
Abstract
Bronchopulmonary dysplasia (BPD) remains a main complication of extreme prematurity. Bone marrow derived-mesenchymal stem cells (BM-MSC) prevent lung injury in an O2-induced model of BPD. The low level of lung BM-MSC engraftment suggests alternate mechanisms—beyond cell replacement—to account for their therapeutic benefit. We hypothesized that BM-MSC prevent O2-induced BPD through a paracrine-mediated mechanism and that preconditioning of BM-MSC would further enhance this paracrine effect. To this end, conditioned medium (CM) from BM-MSC (MSCcm) or preconditioned CM harvested after 24 h of BM-MSC exposure to 95% O2 (MSC-O2cm) were administrated for 21 days to newborn rats exposed to 95% O2 from birth until postnatal day (P)14. Rat pups exposed to hyperoxia had fewer and enlarged air spaces and exhibited signs of pulmonary hypertension (PH), assessed by echo-Doppler, right ventricular hypertrophy, and pulmonary artery medial wall thickness. Daily intraperitoneal administration of both CM preserved alveolar growth. MSC-O2cm exerted the most potent therapeutic benefit and also prevented PH. CM of lung fibroblasts (control cells) had no effect. MSCcm had higher antioxidant capacity than control fibroblast CM. Preconditioning did not increase the antioxidant capacity in MSC-O2cm but produced higher levels of the naturally occurring antioxidant stanniocalcin-1 in MSC-O2cm. Ex vivo preconditioning enhances the paracrine effect of BM-MSC and opens new therapeutic options for cell-based therapies. Ex vivo preconditioning may also facilitate the discovery of MSC-derived repair molecules.
Introduction
P
Perinatal lung injury during the late canalicular stage disrupts normal lung development, resulting in alveolar simplification [4]. Similar structural abnormalities have been demonstrated in a newborn rat model of BPD caused by oxygen-induced lung injury [5 –7]. Current therapies show limited efficacy. The poorly understood interactions of the multiple mechanisms that regulate normal and abnormal alveolar development drive the search for new global therapies.
Recent evidence suggests that mesenchymal stem (or stromal) cells (MSC) may represent such a global therapy. MSC hold great promise in regenerative medicine as exemplified by their therapeutic benefit in various diseases, including among others neurodegenerative, cardiovascular [8], and respiratory disorders [9 –11]. Moreover, in an oxygen-induced BPD model, we [12] and others [13,14] recently demonstrated that a tracheal injection of bone marrow (BM)-derived MSC efficiently prevented the arrest in lung development. Low rates of MSC lung engraftment suggest an alternate mechanism—beyond cell replacement—to account for the therapeutic benefit. We hypothesized that MSC prevent oxygen-induced BPD via a paracrine-mediated effect. To further explore the mechanisms and offer strategies to narrow the search for the many factors likely to be involved in this pneumo-protective effect, we further reasoned that preconditioning of MSC, by ex vivo exposure to hyperoxia, would improve the therapeutic benefit of the paracrine effect in an oxygen-induced BPD model.
Methods
Cell isolation, culture, and characterization
BM-MSC were harvested from femurs and tibiae of adult Sprague-Dawley rats as previously described [12]. Plastic adherent cells were allowed to grow to ∼80% confluency, then trypsinized and reseeded until passage 2. Fluorescence-Activated Cell Sorting analysis of cell surface markers was performed as previously described [12] according to the International Society for Cellular Therapy guidelines [15]. The differentiation potential of BM-MSC along adipogenic, osteogenic, and chondrogenic cell lineages was assessed as previously described [12]. Rat lung fibroblasts (RLF), used as control cells, were obtained from adult Sprague-Dawley rats following a published protocol [16].
Conditioned medium generation
Passage 2 BM-MSC or RLF ∼80% confluent cultures were rinsed with phosphate buffered saline and then incubated in serum-free Dulbecco's modified Eagle's medium (DMEM; Gibco Canada, Burlington, ON, Canada) for 24 h. Conditioned medium (CM; MSCcm or RLFcm) from an equal number of cells (∼1.5×106 cells/flask) was filtered through a 50 μm mesh then concentrated, ∼25-fold, using Amicon Ultra Centrifugal Filter Device (Millipore Corporation, Billerica, MA) with a 3kD molecular weight cutoff [17]. Preconditioned medium (MSC-O2cm or RLF-O2cm) was obtained by incubating for 24 h the serum-free cultured cells in a 95% oxygen incubator (Xvivo; BioSpherix, Redfield, NY).
Animal model and treatment
Sprague-Dawley (Charles River) rat pups were exposed to normoxia (21% oxygen, control group) or hyperoxia (95% oxygen, BPD model) from birth to postnatal day 14 (P14) in sealed Plexiglas chambers with continuous oxygen monitoring (BioSpherix) as previously described [18]. Litters were normalized at P0 to control for the effects of litter size and nutrition on growth. Dams were switched between normoxic and hyperoxic chambers every 48 h to prevent damage to the dam's lungs. Concentrated DMEM (as non cell-derived control) or CM (MSCcm, MSC-O2cm, RLF-O2cm) were administrated daily (1 μL/g) from P0 to P20 to rat pups intraperitoneally (i.p.). Rats were sacrificed at P21 via an injection of pentobarbital i.p. All procedures were approved by the Institutional Animal Care and Use Committee at the University of Alberta.
