
Abstract
Human high-grade gliomas (hHGG) remain a therapeutic challenge in neuro-oncology despite current multimodality treatments. We recently demonstrated that murine embryonic stem cell (mESC)-derived astrocytes conditionally expressing proapoptotic genes can successfully be used to induce apoptosis and tumor shrinkage of hHGG tumor in vitro and in an in vivo mouse model. The first step in the translation of these results to the clinical settings, however, requires availability of human embryonic stem cells (hESC)- and/or induced pluripotent cell (hiPSC)-derived astrocytes engineered to express proapoptotic genes. The potential for directed differentiation of hESCs and hiPSCs to functional postmitotic astrocytes is not fully characterized. In this study, we show that once specified to neuro-epithelial lineage, hiPSC could be differentiated to astrocytes with a similar efficiency as hESC. However, our analyses of 2 hESC and 2 hiPSC cell lines showed some variability in differentiation potential into astrocytic lineages. Both the hESC- and hiPSC-derived astrocytes appeared to follow the functional properties of mESC-derived astrocytes, namely, migration and tropism for hHGG. This work provides evidence that hESC- and hiPSC-derived cells are able to generate functionally active astrocytes. These results demonstrate the feasibility of using iPSC-derived astrocytes, a new potential source for therapeutic use for brain tumors and other neurological diseases.
Introduction
E
Human high-grade gliomas (hHGG) remain a therapeutic challenge despite current multimodality treatments with an average survival time of 12 months for glioblastoma, the most common hHGG [11,12]. Limitation of current therapies includes their high invasiveness, which precludes effective surgical removal [13]. HGG consist of heterogeneous population of cells, some of which have been shown to extensively proliferate, self-renew, infiltrate, and be solely responsible for the growth of the main tumor mass [13]. Direct targeting of these infiltrative hHGG cells by genetically modified stem cell-derived astrocytes could be a promising therapeutic approach [14]. Our recent work supports this concept showing that cell-based gene delivery by mESC-derived highly migratory astrocytes engineered to express the proapoptotic gene TRAIL induced 20%–40% apoptosis in a panel of human glioma cells in vitro and significantly decreased tumor volume by 31% in a mouse model of human glioma xenografts in vivo [6 –10]. Astrocytes were selected as a delivery vehicle because they represent the most abundant cell type in the brain and are critical for the normal physiology of the CNS. Astrocytes are native to the CNS, which should maximize their survival and function after transplantation. They normally provide trophic and tropic support to neurons, and have important functions in protecting neurons from toxic levels of glutamate and potassium [15]. Therefore, transplanted astrocytes should be innocuous or protective within the host brain. Astrocytes are a highly secretory cell type and are able to generate large amounts of transgenic protein. Additionally, normal astrocytes have the ability to migrate along white matter tracts after transplantation into the brain [16,17]; this migratory capacity may be useful for delivery of gene therapy to infiltrative tumors like malignant gliomas. Having successfully demonstrated efficacy of mESC-derived astrocytes to treat gliomas, our next objective is to translate this work to the clinical setting. For these reasons, we first focused on establishing efficient protocols to generate iPSC-derived astrocytes.
Although methods for the directed differentiation of hESCs toward the astrocyte lineage remain scant, hESC-derived neural progenitor cells can generate astrocytes during gliogenic stages in the absence of any defined treatment [18 –20]. In the present study, we compared the astrocyte differentiation potential of human iPSCs (hiPSC) to hESCs. We found that human iPSCs, iPS-IMR90-4, and iPS-Foreskin-1 differentiate to neuro-epithelial (NE) cells and then to neurons and astrocytes with the same time course as that of hESCs, WA09 and WA01. However, their differentiation from NE cells to astrocytes had differences comparing between and among cell lines. Both the hESC- and hiPSC-derived astrocytes appeared to follow the functional properties of astrocytes, namely, migration and tropism for hHGG. This study provides detailed demonstration that hESC- and hiPSC-derived cells are able to generate functionally active astrocytes.
