
Abstract
Transplanted murine bone marrow (BM) progenitor cells recruit to the injured pancreas and induce endogenous beta cell proliferation to improve islet function. To enrich for analogous human progenitor cell types that stimulate islet regeneration, we purified human BM based on high-aldehyde dehydrogenase activity (ALDHhi), an enzymatic function conserved in hematopoietic, endothelial, and mesenchymal progenitor lineages. We investigated the contributions of ALDHhi mixed progenitor cells or culture-expanded, ALDH-purified multipotent stromal cell (MSC) subsets to activate endogenous programs for islet regeneration after transplantation into streptozotocin-treated NOD/SCID mice. Intravenous injection of uncultured BM ALDHhi cells improved systemic hyperglycemia and augmented insulin secretion by increasing islet size and vascularization, without increasing total islet number. Augmented proliferation within regenerated endogenous islets and associated vascular endothelium indicated the induction of islet-specific proliferative and pro-angiogenic programs. Although cultured MSC from independent human BM samples showed variable capacity to improve islet function, and prolonged expansion diminished hyperglycemic recovery, transplantation of ALDH-purified regenerative MSC reduced hyperglycemia and augmented total beta cell mass by stimulating the formation of small beta cell clusters associated with the ductal epithelium, without evidence of increased islet vascularization or Ngn3+ endocrine precursor activation. Thus, endogenous islet recovery after progenitor cell transplantation can occur via distinct regenerative mechanisms modulated by subtypes of progenitor cells administered. Further, understanding of how these islet regenerative and pro-angiogenic programs are activated by specific progenitor subsets may provide new approaches for combination cellular therapies to combat diabetes.
Introduction
T
Our previous studies have established that transplantation of murine bone marrow (BM)-derived progenitor cells augmented glycemic control in streptozotocin (STZ)-treated mice by increasing insulin production and proliferation of recipient-derived beta cells [7]. Further, islet recovery can be induced by hematopoietic chimerism in overtly diabetic NOD mice [8], and simultaneous infusion of murine BM cells with allogeneic multipotent stromal cells (MSC) can optimize islet repair and protect against T-cell-mediated beta cell deletion [9]. Recently, high-dose intercardiac infusion of human BM-derived MSC into STZ-treated NOD/SCID mice has shown the ability to regenerate islets [10]. Thus, human BM represents a realistic source of expandable progenitor cells for the development of regeneration-inducing therapies for diabetes.
The mechanisms whereby the postnatal endocrine pancreas can undergo repair or regeneration after injury remain controversial. Lineage-tracing studies by Dor et al. have established that after partial pancreatectomy, there is an increase in mature beta cell replication, and this represents the principle mechanism responsible for restoring beta cell number [11]. In contrast, other groups have suggested that beta cells can also be regenerated via neogenesis, the budding of new islets from the ductal epithelia after partial pancreatectomy [12], or ductal ligation [13]. Although the existence and identity of a pancreas-derived endocrine precursor have been debated, Seaberg and colleagues have clonally isolated multipotent pancreatic precursor cells from mouse islets and ductal regions [14], and Ngn3+ endocrine precursors associated with the ductal lining have also been identified [15,16]. Despite the controversy, the regenerative programs activated during islet recovery induced by human progenitor transplantation are largely uncharacterized and remain the key to understanding endogenous beta cell regeneration.
To identify human progenitor lineages that augment beta cell regeneration after transplantation, and to further investigate the endogenous regenerative mechanisms involved, we purified human BM cells using a conserved progenitor function, high-aldehyde dehydrogenase (ALDHhi) activity [17]. ALDH is a cytosolic enzyme involved in the production of retinoic acid and in progenitor cell self-protection from oxidative stress. Fluorescence-activated cell sorting (FACS) of human BM ALDHlo or ALDHhi cells isolates heterogeneous populations with low- or high-clonal progenitor activities, respectively [18,19]. Thus, ALDH purification represents a clinically applicable method for the simultaneous enrichment of progenitor cells from hematopoietic [20,21], endothelial [19], and mesenchymal lineages [18,19]. We have previously shown that after hematopoietic reconstitution, the progeny of transplanted human umbilical cord blood (UCB)-derived ALDHhi cells traffic efficiently to the ductal and peri-islet regions of the pancreas [22], and transplanted human BM ALDHhi progenitor cells promote revascularization in a mouse model of critical limb ischemia [18]. We hypothesized that transplantation of ALDHhi progenitor cells, or ALDH purified, ex vivo cultured MSC will aid in the growth [7] and vascularization [23] of islets during regeneration in STZ-treated NOD/SCID recipients.
Materials and Methods
Progenitor cell isolation and propagation
Human BM was obtained with informed consent from the London Health Sciences Centre, London, Ontario. Mononuclear cells (MNC) were isolated by Hypaque-Ficoll centrifugation (Pharmacia), followed by erythrocyte lysis. Cells were incubated with Aldefluor™ reagent (StemCell Technologies) and selected by FACS (FACS Aria; BD Biosciences) for low-side scatter and low- or high-ALDH activity at the London Regional Flow Cytometry Facility as previously described [18,20 –22]. BM-derived ALDHlo and ALDHhi cells were plated at limiting dilution in AmnioMax media+supplement for the expansion of MSC grown on plastic as previously described [18].
Transplantation into hyperglycemic NOD/SCID mice
Immunodeficient NOD/SCID mice (Jackson Labs) were intraperitoneally-injected with 35 mg/kg/day STZ (Sigma-Aldrich) for 5 days as previously described [7]. On day 10, hyperglycemic mice (15–25 mM) were sublethally irradiated (300 cGy) and transplanted by tail vein injection with phosphate-buffered saline (PBS), uncultured ALDHlo (2×106) or ALDHhi cells (2×105), or culture-expanded BM-derived ALDHlo or ALDHhi MSC (5×105). Systemic blood glucose concentrations were monitored weekly for 42 days using a glucose monitor (Abbott Diabetes Inc.). About 24 h before sacrifice, mice were injected with 200 μg of EdU (Invitrogen) to label dividing cells. After cervical dislocation, blood was collected from the left ventricle for serum insulin quantification by mouse insulin ultra-sensitive ELISA (ALPCO). The pancreas and BM were collected for analysis of human cells by FACS (FACS Caliber; BD Biosciences) as previously described [22].