Echo Doppler
Pulmonary artery acceleration time (PAAT) normalized by right ventricular ejection time (RVET), a valid measure of mean pulmonary arterial pressure in rodents, was assessed with Doppler echocardiography at P15, as previously described [19].
Right ventricular hypertrophy
Right ventricular hypertrophy (RVH) was assessed by Fulton's index at P21. The right ventricle (RV) free wall was separated from the left ventricle and the septal wall (LV+S). The tissue was dried overnight then weighted the following day and the RV/LV+S ratio calculated [20].
Pulmonary arterial medial wall thickness
To assess pulmonary artery remodeling, the percent medial wall thickness (MWT) was calculated as (external diameter-lumen diameter)/vessel diameter [20].
Lung morphometry
After chest opening and ligation of right main bronchi, left lungs were inflated and fixed in situ via the trachea with a zinc formalin solution at a constant pressure of 20 cm H2O. After tracheal ligation and right lung removal, left lungs was placed in fixative overnight, then processed and paraffin embedded. Five micrometers thick coronal sections were cut and stained with hematoxylin and eosin. Alveolar structures were quantified using the mean linear intercept methods as previously described [18,21].
In vitro cell viability assay
Cell viability of BM-MSC, RLF, and rat lung epithelial cells (RLE, ATCC, Manassas, VA., ATCC # CRL-2300) exposed to normoxia and 95% O2 was evaluated by measuring the mitochondrial-dependent reduction of colorless 3-(4,5-Dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide (MTT; Invitrogen, Eugene, Oregon) to a colored blue formazan as previously described [22]. This was dissolved in dimethyl sulfoxide and the absorbance of each sample was spectrophotometrically measured at 550 nm with a Spectra Max 190 (Molecular Devices) microplate reader.
Hydroxy radical antioxidant capacity
To evaluate the antioxidant capacity of various CM, the hydroxy radical antioxidant capacity (HORAC) Assay, CM samples were prepared from RLF and MSC and HORAC evaluated using the OxiSelect™ HORAC Activity (Cell Biolabs, Inc., San Diego, CA) kit according to manufacturer's instructions. Briefly, 20 μL sample or gallic acid antioxidant standards were loaded into a 96-well plate and 140 μL fluorescein probe solution was added, followed immediately by the addition of 20 μL Fenton reagent. The wells were read every 5 min using 480 nm excitation and 530 nm emission wavelengths. The relative fluorescence at each time point was used to determine the HORAC of each sample, which was expressed as μmol gallic acid equivalents (GAE). HORAC was normalized to number of cells.
Superoxide dismutase, catalase, and glutathione reductase activity in CM were all measured directly using kits from Biovision (Mountain View, California) according to the manufacturer's instructions.
Growth factor analysis
Hepatocyte growth factor (HGF), Keratinocyte growth factor (KGF), Vascular endothelial growth factor (VEGF), and Insulin growth factor (IGF) levels were determined in CM of normoxic- and hyperoxic-exposed RLF and MSC by ELISA (R&D Systems®, Minneapolis, MN) according to the manufacturer's instructions.
Immunoblot analysis
CM was subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis in gels containing 10% acrylamide, and transferred to nitrocellulose. Membranes were blocked in 5% milk/1×Tris Buffered Saline (TBS)/0.1% Tween-20 and then immunoblotted at a 1 μg/mL dilution with rabbit polyclonal antibody to Stanniocalcin 1 (ab83065; Abcam, Cambridge, MA) in 5% bovine serum albumin/1×TBS/0.1% Tween-20 overnight at 4°C, as we previously described [22]. After extensive washing, membranes were incubated with peroxidase-conjugated goat anti-rabbit secondary antibody in 5% milk/1×TBS/0.1% Tween-20. After further washing, the intensity of the bands was normalized to the intensity of a reporter protein (actin) via chemiluminescence.
Statistical analysis
All data are expressed as mean±SEM. Intergroup differences were examined using the analysis of variance and Tukey's multiple comparison procedure as a post hoc test. A value of P<0.05 was considered statistically significant. All evaluations were done by investigators blinded to the experimental groups.
