obtained (Fig. 4A), though, in comparison, the strength of the signal was slightly decreased, most likely due to prefixation with aldehydes. These sections were further processed to electron microscopy. Sections labeled with an anti-human nestin antibody that is not able to recognize rat nestin showed no staining at all (data not shown). In contrast, a clear signal was obtained in sections labeled with anti-rat-nestin (clone Rat-401) and quantum dots (Figs. 4B and 5).
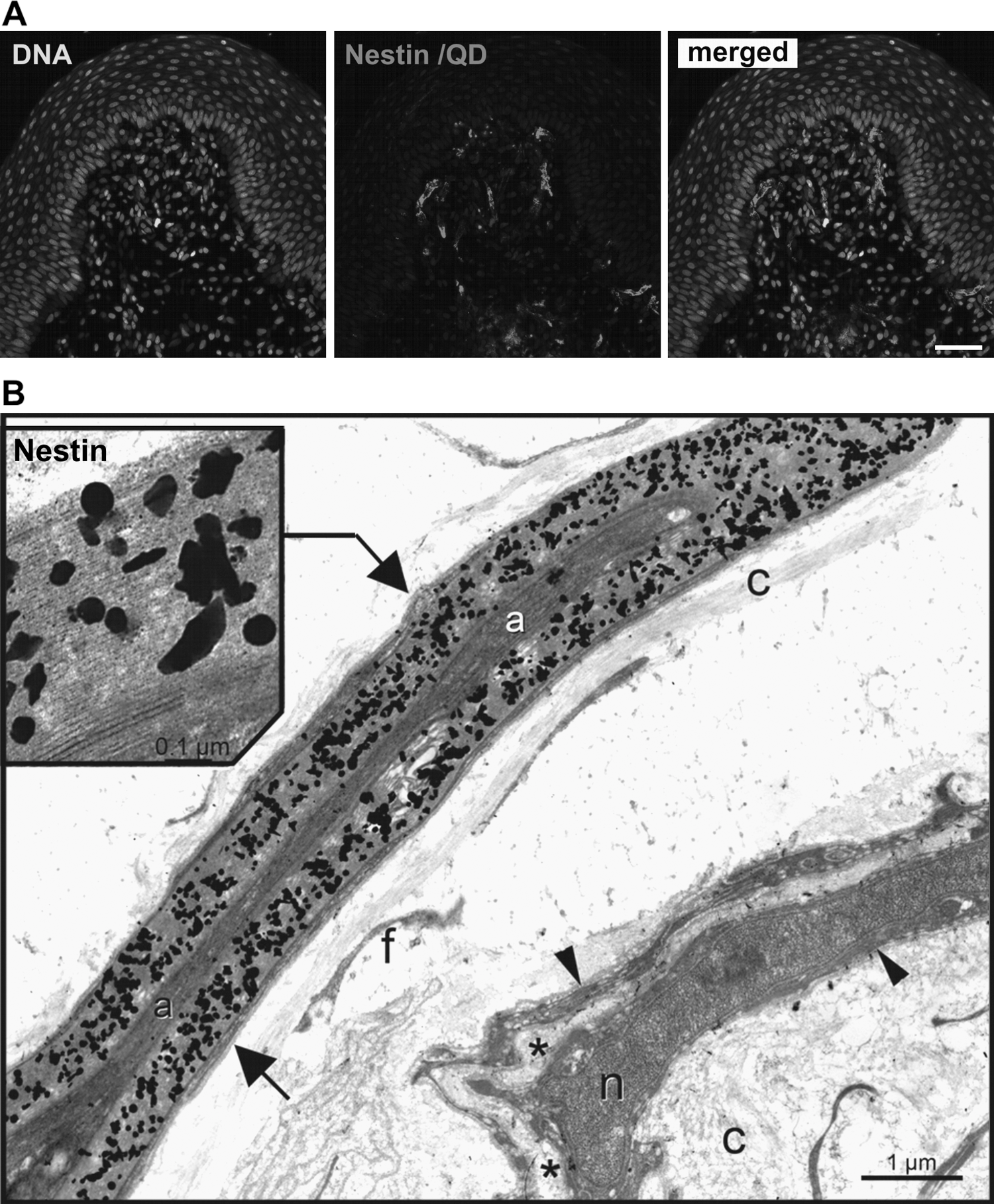
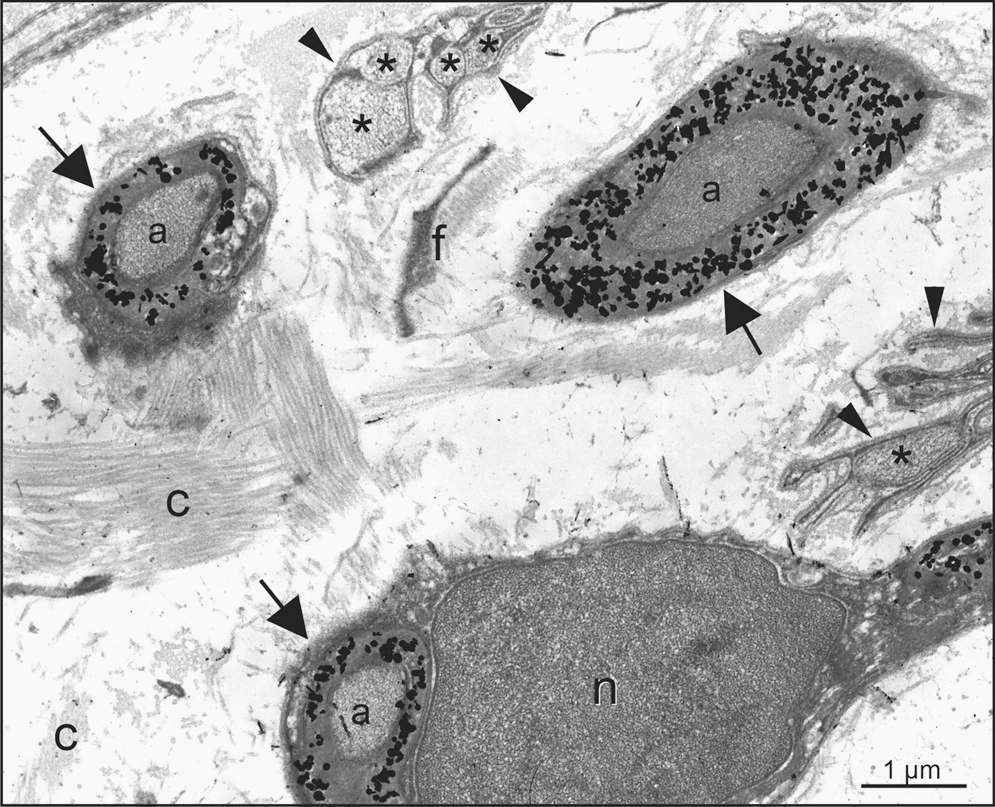
In cross-sectioned nerves of palatal rugae, nestin exclusively can be localized at the ultrastructural level (by silver-intensified quantum dots) within the myelinating regions of Schwann cells, that is, within the myelin layers (arrows) ensheathing an axon. Nestin is never found within the cytoplasmic part (arrowheads) of Schwann cells that ensheathe unmyelinated axons (asterix).
Anti-nestin immunoreactivity was localized exclusively at the ultrastructural level (by silver-intensified quantum dots) within the myelinating regions of Schwann cells, that is, within the myelin layers (Fig. 4B, arrows) ensheathing an axon. Nestin expression was not observed within the cytoplasmic part of Schwann cells that ensheathe unmyelinated axons (arrowheads). Blow-up visualized anti-nestin labeling was observed in multilayered (myelinating) membrane areas of Schwann cells ensheathing a single axon. The axon itself remained unlabeled. Schwann cell soma was not immuno-reactive to anti-nestin, as well as the Schwann cell nucleus. Other cells within the palate such as keratinocytes, fibroblasts, and endothelial cells were anti-nestin negative.
Cell isolation and culture of nestin-positive sciatic Schwann cells
To investigate the potential biological similarity between pNC-SCs, which may be closely related to cranial Schwann cells and Schwann cells from other (-trunk) anatomical regions, SNs from adult rats were dissected followed by subsequent isolation and cultivation of Schwann cells. Immunohistochemical staining of freshly dissected SN revealed expression of nestin in all cells (Fig. 6A). In contrast, SN Schwann cells were immuno-negative for p75NTR (data not shown). Such nestin-positive fibres were further dissociated und cultivated in DMEM containing 10% FCS for at least 14 days.