Materials and Methods
Cells and culture conditions
hESCs (WA-09 and WA-01; WiCell) and hiPSC lines (iPS-IMR90-4 and iPS-Foreskin-1; WiCell) were initially plated on irradiated mouse embryonic fibroblasts [MEFs, hESC shared research facility (SRF); MSSM]. Dulbecco's modified Eagle's medium (DMEM) with F12 (DMEM/F12; Cellgro) containing 20% knockout serum replacement (Invitrogen), 0.1 mM β-mercaptoethanol (Sigma), 1×nonessential amino acid (Invitrogen), and 20 ng/mL fibroblast growth factor-2 (FGF-2; R&D Systems) was changed daily. For feeder depletion, cells were passaged by mechanic dissection and enzymatic treatments [Trypsin-EDTA (0.25%); Invitrogen], and then small clusters were plated on Matrigel (BD Bioscience)-coated plate with mTeSR media with Rock inhibitor (hESC SRF) [21].
After separation from feeder cells, aggregates of human iPSCs or hESCs were cultured in low attachment plates (Costar) in the presence of knockout serum replacement to generate embryoid bodies (EBs). At day 6, EBs were plated intact onto Matrigel-coated plate in neurobasal medium containing N2 (Invitrogen) (2×), FGF-2 (10 ng/mL), and epidermal growth factor (EGF; R&D Systems) (20 ng/mL) for 3 days (day 6–9), followed by FGF+EGF+ciliary neurotrophic factor (CNTF; R&D Systems) (20 ng/mL) for an additional 3 days (day 9–12), and finally in FGF+CNTF (day 12–15) and in CNTF alone (day 15 onward). To improve differentiation toward astrocytic lineages, 3 additional conditions were tested: cardiotrophin-1 (CT-1; R&D Systems) (50 ng/mL) alone, CT-1 in combination with CNTF, and recombinant Jagged-1 (JAG1-Fc; R&D Systems) (500 ng/mL) in combination with CNTF. Neural tube-like rosettes were detached mechanically at day 15 of differentiation and cultured in poly-L-lysine, laminin (BD), or Matrigel-coated plate in the same medium. Noggin (500 ng/mL) was added to cultures for the first 12 days, and SB431542 (10 μM) was added from day 0 to 5 according to published protocols [22,23]. Recombinant Noggin and SB431542 was purchased from R&D Systems.
Immunophenotyping
Cells were fixed using 4% paraformaldehyde (Electron Microscopy Sc.) for 20 min, washed with PBS, permeabilized using 0.5% Triton X in PBS, and blocked using 1% bovine serum albumin (Invitrogen), 10% goat serum (Chemicon Intl), and/or donkey serum (Jackson Imm. Res.) in PBS. Primary antibodies used for microscopy included PAX6 (1:200; Covance), Oct4 (1:200; Biovision), Nestin (1:200; R&D Systems), Tuj1 (1:200; Covance), Nanog (1:200; R&D Systems), GFAP (1:1,000; Millipore), A2B5 (1:200; Millipore), Musashi-1 (1:200; Millipore), GLT-1 (1:500; Millipore), O4 (1:100; Millipore), Aquaporin-4 (AQ-4, 1:200; SantaCruz Biotechnology), S100 (1:200; Millipore), and GLAST (1:200; Abcam). Primary antibodies for flow cytometry included SSEA4 (R&D Systems), Oct4 (1:200), GFAP (1:200), Nestin (1:100), A2B5 (1:100), Mushashi-1 (1:100), and Tuj1 (1:100). Secondary antibodies used were Alexa 488–conjugated anti-rabbit, anti-goat, anti-mouse, or anti-guinea pig IgG and Cy3-conjugated anti-rabbit IgG. Negative controls included isotype-specific primary serum and secondary antibodies.
Reverse transcriptase PCR
Total RNA was extracted using an RNeasy kit (Qiagen). All RNA preparations were spectrophotometrically quantified before RT. The RT was performed on 1 μg of total RNA by using the Omniscript kit (Promega) and oligo-DT primers (Invitrogen) in a total volume of 20 μL. The 12.5 μL PCR mixtures each contained 0.5 μL of cDNA template. All PCR amplifications were for 40 cycles. Amplified materials were examined on 1.8% agarose gels and photographed using an Eagle Eye II imager (Stratagene).