Results
CM reduces O2-induced pulmonary hypertension
Pulmonary hypertension, one of the most important comorbidities of severe BPD [23], was assessed at P15 by echocardiography (PAAT/RVET; Fig. 1A, B) and at P21 by RVH (Fig. 1C) and pulmonary arterial MWT (Fig. 2). At P15, vehicle (DMEM) treated O2-exposed rat pups showed a significant 39% reduction in PAAT/RVET compared with room air control-housed animals. MSCcm attenuated the decrease in PAAT/RVET but not significantly, while MSC-O2cm significantly improved PAAT/RVET values. At P21, all O2-exposed groups, except for the O2+MSC-O2cm group, showed significant RVH compared with the room air control group, ranging from a+94% increase in the O2+DMEM vehicle group to a +72% increase in the O2+MSCcm group. Treatment with MSC-O2cm significantly attenuated RVH. CM from O2-exposed RLF (RLF-O2cm) attenuated the decrease in PAAT/RVET and attenuated RVH, but not significantly. Pulmonary artery MWT was significantly increased in vehicle (DMEM) treated O2-exposed rat pups (Fig. 2). Both, MSCcm and MSC-O2cm significantly attenuated MWT. RLF-O2cm had no effect.
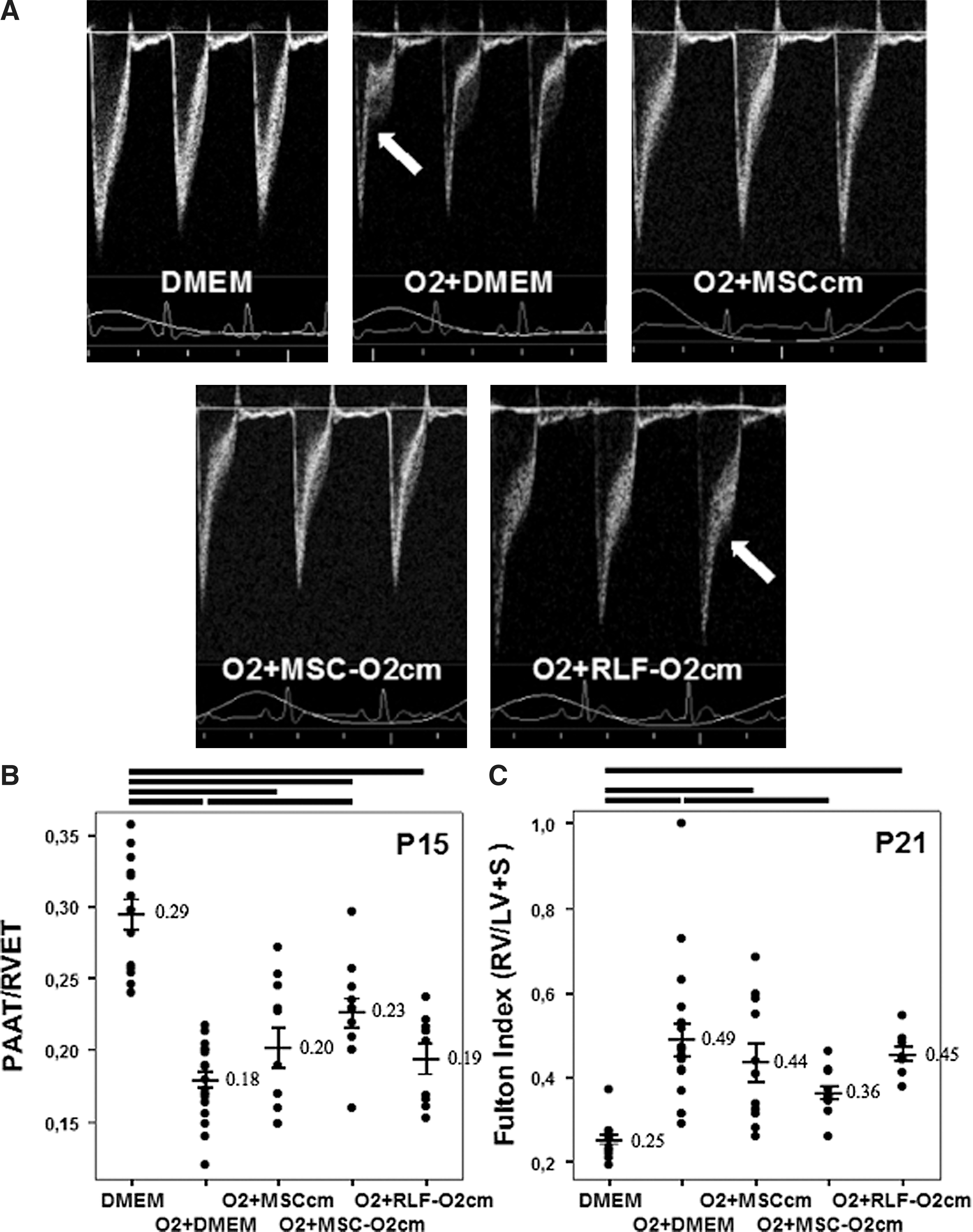