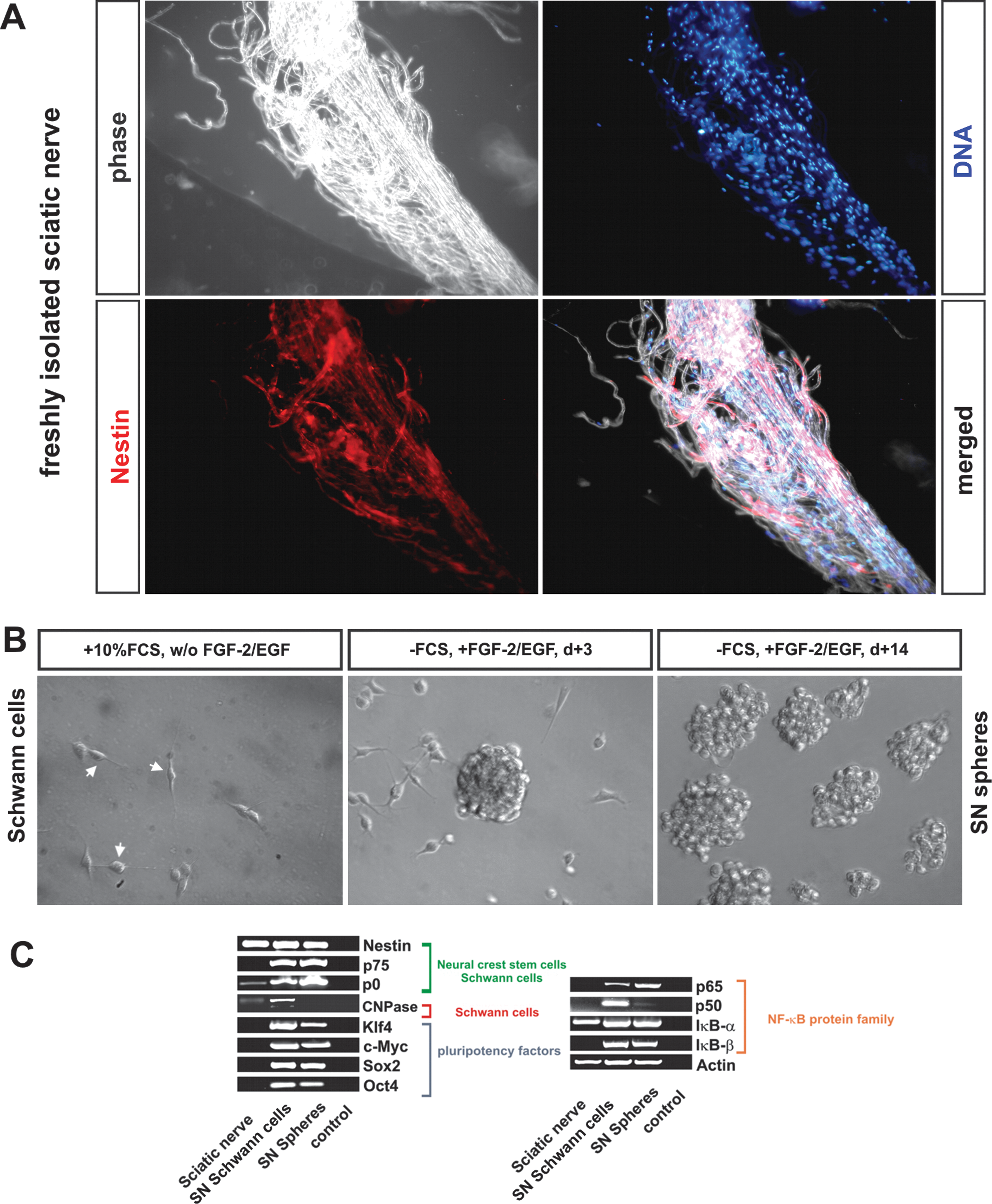
Cultivated sciatic Schwann cells can form neurospheres and induce the expression of pluripotency markers and NF-κB protein family members
Schwann cell populations of high purity can be efficiently isolated from adult SNs. Here, we analyzed whether Schwann cells from the SN could be cultivated as neurospheres. After expansion in DMEM/10% FCS and after serum deprivation and FGF-2/EGF supplementation, neurosphere formation has been observed after 3 days of serum-free cultivation (Fig. 6B). After at least 14 days, nearly all cells were in suspension and showed growth as neurospheres (Fig. 6B) and were processed for further experiments.