In vitro migration assay
To assess the migratory capacity of hESC- and hiPSC-derived astrocytes, transwell experiments were performed according to published protocols [10,24,25]. For these experiments, hESC- and hiPSC-derived astrocytes were plated in the top well (5×104 cells/well) of 8.0 μm pore size transwell culture dishes (BD Bioscience). In the bottom well culture media or human malignant glioma cells (U87; 5×104 cells/well) (American Type Culture Collection) were added and incubated in DMEM+10% serum. At 24 and 48 h after plating, the migration of hESC- and hiPSC-derived astrocytes was determined by fixing the membrane, staining the cells using the Hema3 staining kit (Fisher Diagnostics), directly counting the number of migrated cells in 10 high-power fields, and calculating the mean. Experiments were done in triplicates.
Statistical analysis
Statistical difference was assessed with Student's t-test, with significance set at P<0.05.
Results
In vitro differentiation of hESC and hiPSC to NE cells
The 2 hiPSC lines used exhibited typical characteristics of hESCs, including large nucleus and multiple, prominent nucleoli, growing as colonies and expressing pluripotency markers such as Oct4 and Nanog (Fig. 1B, C). EBs generated from hiPSC and hESC were indistinguishable across lines by gross morphological analysis (Fig. 2A). The EBs were cultured for 5 days in HES media lacking FGF-2 and treated with SB43152 and Noggin and then plated at day 6 for NE differentiation. Columnar NE cells appeared at day 9 and formed typical neural tube-like rosettes at day 14 (Fig. 2B). hiPSC rosettes were morphologically indistinguishable from those differentiated from hESCs. Immunostaining showed that they were similarly positive for Nestin (Fig. 2C), a known marker of neural progenitor cells, neural stem cells, and reactive glial cells. By day 14, most of these cells became positive for Pax6, Musashi-1 [26], A2B5 (a marker for astrocyte precursors and type 2 astrocytes) [27], and Nestin (Figs. 3A and 4).

Morphology and pluripotent marker expression of human embryonic stem cells (hESCs) (WA09 line) and induced pluripotent cells (hiPSCs) (Foreskin).
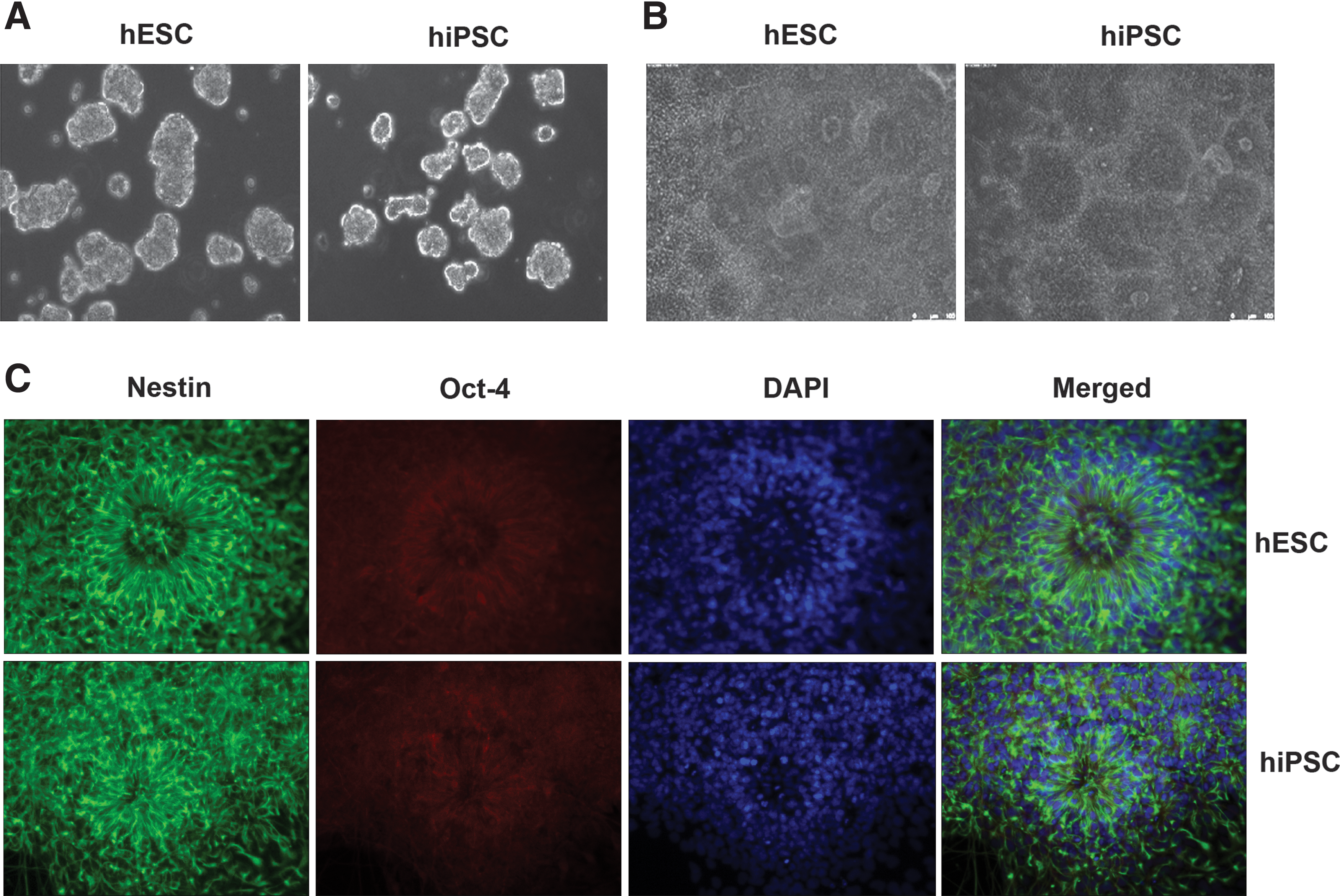
hESCs and hiPSCs can both generate EBs and neuro-epithelial (NE) cells. Phase-contrast microphotographs show

Developmental potential of NE cells into neuron and astrocytes.

Flow cytometry analysis of hESC-derived cells labeled with neural- and glial-specific markers at 2, 3, and 5 weeks of differentiation.
hESCs and hiPSCs sequentially differentiate into neurons and astrocytes