Conditioned medium (CM) harvested from O2-exposed mesenchymal stem cells (MSC; MSC-O2cm) reduces pulmonary hypertension associated with O2-induced bronchopulmonary dysplasia
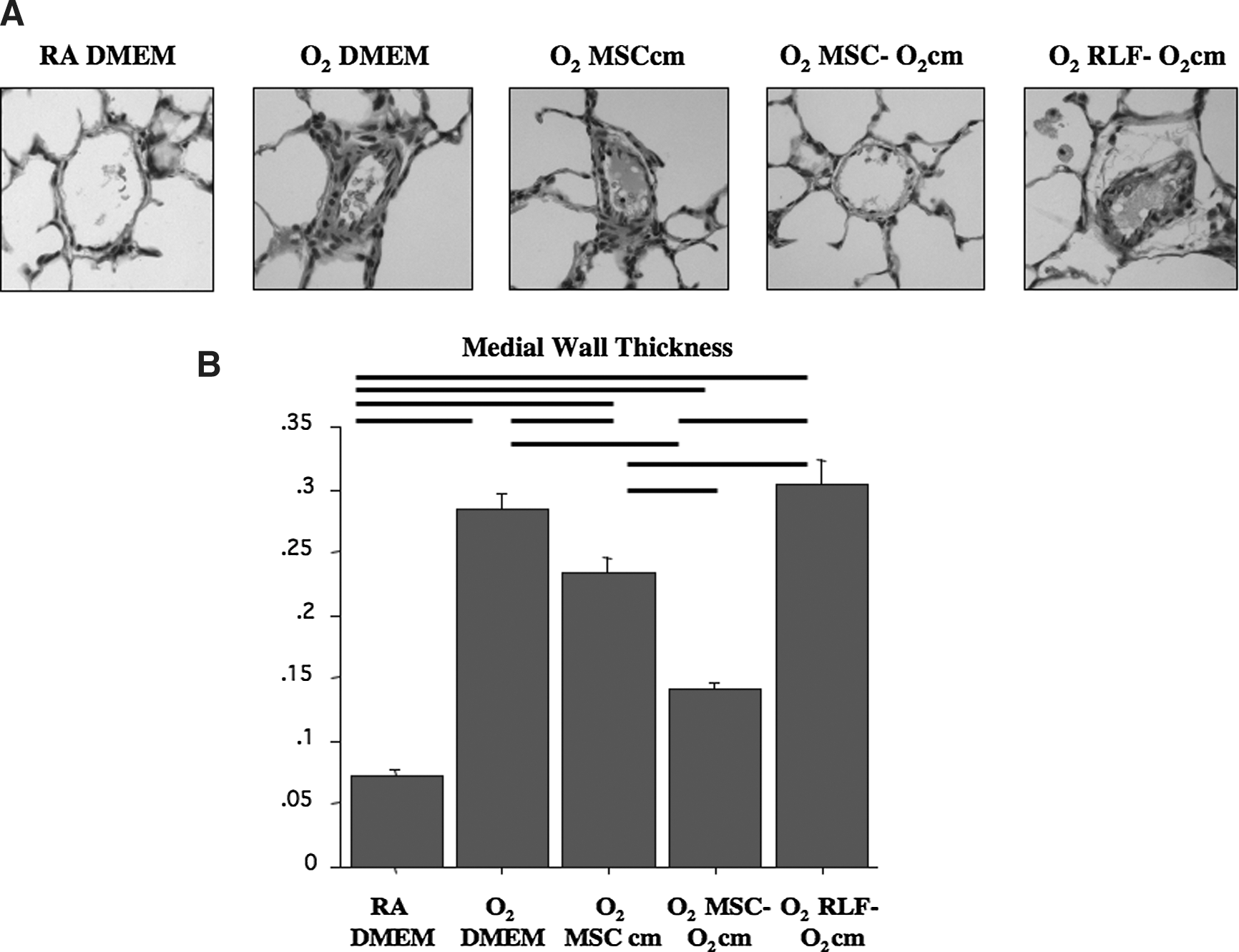
Pulmonary arterial medial wall thickness (MWT).
CM from MSC improves lung structure in O2-induced neonatal lung injury
Hyperoxic-exposed rat pups displayed the characteristic features of alveolar simplification with larger and fewer alveoli. MSCcm treatment improved lung architecture and this therapeutic benefit was further pronounced with preconditioned MSC-O2cm (Fig. 3). No effect was observed in the RLF-O2cm treated group.

CM of MSC improves lung histology and morphometry in O2-induced BPD. Sections of lung tissue from hyperoxia-exposed rats displayed the characteristic features of alveolar simplification with larger and fewer alveoli, which was not seen in control rats
Lung morphometry confirmed these observations. The mean linear intercept increased by 20% in the O2+DMEM group as compared with the room air control group. MSCcm significantly reduced the mean linear intercept by 9% as compared with O2+DMEM (Fig. 3). MSC-O2cm further reduced the mean linear intercept compared with O2+DMEM. There was no protective effect with RLF-O2cm (Fig. 3B).