For the PCR analysis, freshly isolated SNs from adult rats were collected. After isolation of RNA, cDNA was generated and processed for PCR analysis. In addition, RNA from conventionally cultivated SN Schwann cells and Schwann cells cultivated as neurospheres was reversely transcribed and analyzed by PCR. SN, Schwann cells and Schwann cell spheres showed similar levels of the housekeeping gene actin and comparable expression of the intermediate filament nestin (Fig. 6C). The expression level of P0, a marker for myelinating Schwann cells and multipotent neural crest-derived cells [14], increased in cultivated Schwann cells compared with freshly isolated SN sample. If cultivated as spheres, the level of P0 was slightly elevated in comparison to conventionally cultivated cells. In contrast to cultivated Schwann cells, no message for p75NTR was detected in freshly isolated adult SNs (Fig. 6C).
Importantly, the expression of the pluripotency markers Oct4, Klf4, c-Myc, and Sox2 was exclusively observed in cultivated cells and not in freshly isolated SN. The induction of expression of these factors correlated with the induction of the expression of the NF-κB protein family members p65, p50, and IκB-β. In contrast, IκB-α was present in all samples analyzed.
SN neurospheres and pNC-SCs are able to grow in clonal manner
pNC-SC-spheres form neurospheres with similar morphology as SN derived neurospheres (Fig. 7A). To investigate the potential to grow as clonally derived neurospheres, pNC-SCs and SN spheres were dissociated and processed for a limited dilution assay. After 96 h of cultivation, sphere formation was observed in 1.85% of pNC-SCs and 3.52% of SN sphere-derived cells (Fig. 7B).
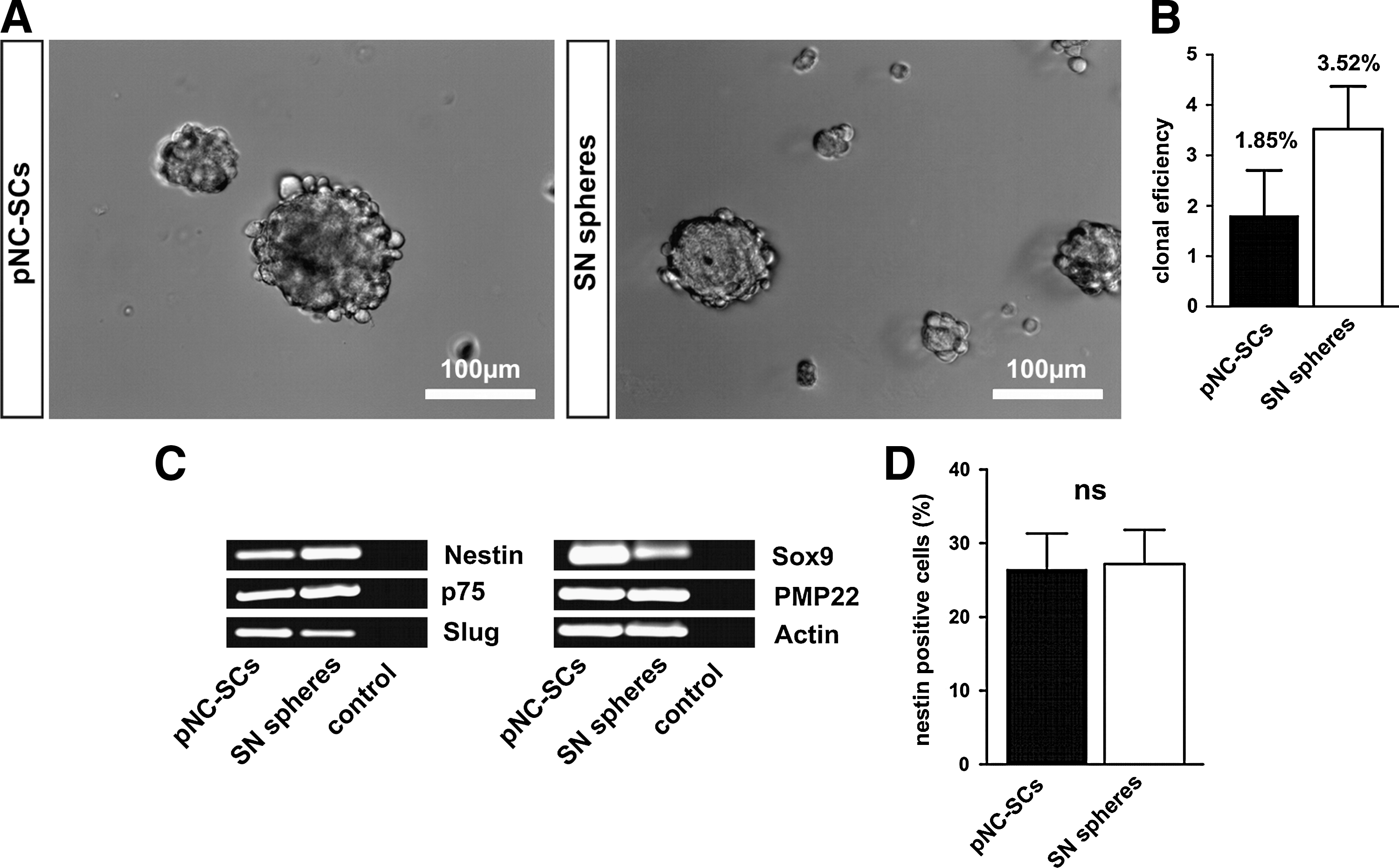
Sciatic Schwann cell neurospheres express similar levels of neural crest markers, nestin, and PMP22 as compared with pNC-SCs
pNC-SCs and Schwann cell-derived neurospheres were collected on glass slides using a cytospin centrifugation for immunocytochemistry or lysed for subsequent RNA isolation and reverse transcription-PCR analysis of the expression of neural crest and Schwann cell markers. As shown in Fig. 7C, SN Schwann cell neurospheres and pNC-SCs showed comparable levels of the neural crest/Schwann cell markers nestin, p75NTR, Slug, and Sox9. Interestingly, we detected a high level of the Schwann cell marker peripheral myelin protein 22 (PMP22) in both samples investigated. Anti-nestin staining revealed no significant differences in the percentage of nestin positive cells between pNC-SCs (26.4%+10.9%) and SN spheres (27.1%+9.3%) (Fig. 7D).