Quantification of islet size, number, and beta cell mass
The splenic portion of the pancreas was frozen in optimal cutting temperature media (Tissue Tek; Sakura-Finetek) and sectioned at 10 μm such that each slide contained 3 sections, each 150 μm apart. Sections were fixed in 10% buffered formalin (Sigma) and blocked with mouse-on-mouse reagent and peroxidase (Vector Laboratories). Mouse antimouse insulin antibody (Sigma) diluted 1/1,000 was detected with peroxidase-labeled rabbit antimouse antibody and DAB (Vector), counterstained with hematoxylin. Size and number of islets were quantified by light microscopy (Axioscope Z2; Zeiss) analyzing 4 areas (1 mm2) selected randomly per section from a total of 3 sections/mouse and analyzed using Northern Eclipse software. Beta cell mass was calculated by beta cell area/(total area–beta cell area) X pancreas weight.
Quantification of islet blood vessel density
Rabbit antimouse von Willebrand factor (vWF) antibody (1/200; Chemicon) was detected with fluorescein-labeled goat antirabbit IgG secondary antibody (diluted 1/200, Vector), costained with mouse antimouse insulin antibody (1/1,000; Sigma) detected with Texas red-labeled rabbit antimouse antibody (1/200; Vector), and mounted in VectaShield with DAPI (Vector). Intra- and extra-islet blood vessel densities were quantified by counting vWF+ vessels/islet by randomly selecting 4 islets/section for 3 sections/mouse and by counting vWF+ vessels/section (not including islet-bound) for 3 sections/mouse.
Quantification of islet and duct association
Rabbit antimouse ck19 (1/200; Abcam Inc.) was detected with fluorescein-labeled goat antirabbit antibody (1/200; Vector) and costained with antimouse insulin as just described. Each islet in 3 pancreatic sections per mouse was designated as not associated with, or in direct contact with, ck19+ ducts.
Analysis of proliferating cells
EdU+ cells were detected using the Click-iT™ EdU imaging kit (Invitrogen) and costained for antimouse insulin or vWF as just described, to identify islet- or vessel-derived cell proliferation, respectively.
Analysis of Ngn3 expression
Goat antimouse Ngn3 antibody (1/200; Santa Cruz Inc.) was detected with fluorescein-labeled rabbit antigoat antibody (1/5,000; Vector) and costained with antimouse insulin as just described. Embryonic day 18 mouse pancreas sections were used as positive controls, and ALDHlo or ALDHhi MSC transplanted mice pancreata were stained for Ngn3-expression at day 10, 14, 17, or 42.
Statistics
Data were expressed as mean±standard error of the mean. Analysis of significance was performed by two-way ANOVA for blood glucose data, and by one-way ANOVA followed by Tukey's posthoc test for all other analyses.
Results
Transplantation of human ALDHhi cells improved systemic hyperglycemia
We FACS-purified human BM based on low-side scatter and low versus high Aldefluor™ fluorescence (Fig. 1a). ALDHhi cells were rare in human BM (0.8%±0.2%), whereas cells with 5-fold lower ALDH activity (ALDHlo) represented 8.2%±1.3% of BM cells, respectively. We have previously shown that UCB- [20] and BM-derived [18] ALDHhi cells demonstrated enriched expression of progenitor cell surface markers (CD34, CD133) and that BM-derived ALDHhi cells were enriched for hematopoietic, endothelial, and multipotent mesenchymal-stromal CFC in vitro [18].
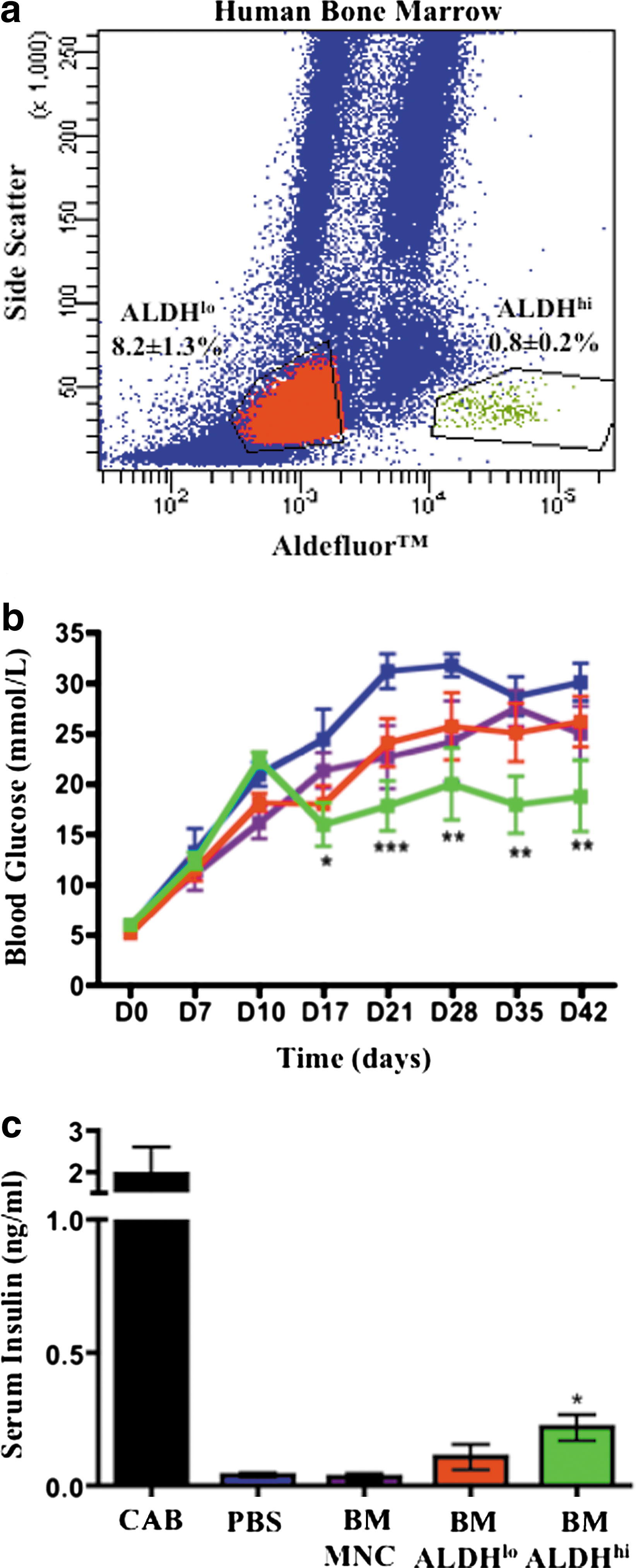
Transplantation of human BM-derived ALDHhi cells improved blood glucose and increased serum insulin in STZ-treated NOD/SCID mice.