BM-MSC are resistant to hyperoxia
BM-MSC, RLF, and RLE were exposed to normoxia and 95% O2 for 7 days and viability was assessed by a colorimetric MTT assay. BM-MSC, RLF, and RLE viability were the same at day 1 and 3; RLE viability was significantly higher at day 7 in normoxia compared with BM-MSC and RLF (Fig. 4A). Conversely, BM-MSC demonstrated significantly better viability in hyperoxia compared with RLF and RLE (Fig. 4B).
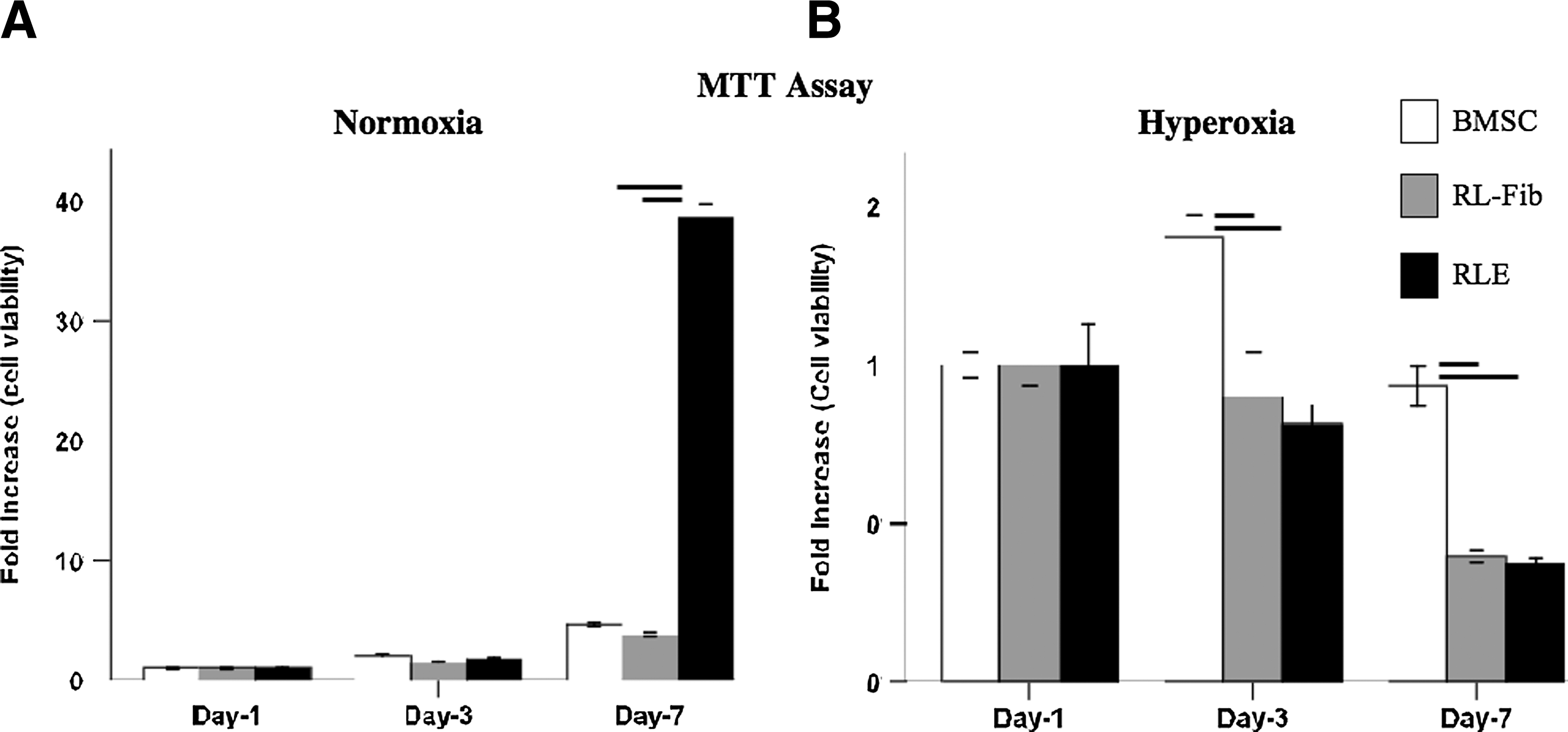
Bone marrow (BM)-MSC are resistant to hyperoxia. BM-MSC, RLF, and rat lung epithelial cells (RLE) were exposed for 7 days to normoxia and to 95% hyperoxia.
O2-preconditioning does not alter the antioxidant capacity of MSC-O2cm
To assess possible effects of ex vivo O2-preconditioning, we evaluated the HORAC of CMs harvested from RLF and MSC before or after a 24 h exposure to 95% O2 (Fig. 5). The basic HORAC level of MSCcm was significantly higher than the basic HORAC level of RLFcm and RLF-O2cm. Preconditioning did not increase the HORAC levels in MSC-O2cm compared with MSCcm. There was no difference in superoxide dismutase, catalase, and glutathione reductase activity between CM (data not shown).