Conventionally cultivated Schwann cells, Schwann cells precultivated as neurospheres, and pNC-SCs are able to differentiate into ectodermal, mesodermal, and endodermal cells
Expression of the reprogramming factors - Oct4, Klf4, Sox2, and c-Myc - suggested that conventionally cultivated Schwann cells and/or Schwann cells and pNC-SCs cultivated as neurospheres might be multipotent.
Thus, neurospheres were dissociated as described in Materials and Methods or Schwann cells were trypsinized and processed for spontaneous differentiation assays. After immunocytochemical staining, the immuno-reactivity for specific marker proteins was detected using confocal microscopy. After 7 days of cultivation in differentiation medium pNC-SCs, conventionally precultivated Schwann cells or Schwann cells precultivated as neurospheres were fixed and stained for the lineage specific markers: β-III-tubulin for the ectoderm, αSMA for the mesoderm, and AFP for endoderm (Fig. 8). After 7 days of differentiation, 3.5%+1.8% of pNC-SCs showed expression of β-III-tubulin (Fig. 8B). Schwann cells precultivated as SN spheres and differentiated for 7 days showed no significant difference in the percentage of β-III-tubulin expressing cells compared with the pNC-SC approach (2.8%+0.5% for SN spheres vs. 3.5%+1.8% for pNC-SCs) (Fig. 8B). Importantly, Schwann cells that were not precultivated as neurospheres showed only 1.8%+0.6% of β-III-tubulin positive cells,— a significantly lower percentage compared with both pNS-CSs and the SN sphere approach (Fig. 8B).

A prolonged differentiation of SN sphere cells resulted in significantly increased percentage of β-III-tubulin expressing cells (10.1%+1.0%) (Fig. 8B). No significant differences in mesodermal and endodermal differentiation were observed between the approaches (Fig. 8B). As a control, pNC-SCs and SN spheres were stained with antibodies against β-III-tubulin, αSMA, and AFP (data not shown), thus resulting in no positive signal for the lineage markers.