The above obtained rosettes were mechanically isolated and replated on poly-l-lysine/laminin-coated plate in CNTF-containing neurobasal media. Immunostaining at 3 weeks showed a robust expression of Tuj1 (Fig. 3B), a marker of neuronal precursor, and a fairly strong expression of GFAP+ astrocytes when the hESCs and hiPSCs were cultured in defined media. These cells also expressed Nestin at this time point. Immunostaining with O4 (oligodendrocyte precursor marker) did not show positive staining. At 5 weeks most of the hESC-derived NE cells differentiated into GFAP+ astrocytes (Fig. 3C). A few GFAP+ cells showed faint positivity for AQ-4, a marker for the main water channel in the perivascular membranes of astrocytes, Nestin, or GLAST. RT-PCR analysis (Fig. 3D) confirmed these findings. Together, these results indicate that both the hESCs and hiPSCs can be successfully differentiated into astrocytic lineage using chemically defined media.
Figure 4 summarized the expression of various cell markers during differentiation as quantified by FACS. GFAP-positive cells increased steadily during differentiation from 18% at 2 weeks to 78% at 5 weeks. Simultaneously, Oct4 and Musashi-1 (neural precursor marker) decreased from 40% and 67% at 2 weeks to 18% and 44% at 5 weeks, respectively. Similar results were seen in hiPSCs, where GFAP-positive cells were found in 70% of foreskin iPSCs and 55% of IMR90 iPSCs.
To further optimize the differentiation toward astrocytic lineage, we checked the expression of Notch receptors as previous studies showed that activated Notch enhances astrocytic differentiation in NSC from developing and adult CNS in the rodent [28 –30]. Our RT-PCR analysis results showed that hESC cells abundantly expressed Notch 1 and Notch 2 receptors, suggesting their potential for responding to Notch ligands. When cells were cultured in CNTF alone or in combination of CNTF with the Notch activator JAG1-Fc, RT-PCR analyses did not show any incremental difference in GFAP expression (data not shown). Additionally, to investigate the improved astrocytic differentiation potential we compared the effect of CT-1 [31] alone or in combination with CNTF on astrocytic differentiation. Immunostaining and flow cytometry analyses did not show any incremental difference in GFAP expression when cells were cultured in CT-1-containing neurobasal media at 3 weeks of differentiation (data not shown).
hESC and hiPSC-derived astrocytes maintain migratory capacity and tropism for hHGG
The migration pattern and “homing” of neuronal stem cell and mesenchymal stem cells in the brain has been reported [32 –34]. To confirm that hESC and hiPSC-derived astrocytes exhibit similar migratory properties, we performed an in vitro transwell migration assays. As shown in Fig. 5, both the hESC- and hiPSC-derived astrocytes showed migratory capacity and a significant tropism for hHGG cells 48 h after plating (P<0.05, Student's t-test). These results are similar to those we published for mESC-derived astrocytes [10].