Hyperglycemic (15–25 mM at day 10) NOD/SCID mice were intravenously injected with human ALDHlo cells (2×106), ALDHhi cells (2×105), or unpurified MNC (20×106) on day 10. Cell doses were chosen according to the availability of sorted cell populations and the relative frequencies of progenitor CFC in vitro. Compared with PBS-injected controls that remained severely hyperglycemic (>25 mM), transplantation of 2×106 BM-derived ALDHlo cells led to temporary improvement in glycemia at day 17, with a return of severe hyperglycemia from days 21 to 42 (Fig. 1b). In contrast, transplantation of 10-fold fewer (2×105) BM ALDHhi cells led to improved systemic glycemia from days 17 to 42 (Fig. 1b). Although blood glucose was stabilized after the transplantation of ALDHhi cells, the plateau in glycemia (16.0±2.1–20.1±3.6 mM) was not significantly reduced compared with day 10 (22.5±1.3 mM), indicating that transplanted ALDHhi cells could improve but not reverse established hyperglycemia. Interestingly, mice transplanted with unpurified MNC (20×106) did not exhibit improved glycemia (Fig. 1b).
At day 42, STZ-injected mice showed >10-fold reduced serum insulin concentrations compared with citric acid buffer (CAB)-injected vehicle controls (P<0.01, Fig. 1c). However, after transplantation, only BM-derived ALDHhi cell transplanted mice had significantly increased serum insulin compared with PBS controls (Fig. 1c), corresponding to the observed decrease in blood glucose in this cohort, indicating that modest stimulation of insulin production could impact hyperglycemia in our model.
Transplanted ALDHhi cells engrafted the mouse pancreas
To promote survival of transplanted human cells [20], recipient mice were sub-lethally irradiated (300cGy) before transplantation, and human cell engraftment in the mouse BM and pancreas was quantified by flow cytometry at day 42 (Fig. 2a). Mice transplanted with MNC or ALDHhi cells showed consistent human hematopoietic reconstitution (Fig. 2b). In contrast, transplantation of 2×106 ALDHlo cells did not significantly engraft recipient BM (Fig. 2b). Transplanted ALDHhi cells from human BM also showed consistent but low frequency (<0.4%) human cell engraftment in the murine pancreas (Fig. 2c), whereas transplanted human MNC or ALDHlo cells were not detected in the pancreas. As described by our previous studies [7,22], ALDHhi cells engrafted in ductal regions and surrounding murine islets did not demonstrate insulin expression (Fig. 2d).

Engraftment of human BM-derived ALDHhi progenitor cells in the BM and pancreas STZ-treated NOD/SCID mice.
To address the kinetics of human cell recruitment to the injured pancreas, mice transplanted with ALDHlo (2×106) or ALDHhi (2×105) cells at day 10 were analyzed for human cells in the pancreas at day 14, 17, or 24. At day 14, human cells were not detected in the pancreas of mice transplanted with either ALDHlo (n=3) or ALDHhi cells (n=3). At days 17–24, human cells were detected at low frequency (0.1%–0.2%) in 5 of 6 mice transplanted with ALDHhi cells compared with only 1 of 8 mice transplanted with ALDHlo cells. Nonetheless, human ALDHhi cell were detected in the pancreas at a time that correlated with the induction of improved systemic glycemia.
Transplantation of cultured MSC from independent human BM samples showed variable capacity to reduce hyperglycemia
Since human BM-derived ALDHlo and ALDHhi populations contained cells with mesenchymal-stromal function previously associated with beta cell repair [10], we sought to investigate the capacity of ex vivo expanded ALDHlo or ALDHhi MSC to improve hyperglycemia after transplantation. ALDHlo or ALDHhi MSC from 6 independent BM samples were expanded for 3–5 days and tail vein injected at 5×105 MSC on day 10 (Table 1). Independent of whether MSC were derived from ALDHlo or ALDHhi cells, mice transplanted with MSC from 3 independent BM samples showed consistently reduced hyperglycemia, whereas ALDHlo or ALDHhi MSC from 3 alternative BM samples showed little capacity to reduce hyperglycemia (Table 1). Thus, ex vivo expanded MSC showed donor-dependent variability in their capacity to improve glycemia after transplantation.
ALDHlo or ALDHhi MSC established from 6 independent BM samples were expanded for 3–5 days in ex vivo culture and transplanted at day 10 into hyperglycemic NOD/SCID mice as a single dose of 5×105 MSC. Compared with time-matched PBS controls, transplantation of either ALDHlo or ALDHhi MSC from 3 independent BM samples consistently improved systemic glycemia. In contrast, transplantation of ALDHlo or ALDHhi MSC from 3 additional human BM samples did not improve systemic glycemia. Data represent mean±SEM.
P<0.05 compared with PBS controls.
ALDH, aldehyde dehydrogenase-activity; BM, bone marrow; PBS, phosphate-buffered saline; SEM, standard error of the mean; MSC, multipotent stromal cell.