Discussion
Recently, we identified neural crest-related stem cells within the palatum of rats and humans [3]. Here, we investigated the niche of NC-SCs within the rat palate. Using correlative fluorescence and transmission electron microscopy, we identified nestin-positive cells within the palate. Nestin is a well-established marker for neural stem cells [15]. Electron microscopy revealed anti-nestin immunoreactivity exclusively in myelin layers of Schwann cells but not in Schwann cells without multilayered myelin sheaths. Already in 1990, Friedman et al. showed that a monoclonal antibody raised against a nestin epitope recognizes Schwann Cells in mature and developing peripheral nerves, but this result has been mostly overlooked [16].
Cells positive for nestin, S100, and p75NTR were detected in ridges of the epidermal rugae. These structures contained β-III-tubulin-positive fibers reminiscent of Meissner corpuscles. Ultrastructural analysis by electron microscopy did not support that the structures identified here are Meissner corpuscles, as a typical MC harbors a compact corpuscular structure. Here, we did not find a corpuscular structure, but these structures are only confined to ridges of the rugae. Further, a typical Meissner corpuscle contains lamellated Schwann cells. Here, we do not find lamellated cells but nerve endings ensheathed by a single nonmyelinating Schwann cell (so called "free nerve endings"). In addition, multilayered myelinating Schwann cells, ensheathing nerve endings that were nestin-positive, were observed. Taken together, we assume that these structures resemble lanceolate nerve endings of multimodal receptors that were previously described within the rat hard palate [17].
The term "reprogramming" was used for the first time, as the conversion of glial cells into melanocytes succeeded [4]. In addition, multipotent rat neural crest stem cells can be derived from embryonic SNs [18].
In both studies, embryonic Schwann cells were employed, whereas here we report reprogramming of adult Schwann cells.
Further, fate mapping with CRE lacZ reporter mice showed a neural crest origin of Schwann cells within the embryonic SN [19]. On the other hand, differences between the cranial and trunk neural crest cells were reported (reviewed in ref. [20]). To compare our cranial NC-SCs, we developed a method for the isolation and cultivation of adult sciatic Schwann cells. Both cranial NC-SCs and cultivated SN Schwann cells showed expression of nestin and p75NTR at RNA level.
Adult sciatic Schwann cells could be easily propagated as neurospheres. Since others and we have previously reported the expression of pluripotency factors in neural crest-related stem cells [3,21 –23], we analyzed the expression of these factors in Schwann cells. In addition, we included NF-κB family members, as this transcription factor can be activated in Schwann cells via the p75NTR [24]. To our surprise, we could not detect the expression of these transcription factors and p75NTR in freshly isolated SNs. However, the cultivation either as adherent Schwann cell cultures or as neurospheres in serum-free medium strongly activated the expression of pluripotency factors, NF-κB subunits p50, p65, and the NF-κB-inhibitor IκB-β. In contrast, the mature NF-κB repressing subunit IκB-α was already expressed in freshly isolated nerves. Although the pluripotency factors were already expressed in conventionally cultivated Schwann cells and the precultivation of SN Schwann cells seems not to be mandatory for the multilineage differentiation, it largely increases the percentage of ectodermal cells in the differentiation assay (see Fig. 8). Interestingly, palatal and sciatic Schwann cells differentiated predominantly into ectodermal and mesodermal lineage as demonstrated by significantly higher number of ectodermal and mesodermal cells compared with AFP positive cells in both approaches. Both Schwann cell populations—pNC-SCs and SN Schwann cells are multipotent, as they are able to generate cells of all 3 germ layers. In contrast to multipotent cells, pluripotent cells, such as embryonic stem cells and induced pluripotent cells, can give rise in addition to ectodermal, mesodermal, and endodermal cells, to cells of the germ line.
Interestingly, palatal tissue showed a slightly different expression pattern (see Table 1) when compared with SNs, as demonstrated by the expression of p75NTR (Figs. 2 and 3) and the pluripotency factors [3] that are already found in freshly isolated palatal rugae. This result might be explained by a state of latent injury within mammalian palate caused by mechanical stress, which may lead to an increase of the frequency of multipotent neural crest stem cells expressing nestin and p75NTR. Thus, the cellular reprogramming might occur in cultivated SN Schwann cells and not necessarily in pNC-SCs that show the expression of pluripotency factors, p75 and p65 already in their niche.
Data from ref. [3].