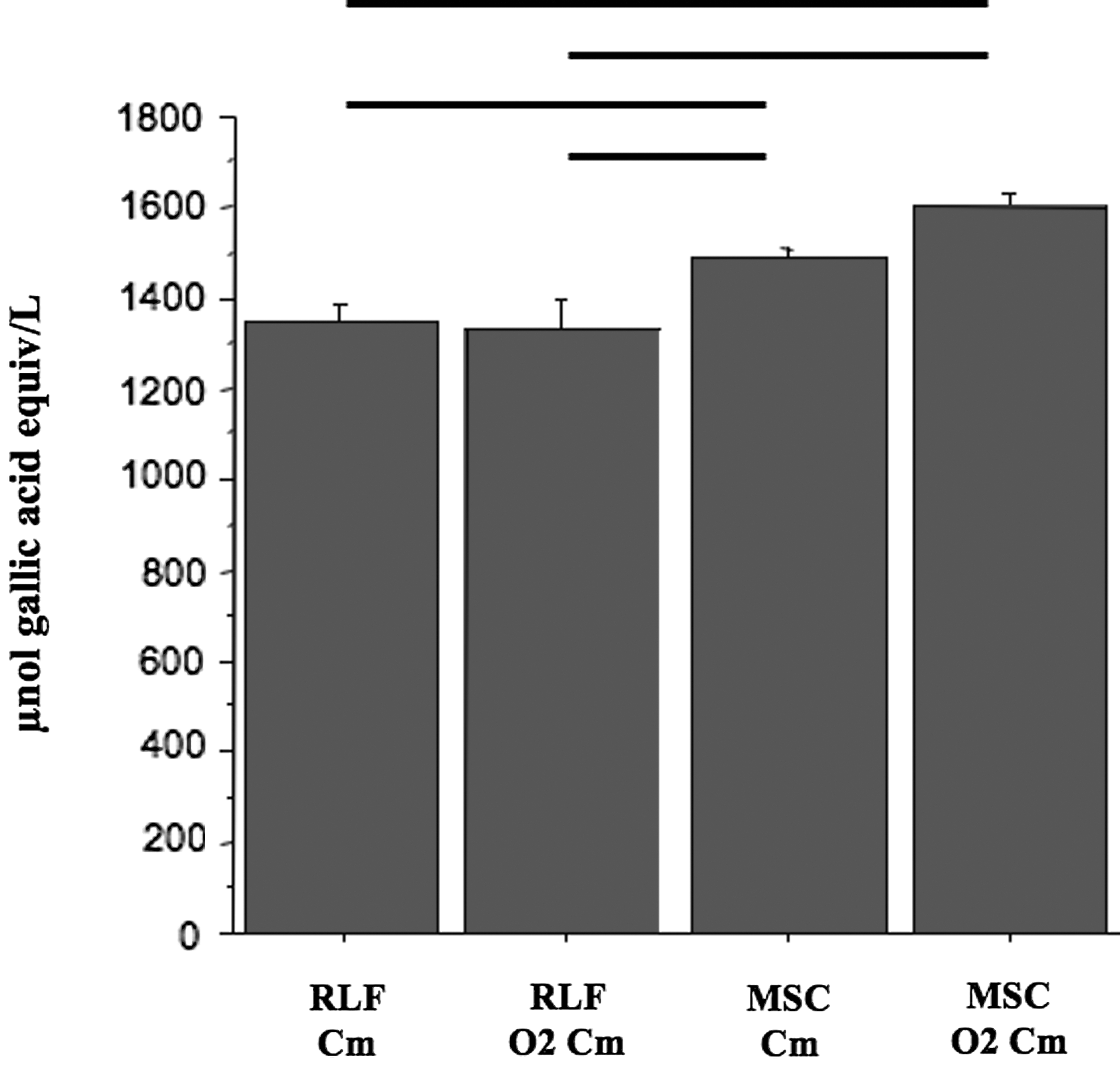
O2-preconditioning of MSC does not enhance the antioxidant capacity of the MSC-O2cm. Both CMs harvested from MSC show significantly higher antioxidant capacity as compared with CMs harvested from RLF. Preconditioning of MSC did not result in an increase in antioxidant capacity. n=6 per group. Errors bars represent the mean±SEM.—: P<0.05 between group pairs linked by this symbol.
O2-preconditioning of MSC enhances the expression of STC-1 in MSC-O2cm
To investigate putative protective factors produced by MSC, we performed immunoblot experiments for STC-1, a peptide hormone secreted by MSC with antioxidant and anti-apoptotic effects. Expression of STC-1 was not different between RLF and MSC in normoxic conditions (Fig. 6). After preconditioning in hyperoxia, Stanniocalcin-1 expression (1) significantly increased in MSC O2cm compared with normoxic MSCcm; (2) significantly increased in MSCO2cm compared with RLFcm and RLF-O2cm; (3) decreased nonsignificantly in RLF O2cm (Fig. 6). Conversely, we found no differences in HGF, KGF, VEGF, and IGF levels between CM of normoxic and preconditioned RLF and MSC (data not shown).