We selected BM18 and BM20 to represent nonregenerating and regenerating samples for further analysis. Although BM18 MSC did not reduce hyperglycemia when compared with PBS controls, mice transplanted with BM20 ALDHlo or ALDHhi MSC showed improved glycemia (Fig. 3a) and increased serum insulin (Fig. 3b). Glucose levels stabilized in the moderately hyperglycemic range (13.9±2.7–17.8±3.0 mM) similar to day 10 concentrations (Fig. 3a). Similar to findings by Lee et al. [10], improved glycemia was independent of long-term engraftment in the pancreas where human cells were not detected by FACS at day 17 or 42.
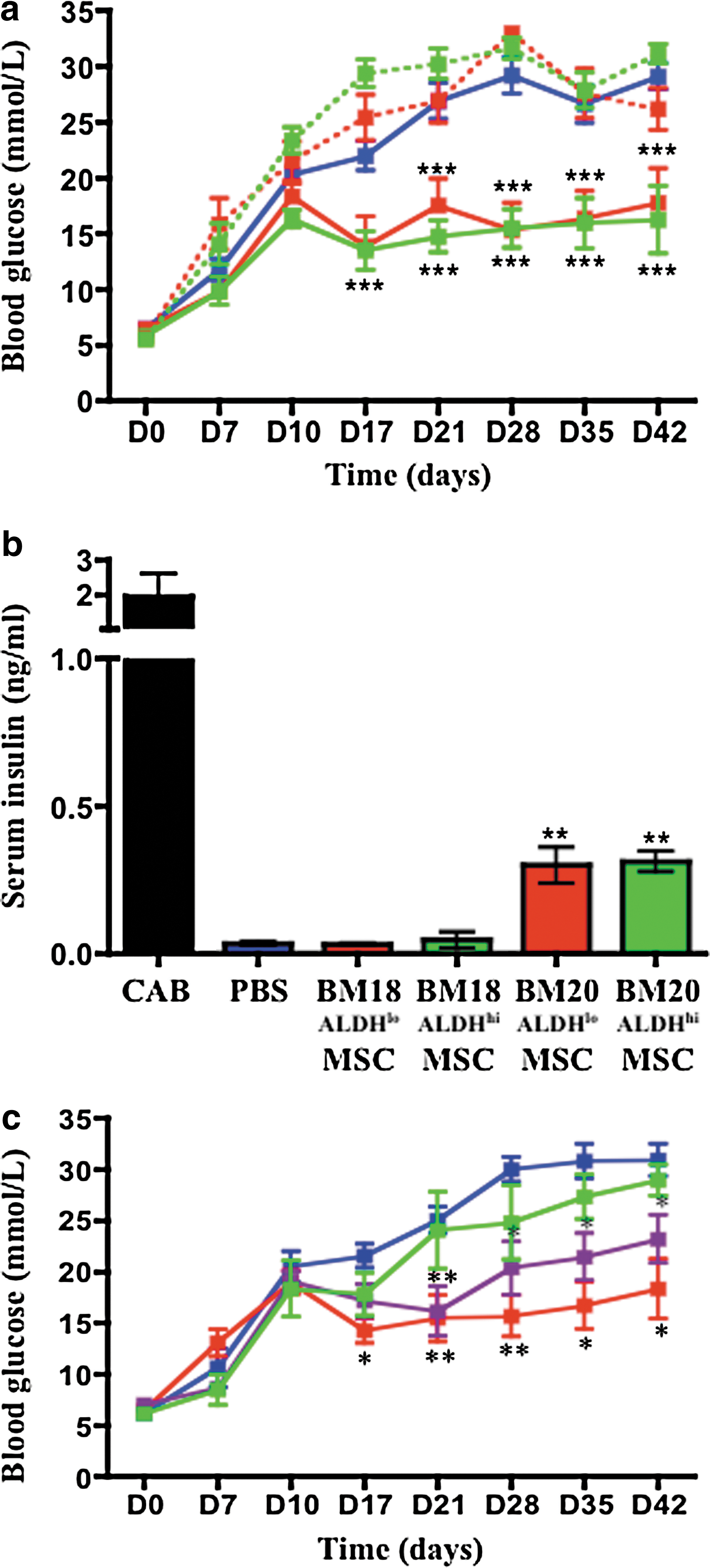
Transplantation of regenerative human BM ALDHlo/hi MSC at early passage improved blood glucose and increased serum insulin in STZ-treated recipients. STZ-treated (35 mg/kg/day, days 1–5), hyperglycemic (15–25 mM) NOD/SCID mice were tail vein injected at day 10 with PBS, or 5×105 ALDHlo or ALDHhi MSC from BM18 (nonregenerative), or 5×105 ALDHlo or ALDHhi MSC from BM20 (regenerative).
Prolonged expansion of human MSC diminished hyperglycemic reduction
To quantify the effect of MSC expansion on the induction of islet regeneration, ALDHhi MSC (BM20) were expanded for 2–4 passages, 5–6 passages, or 7–8 passages, before transplantation. Each MSC cohort possessed similar viability and expressed characteristic stromal surface markers (>95% CD90+, CD105+, and CD73+) and multipotent differentiation capacity at harvest (data not shown). Compared with PBS controls, transplantation of early passage (P2–P4) MSC showed sustained improvement in glycemia, P5–P6 MSC induced intermediate glycemic reduction, and late-passage (P7–P8) MSC did not significantly improve glycemia (Fig. 3c). Despite no evidence of population-specific differentiation or senescence in vitro, prolonged expansion of ALDHhi MSC diminished the capacity to improve hyperglycemia after transplantation (Fig. 3c).