Besides the reported glial characteristics of neural stem cells within the subventricular zone [25], it has been demonstrated for the first time by Kondo and Raff that CNS glial cells such as oligodendrocyte precursors are able to be reprogrammed to multipotent CNS stem cells which are able to differentiate into neurons and glial cells culture as neurosphere-like bodies in presence of FGF [26].
Further, it has been shown that oligodendrocytes are able to re-enter the cell cycle and that this event is directly driven by FGF-2 [27]. Sharif et al. showed that mature astrocytes can be sequentially converted into neural progenitors and stem cells which show several typical hallmarks of NSCs as formation of neurospheres, self-renewal, and proper differentiation [28]. Further, Müller cells, which represent specific glial cells of the neural retina, seem to play an important role in the retinal regeneration through re-entering the cell cycle and dedifferentiation into multipotent progenitor cells in response to FGF-2 [29,30], reviewed in ref. [31]. Importantly, these glial cells have also been demonstrated to generate nestin-positive neurospheres [32].
Recently, it has been reported that forced expression of a single neurogenic transcription factor reprogrammed astrocytes into functional neurons [33,34].
Even though cellular reprogramming of mature glial cells into adult stem cells has been described mostly within the CNS, similar mechanisms seem to exist in the peripheral nervous system. To our knowledge, this is the first evidence showing that cells derived from the peripheral nervous system can be reprogrammed to become multipotent as described for CNS glial cells.
Mechanistically, an adult cell can be reprogrammed by alteration of its gene expression [6]. Indeed, cellular reprogramming has been elegantly demonstrated by transplantation of neural crest stem cells that differentiate into bone, cartilage, and connective tissue induced by their new cellular niche [35].
Adult, cultivated somatic cells can be reprogrammed into pluripotent cells through forced expression of Oct4, Klf4, c-Myc, and Sox2 [36], the so-called pluripotency factors [6].
Until now, such cellular reprogramming has been achieved using different protocols in several cell types, for example, in multipotent adult neural stem cells using Oct4 alone [37].
However, cellular reprogramming seems not to be limited to the conversion of adult cells into pluripotency. Kohno et al. showed in their study that epithelia-like cells lacking stem cell characteristics can be efficiently converted into adult stem cells through neurosphere culture and postulated that such neurosphere formation can be a cellular reprogramming system [38]. Here, we propose a model for cellular reprogramming of adult myelinating Schwann cells.