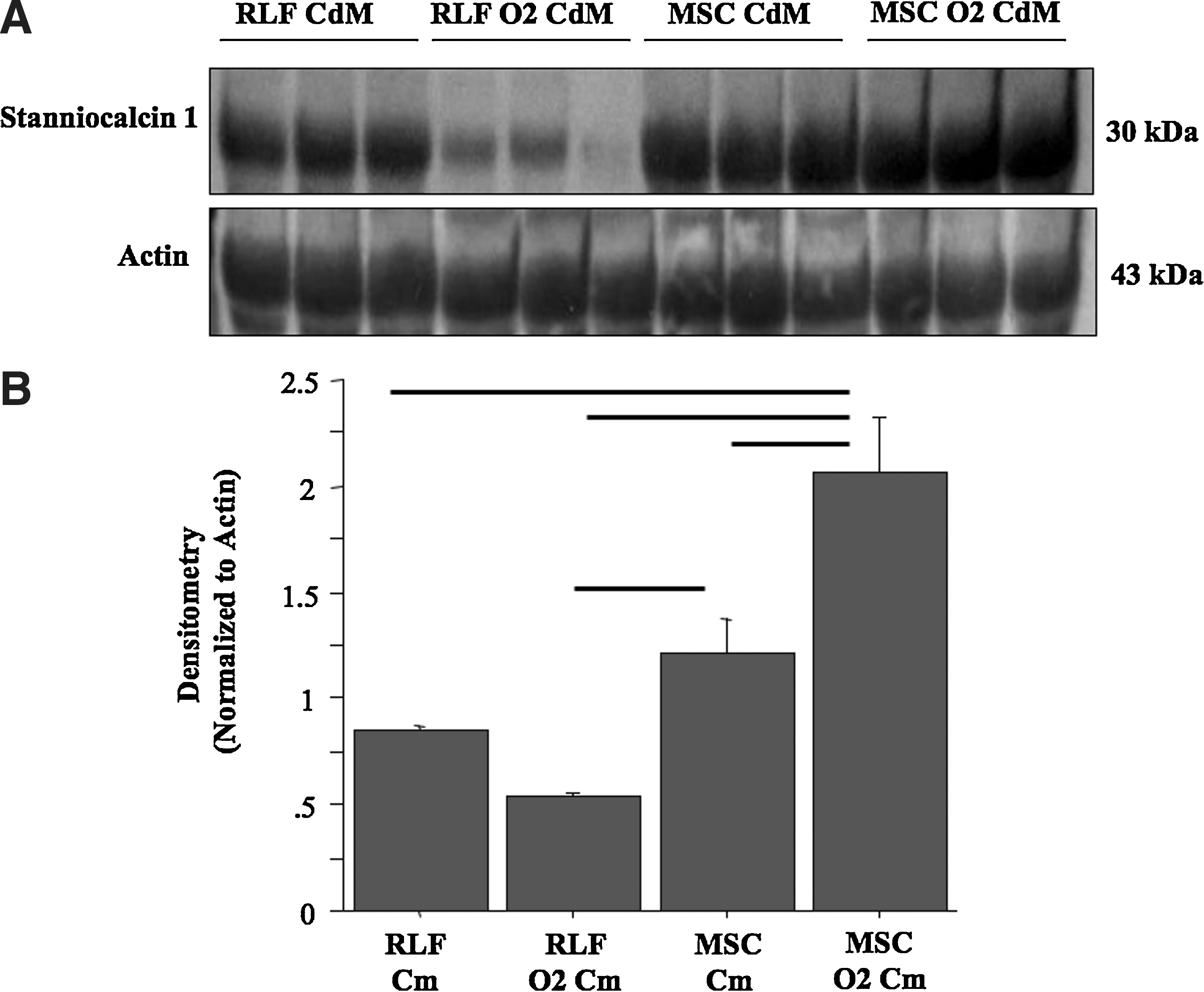
O2-preconditioning of MSC enhances the expression of STC-1.
Discussion
The aim of this study was to evaluate whether the paracrine mechanism accounts for the therapeutic benefit of BM-MSC in an oxygen-induced BPD model. Our findings demonstrated that CM by itself prevents chronic O2-induced alveolar damage in newborn rats. Further, we showed that preconditioning BM-MSC by prior ex vivo exposure to hyperoxia enhanced the beneficial effect.