Uncultured ALDHhi cells and ex vivo expanded MSC induced endogenous islet regeneration with different characteristics
To characterize the endogenous mechanisms by which transplanted ALDHhi cells support islet regeneration, pancreata of transplanted mice were stained for murine insulin (Fig. 4a–d) and analyzed for islet number (Fig. 4e), islet size (Fig. 4f) and total beta cell mass (Fig. 4g). STZ administration induced 10-fold reduced islet number (0.16±0.03 islets/mm2), and 2-fold reduced islet circumference (225±25 mm) compared with CAB-injected controls, confirming severe islet damage at day 10. Although transplantation of uncultured BM-derived ALDHlo or ALDHhi cells had no effect on islet number (Fig. 4e), we observed significantly increased islet size after ALDHlo or ALDHhi cell transplantation (Fig. 4f). In contrast, transplantation of BM20-derived ALDHlo or ALDHhi MSC increased the number of insulin+ beta cell clusters (Fig. 4e), whereas islet size remained similar to PBS-injected controls (Fig. 4f). As an overall indication of endocrine recovery, transplantation of BM ALDHhi cells or BM20-derived MSC significantly increased total beta cell mass compared with PBS controls (Fig. 4g). Thus, recovered beta cell mass occurred through increased islet size (but not number) after transplantation of uncultured ALDHhi progenitors, or by increased islet number (but not size) after transplantation of ALDHlo or ALDHhi MSC, suggesting that distinct regenerative mechanisms were activated by the different progenitor subtypes administered.
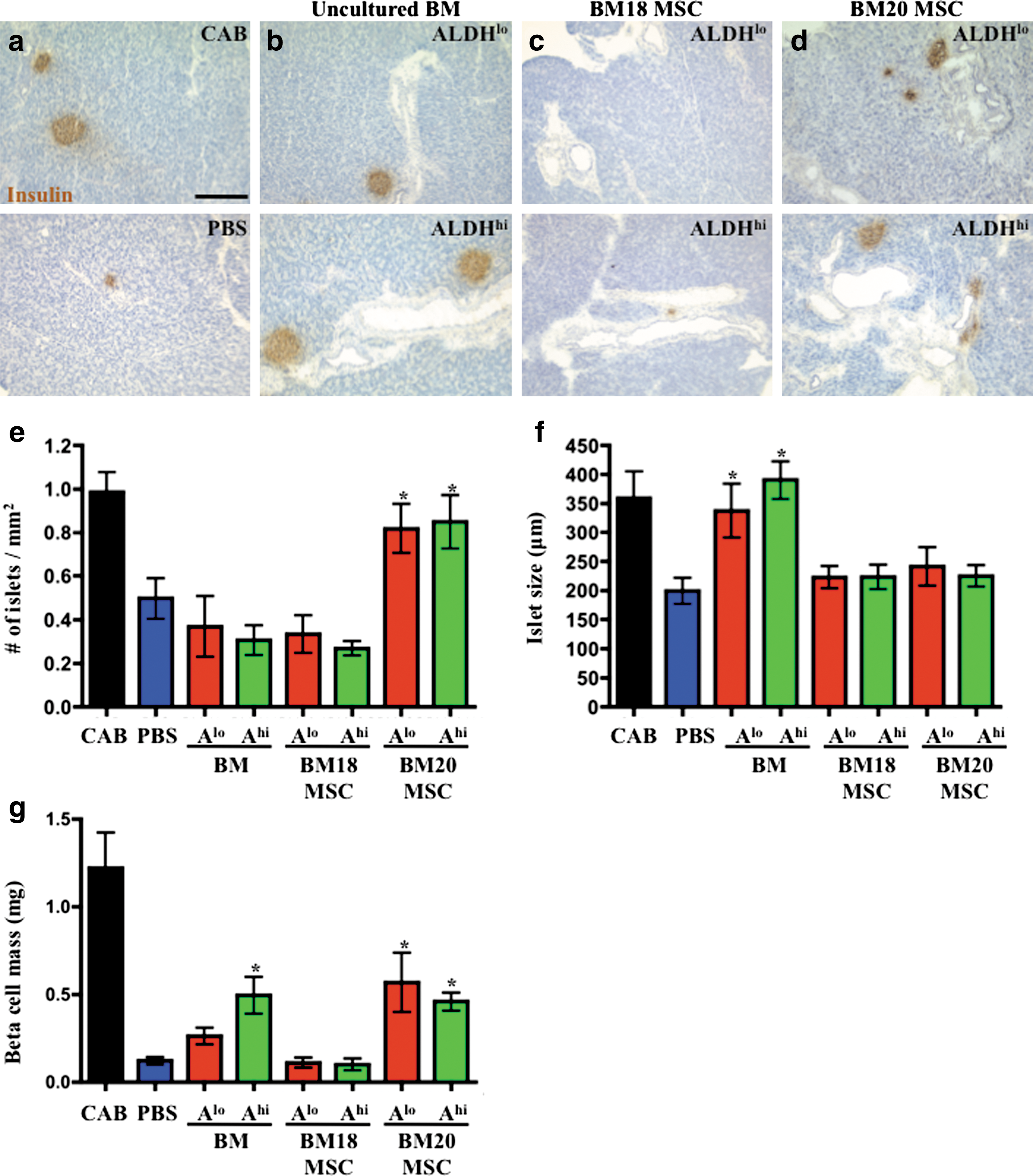
Distinct human progenitor subtypes stimulate endogenous islet regeneration via divergent mechanisms. Representative photomicrographs of insulin+ islets at day 42 in mice injected with
Transplantation of uncultured ALDHhi cells increased islet vascularization
We postulated that improved hyperglycemia (Fig. 1) and increased islet size (Fig. 4f) after human ALDHhi cell transplantation may correlate with progenitor-stimulated islet angiogenesis [18]. Thus, insulin+ islets were costained with vWF+ at day 42 (Fig. 5a–c), a marker of functional endothelial cells within blood vessels. CAB- and STZ-injected control mice showed equivalent islet microvessel densities at day 10, indicating that STZ did not initially target islet microvasculature despite significant beta cell damage (data not shown). Compared with PBS-injected controls, mice transplanted with BM-derived ALDHlo cells or ALDHhi cells showed augmented islet vascularization (Fig. 5d). In contrast, transplantation of ex vivo expanded ALDHlo or ALDHhi MSC from BM20 did not augment islet microvessel density (Fig. 5d). Vessel formation was islet specific, as extra-islet pancreatic tissue vascularity was unchanged after transplantation of any cell type (Fig. 5e). These data indicated that the transplantation of ALDH-purified progenitor cells, but not ex vivo expanded MSC, activated an islet-specific angiogenic program, thereby improving insulin release into the circulation.