During development, multipotent neural crest stem cells expressing nestin, p75NTR, and S100 differentiate into glial restricted progenitor cells. With further maturation, they either give rise to mature myelinating Schwann cells that down-regulate p75NTR or to p75NTR-positive nonmyelinating Schwann cells negative for nestin. Myelinating Schwann cells may undergo cellular reprogramming as a result of the supplementation with FCS or growth factors in neurosphere cultures mimicking injury leading to up-regulation of p75NTR, NF-κB subunits p50 and p65, the NF-κB-inhibitor IκB-β, and the pluripotency factors (see scheme in Fig. 9).
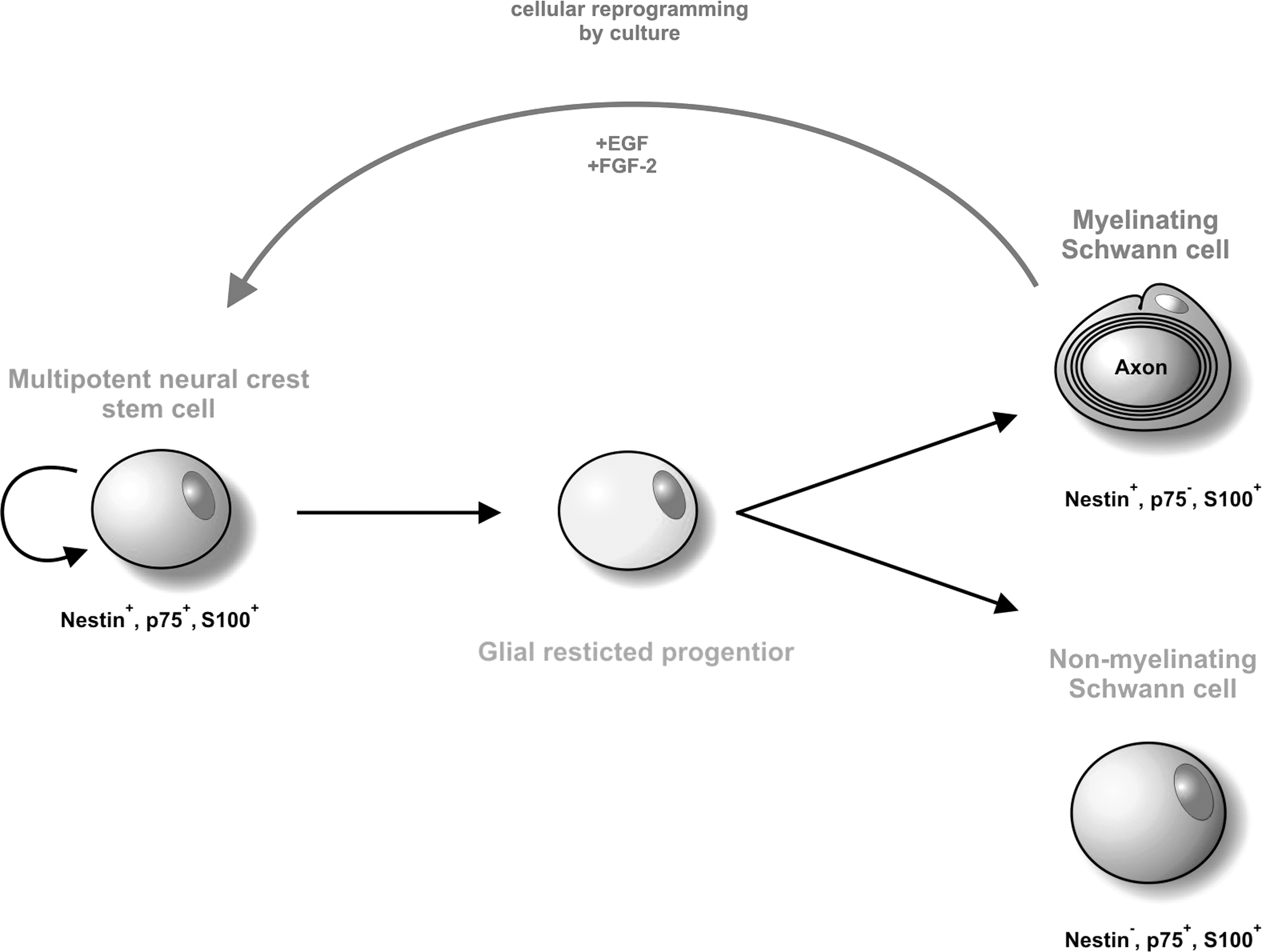
Model of the cellular reprogramming of myelinating Schwann cells into neural crest stem cells. During development, multipotent neural crest stem cells expressing nestin, p75NTR, and S100 differentiate into glial restricted progenitor cells. With further maturation, they either give rise to mature myelinating Schwann cells that down-regulate p75NTR or to p75NTR-positive nonmyelinating Schwann cells negative for nestin. During cell culture, myelinating Schwann cells can undergo cellular reprogramming and de-differentiate into multipotent nestin+, p75NTR+, and S100+ neural crest stem cells, which are able to differentiate into ectodermal, mesodermal, and endodermal cell types.
As just mentioned, in contrast to SN, such nestin/S100/p75NTR and pluripotency factors expressing cells are present within palatal rugae, which can be explained by a state of latent injury within this tissue.
The observation that adult Schwann cells could be reprogrammed into multipotent stem cells may provide an important insight into the etiology of certain cancers associated with Schwann cells [39,40]. These findings might have implications for diagnosis and treatment of Schwannomas.
Thus, a cultivation of myelinating Schwann cell in presence of FCS and as neurospheres in presence of FGF-2 and EGF may act as a cellular reprogramming mechanism into multipotency analogous to the conversion of fibroblast into pluripotency via forced expression of the pluripotency factors. The cellular reprogramming may occur via in vitro dedifferentiation of terminally differentiated Schwann cells, which thereafter are able to generate neural crest progeny in a clonal manner.
Footnotes
Acknowledgments
This study was supported by the University of Bielefed including an FiF grant of the University Bielefed to DW and grants of the German Ministry of Research and Education (BMBF) and DFG to BK and CK. MH and MH were supported by the Systems Biology Initiative (FORSYS, grant 0135262) of the German Ministry of Research and Education (BMBF). The excellent technical help of Angela Krahlemann-Köhler is gratefully acknowledged.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.