Increasing evidence support the paracrine effect of MSC in various adult [9,11,24] and neonatal [12,13] lung injury models by showing a therapeutic benefit despite a low MSC engraftment rate. In vitro evidence shows that MSC-derived CM blocks inflammatory cell response [9], prevents O2-induced alveolar epithelial cell type 2 (AEC2) apoptosis, and enhances AEC2 wound healing and endothelial network formation [12]. In vivo, the importance of MSC-derived soluble factors was demonstrated in an acute liver injury model [17]. In an acute lung injury model, the paracrine effect was underlined ex vivo in perfused human lung [25]. Here, our daily i.p. administration of MSCcm, showing a partial prevention of O2-induced alveolar simplification corroborates findings by Aslam et al. in a chronic murine model of O2-induced BPD [13]. Among the recent reports of prevention of O2-induced BPD in rodents by MSC [12 –14], only one directly compared cell with CM administration, which resulted in a more profound improvement with CM alone [13]. In the current study, the discrepancy concerning the partial effect of MSCcm could be explained by numerous differences such as the tissue culture passage, the cell concentration at harvesting, the final CM concentration fold, the filter molecular weight cutoff, the species, the hyperoxia level, and the administration route. In favor of the latter hypothesis, Chang et al. showed that intratracheal administration of MSC was superior to the i.p. route in preventing O2-induced BPD [14]. Overall, our current data indicate that CM-based therapeutic approaches are promising, and that ex vivo preconditioning, amongst other strategies, could enhance the therapeutic benefit.
The minimal attenuating effect of MSCcm on PAAT/RVET and RVH contrasts with the more striking effect on the MWT. We speculate that MSCcm may have had only a mild effect on vascular compliance, possibly by changing vessel wall composition (collagen content) or the arterial wall dynamic properties (smooth muscle cell properties), which would affect the afterload, PAAT, and RVH. Thus, although the pulmonary artery structure seems improved (as reflected by the MWT), there might be only minor improvements in vascular wall function consistent with a milder effect on the PAAT and RVH.
One of the exciting findings is the enhanced therapeutic benefit obtained with MSC preconditioning. Because in a CM-based therapy, there are no more cells able to adapt to the injurious environment after administration, we hypothesized that ex vivo exposure of BM-MSC to a hyperoxic environment would precondition these cells and enhance the protective paracrine effect in this O2-induced BPD model. Our data, showing prevention of PH (Figs. 1 and 2) and enhanced preservation of alveolar growth (Fig. 3) clearly confirm this hypothesis. MSC preconditioning has been described to enhance the poor viability and the therapeutic effect of the transplanted cells in regenerative medicine and especially in myocardial research [26]. Apart from genetic modifications such as MSC overexpressing the pro-survival kinase Akt-1 [27],various preconditioning strategies—with a demonstrated in vivo benefit—have been applied, including low intensity ultrasound [28], hypoxia [29,30], anoxia [31], hyperbaric oxygenation [32], pretreatment with melatonin [33], trimetazidine [34], and LPS [35] to improve the therapeutic benefit of MSC injection. To our knowledge, this is the first demonstration of an enhanced paracrine effect with CM therapy after ex vivo preconditioning of stem cells. A paracrine strategy may alleviate the potential risk of tumor formation of whole cell therapy reported in animal [36], and in human [37] studies.
Finally, we show that while hyperoxic preconditioning of MSC did not increase the total antioxidant activity, it enhanced the release of the naturally occurring anti-oxidant STC-1. This may in part explain the enhanced therapeutic benefit of MSC-O2cm in this model. This is in accordance with recent data showing the high resistance of MSC to oxidative stress [38] and our own data confirming the enhanced viability of MSC in hyperoxia compared with lung fibroblasts and epithelial cells (Fig. 4). STC-1 was of interest for several reasons: STC-1 is a naturally occurring anti-oxidant and exerts pleiotropic effects including increased resistance of cells to damage from hypoxia, potent anti-inflammatory and renal and endothelial protective actions [39]. More interestingly, STC-1 has recently been described by Darwin Prockop's group to be secreted by MSC and to exert anti-apoptotic effects [40]. STC-1 may be one, amongst many, bioactive molecules to mediate the beneficial paracrine effect of MSC. Conversely, preconditioning did not increase the levels of several known pneumo-protective and paracrine-acting factors secreted by MSC that have been identified in other studies such as HGF, KGF, VEGF, and IGF [18,41 –44]. A more extensive screening of putative mechanisms of MSCcm holds promise to identify new healing molecules [45].
In conclusion, our study provides further evidence for the therapeutic benefit of MSC to prevent experimental neonatal lung injury and that this effect is paracrine mediated. Further, ex vivo preconditioning improves the therapeutic benefit of MSCcm. Our findings may lead to new therapeutic avenues for BPD, and also facilitate the discovery of new MSC-derived pneumo-protective factors. MSCcm-based therapy may harbor the benefit of whole cell therapy without the potential risk of tumor formation.
Footnotes
Acknowledgments
P.W. was supported by the Alberta Heritage Foundation for Medical Research (AHFMR), the Fondation pour la Recherche Médicale, and the Egide-Lavoisier Grant. B.T. is supported by the Canadian Institutes of Health Research (CIHR), AHFMR, Canada Foundation for Innovation (CFI), a Canada Research Chair, and the Stollery Children's Hospital Foundation.
Author Disclosure Statement
No competing financial interests exist.