Transplantation of human ALDHhi progenitor cells increased islet vascularization. Representative photomicrographs of vWF+ blood vessels (green) associated with insulin+ islets (red) 42 days after injection of
Transplantation of uncultured ALDHhi cells induced endothelial and beta cell proliferation
To further investigate the increase in islet size and vascularization, we analyzed EdU incorporation into proliferating cells within islets and associated micorovasculature at onset of blood glucose improvement (day 17) and at the end of our protocol (day 42). In PBS-injected controls, proliferating cells associated with vWF+ blood vessels were extremely rare, indicating a slow turnover of endothelial cells after STZ damage (Fig. 6a). At both early and late time points, EdU+ cells associated with vWF+ blood vessels were numerous after BM-derived ALDHhi cell transplantation (Fig. 6b), confirming sustained vessel remodeling. However, similar to PBS-transplanted controls, transplantation of ex vivo expanded ALDHlo or ALDHhi MSC from BM20 failed to induce significant proliferation of vWF+ cells (Fig. 6c).
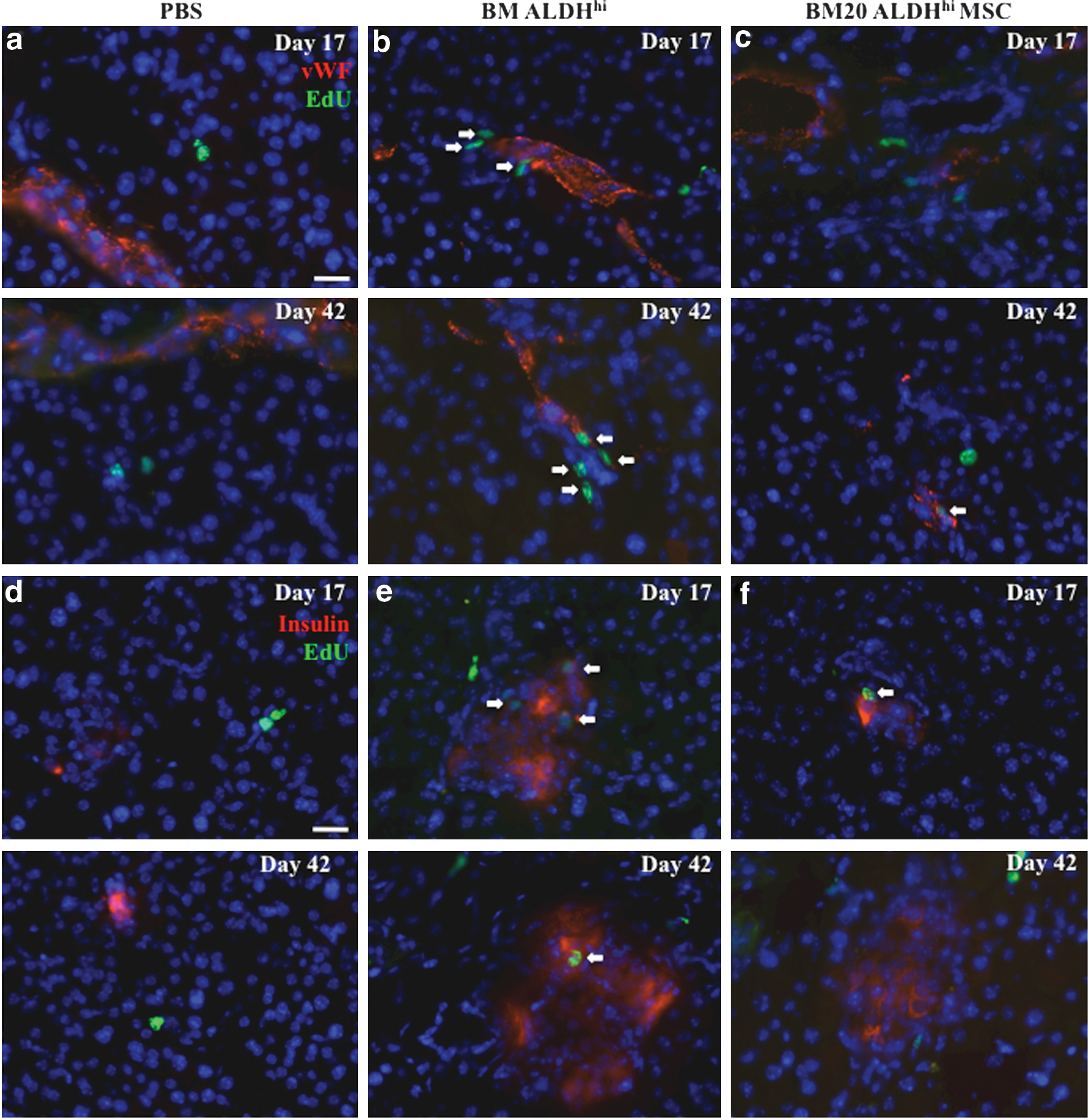
Transplantation of human BM-derived ALDHhi cells induced proliferation of murine vWF+ blood vessels and insulin+ islets. Representative photomicrographs of EdU-labeled proliferating cells associated with vWF+ blood vessels at day 17 (upper) or day 42 (lower) after STZ injection (35 mg/kg/day, days 1–5) followed by transplantation at day 10 with
Analysis of EdU incorporation into insulin+ islets showed that PBS-injected control pancreata did not contain proliferating insulin+ beta cells at early or late time points (Fig. 6d) after STZ damage. EdU+ beta cells were detected at days 17 and 42 after transplantation of BM-derived ALDHhi cells (Fig. 6e). Although EdU+ beta cells were rare in small beta cell clusters at day 17 after ALDHlo or ALDHhi MSC transplantation, beta cell proliferation was not detected at day 42 (Fig. 6f). Therefore, sustained beta cell proliferation was associated with the administration of uncultured ALDH-purified cells, whereas transplantation of cultured regenerative BM20 MSC resulted in smaller islets with limited or attenuated beta cell proliferation.