In vitro transwell migration assay:
Discussion
The application of stem cell technology to create individualized therapies for patients with a variety of diseases promises to revolutionize medicine. Recent studies show that mature, differentiated cells can be reverted to a pluripotent state, thereby creating iPSC. These can then be differentiated into many lineages. In the present study, we demonstrated the successful generation of functional astrocytes from hiPSC and provide evidence that these astrocytes are phenotypically indistinguishable from hESC-derived astrocytes. Our study provides a clear characterization of astrocytic differentiation from hESC and iPSC. These data corroborate previous studies showing astrocytic differentiation from hESC in the presence of Hedgehog (Hh) inhibitor [20] and from iPSC in the absence of CNTF [35].
During embryonic development, neurons and glial cells are generated in distinct phases. A complex interaction between extrinsic environmental cues and intrinsic mechanisms regulates this developmental switch from neurogenesis to gliogenesis [36]. A number of cytokines and growth factors that are expressed in the embryonic cortex, including CT-1, neuropoietin, Notch, FGF, and EGF, have been implicated in the regulation of the neurogenic to gliogenic switch. Many of these extrinsic signals, including CNTF, leukemia inhibitory factor, and CT-1, have been shown to induce astrogliogenesis in cultured precursors via a gp130-JAK-STAT pathway. In our study, we show that a high number of hESC and hiPSC-derived NE cells express GFAP after sequential treatment with FGF-2+CNTF followed by CNTF alone as previously reported [37]. An incremental effect on astrocytic differentiation was not seen when CT-1 was used. Similarly, the addition of the Notch activator JAG1-Fc did not result in increased astrocytic differentiation. On the other hand, in the rodent brain activation of Notch pathways acted as a potent inducer of astrocytic differentiation in collaboration with the gliogenic signal CNTF [38]. In our study, in spite of abundant expression of Notch receptors seen in hESCs, addition of Notch activators to CNTF did not result in any incremental changes on astrocytic differentiation. The discrepancy of our finding with that of previous reports [28,38] could be because the differentiation potential and growth factor requirements of early precursors of murine and human stem/pluripotent cells might be different [20,39]. For example, A2B5, which characteristically labels glial precursors in rodent cells, has been reported to occasionally recognize cells of the neuronal lineage derived from hESCs [40].
In our study we also showed that iPSCs differentiate to astrocytes in a time frame similar to that of hESCs. Similarly, the iPSC-derived NEs, in response to a same set of extracellular molecules, differentiate to produce functional astrocytes. The differences in potential for astrocytic differentiation shown comparing the 2 hESC lines used remain unclear. Whereas the differentiation to NE cells is similar in nature, subsequent differentiation to astrocytes is more favorable in 1 of the 2 lines studied for each hESC and iPSC.
Our data support the possibility that reprogrammed somatic cells might prove to be a viable alternative to embryo-derived cells in regenerative medicine and cell-based therapy. Finally, as the hiPSCs appeared to follow a normal developmental progression to mature and functional astrocytes, it seems possible that disease-specific somatic cells may be reprogrammed and ultimately can be used as a potential patient-specific adjuvant cellular therapy to HGG and other neurological disorder. These results could provide a platform to further establishing patient-specific hiPSC-derived astrocyte–mediated gene therapy for hHGG. We envision to implant the hiPSC-derived astrocytes generated from hHGG patients that will carry proapoptotic gene to the same patient after surgical resection of tumor, thus eliminating the risk of immune-rejection. Thus, the use of hiPSC-derived astrocytes as delivery vehicles of therapeutic genes will be of great interest for the clinical application of stem cell-based cancer therapy.
Footnotes
Acknowledgment
The authors thank Dr. Ihor R. Lemischka for offering us to use some of the shared core facility equipments.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.