Transplantation of ALDH-purified MSC increased islet association with ducts
Similar to embryonic development, islet neogenesis may be initiated in ductal regions to restore beta cell mass after pancreas injury [12,13]. Since transplantation of BM20-derived ALDHlo or ALDHhi MSC increased the production of small beta cell clusters (Fig. 4e, f), we postulated that new islet formation would be most evident in ductal regions after transplantation. Representative photomicrographs show insulin+ islet location with regard to ck19+ ductal structures at days 17 and 42 after PBS, uncultured ALDHhi cell, or BM20 ALDHhi MSC transplantation (Fig. 7a–c). Although islet distribution after transplantation of uncultured BM ALDHhi cells was similar to PBS controls (Fig. 7d, e), there was a significant increase in islets with direct contact to the ductal epithelia at days 17 (Fig. 7d) and 42 (Fig. 7e) after ALDHlo or ALDHhi MSC transplantation. Thus, the ductal region represented a specific niche for islet regeneration after ALDH-purified MSC transplantation.

Transplantation of ex vivo expanded ALDHlo or ALDHhi MSC increased islet association with ck19+ ducts. Representative photomicrographs of insulin+ islets associated with ck19+ ductal epithelia at day 17 (upper) and day 42 (lower) after STZ injection (35 mg/kg/day, days 1–5) followed by transplantation at day 10 with
Transplantation of BM-derived MSC did not activate Ngn3-expression
Xu et al. have shown that newly formed islets emerge in ductal regions from Ngn3+ endocrine precursor cells after duct ligation [16]. To further characterize the mechanism of islet formation in ductal regions after the transplantation of human ALDH-purified MSC, we stained for insulin- and Ngn3-expression throughout the regeneration process (days 10, 14, 17, and 42). Representative photomicrographs show that nuclear localization of Ngn3 was abundant in embryonic day 18 pancreatic islets (Fig. 8a, upper), but that Ngn3-expression was absent at day 10 in adult mice treated with STZ (Fig. 8a, lower). Although insulin expression was induced in ductal regions by day 14, we did not detect endogenous Ngn3-expression in islets or ducts at days 14, 17, or 42 after ALDHlo or ALDHhi MSC transplantation (Fig. 8b, c). Thus, islet regeneration in STZ-treated mice after human MSC transplantation was suggestive of the induction of a neogenic program, but without evidence of transition through an Ngn3-expressing endocrine precursor.
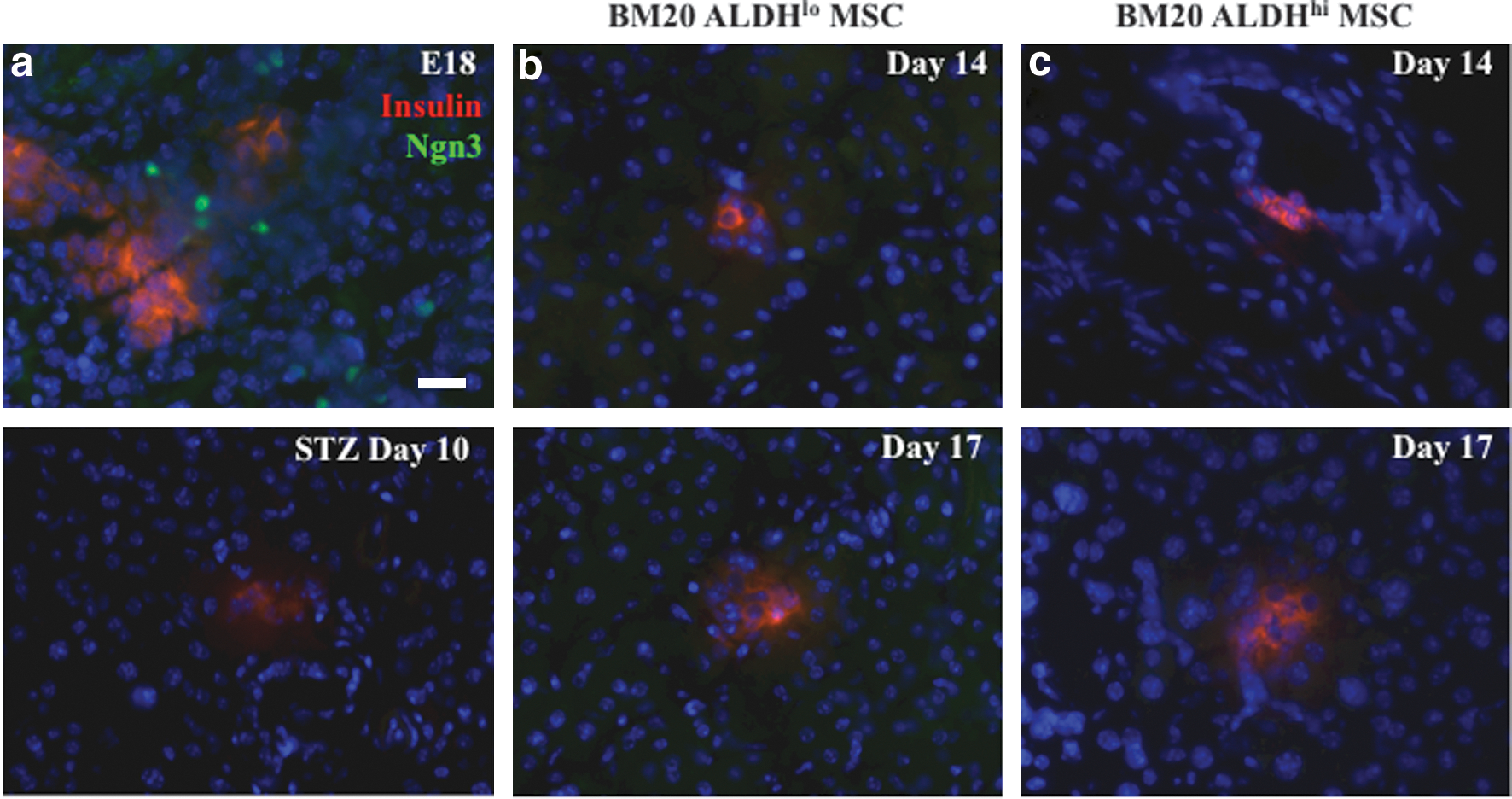
Transplantation of ex vivo cultured BM MSC did not activate endogenous Ngn3 expression. Representative photomicrographs of mouse pancreas sections stained for insulin and Ngn3 at
Discussion
Transplantation of uncultured human ALDHhi cells or culture-expanded MSC into hyperglycemic NOD/SCID mice revealed that the activation of distinct islet regenerative programs was dependent on lineage restriction of the progenitor cells administered. BM-derived ALDHhi cells stimulated islet-specific vascularization and beta cell proliferation, thereby resulting in increased islet size. In contrast, transplantation of culture-expanded human MSC did not stimulate islet proliferative or pro-angiogenic programs, but increased total beta cell mass via the formation of beta cell clusters associated with the ductal epithelium, suggestive of a putative neogenic mechanism, but without endogenous Ngn3-expressing endocrine precursor activation [16].
We have previously shown that BM-derived ALDHhi cells contain pro-angiogenic progenitor cells that improve perfusion after transplantation into mice with acute limb ischemia [18]. Here, we demonstrate that human ALDHhi cells similarly enhance vascularization in STZ-treated islets. Beta cells are known to secrete angiogenic cytokines [24] and chemokines [25] that promote recruitment of pro-angiogenic cells to damaged islets. Interestingly, increased islet vascularization was associated with increased murine beta cell turnover, and transplanted human cell differentiation to insulin-positive beta cells was not observed. During development, signals from inflammatory leukocytes and activated endothelium have also been shown to be crucial for beta cell development and function [26]. However, in immune-deficient NOD/SCID mice, we did not observe increased infiltration of murine macrophages (CD11b+) in islets after STZ treatment and subsequent transplantation. Thus, human ALDHhi cells contain hematopoietic and nonhematopoietic progenitor subtypes [27,28] that could potentially contribute to islet regeneration and revascularization [18]. Further study of progenitor cell communication at the microvascular and beta cell interface during islet regeneration is warranted to uncover the specific progenitor subtypes and secreted target molecules that stimulate endogenous beta cell proliferation and islet vascularization.
Extended passage of regenerative MSC diminished the capacity to reduce hyperglycemia after transplantation, warning that islet regenerative activities of MSC may be compromised by expansion in serum-rich media under atmospheric oxygen tension [29,30]. Differences between early- and late-stage cultures of MSC can impact their regenerative capacity, with upregulation of cell division genes at early stages, and subsequent upregulation of differentiative genes at later stages [31]. Optimization of lineage-specific progenitor cell isolation and expansion require further development to assure efficient delivery of paracrine stimuli to regenerating islets.
Lee and colleagues have previously shown that transplanted MSC possess significant capacity to regenerate islets with minimal long-term engraftment in the pancreas [10]. Similarly, we observed improved hyperglycemia after transplantation of ALDH-purified MSC, but did not detect significant human cell engraftment in the recipient mouse pancreas. Although some MSC have been shown to migrate to the pancreas after STZ treatment [10,32], our data suggest that MSC can act from distant sites to exert paracrine or hormone action in the pancreas. Nevertheless, if local paracrine or contact-dependent mechanisms prove beneficial for robust islet regeneration, direct intra-pancreatic transplantation may further promote islet function by increasing the number of regenerative cells at the site of injury. Thus, more efficient delivery of regenerative MSC to the pancreas or increased survival of MSC at ectopic sites are predicted to further improve the efficiency of islet recovery.
Our data suggest that human MSC may promote islet regeneration through a putative neogenic mechanism. Previous studies have suggested that new islets can be formed from endocrine progenitor cells in ductal regions to restore beta cell mass after partial pancreatectomy [12,33 –36]. Recently, an Ngn3+ multipotent progenitor has been identified in the ductal lining after ductal ligation [16]. Since transplantation of regenerative human ALDHlo or ALDHhi MSC minimally stimulated proliferative or pro-angiogenic programs, but improved glycemic control via an increase in the number of beta cell clusters associated with the ductal epithelium, we investigated whether transplantation of human MSC could promote neogenesis via activation of an Ngn3-expressing precursor. However, we did not observe the induction of endogenous Ngn3-expression in our STZ damage model at 4, 7, or 32 days post-transplantation. Ngn3 is expressed transiently by endocrine progenitors [16], therefore, we cannot rule out the possibility that Ngn3 could have been briefly expressed during MSC-stimulated regeneration.
Our data outline several important caveats for the development of regenerative therapies for diabetes. Autologous BM cells can be isolated in a clinically relevant fashion using ALDH activity, and ALDH-purified MSC are amenable to ex vivo expansion to increase regenerative cell numbers. The divergent regenerative mechanisms presented suggest that a combinatorial cellular therapy may be an effective approach to promote endogenous islet regeneration, using MSC to promote islet formation and pro-angiogenenic cell administration to stimulate subsequent beta cell proliferation and islet vascularization. However, since sample variability and prolonged expansion reduce regenerative capacity, the specific factors secreted by regenerative cell lines warrant extensive investigation to further characterize the regenerative niche. Detailed understanding of the endogenous factors that govern progenitor cell-induced islet regeneration will aid in the development of future therapies to treat diabetes.
Footnotes
Acknowledgments
The authors wish to thank Dr. Lynne Postovit for critical review of the manuscript, and Kristen Chadwick and Karen Morley at the London Regional Flow Cytometry Facility for cell sorting. This work was supported by Johnson and Johnson, Betalogics Division (Skillman, NJ), the Krembil Foundation (Toronto, ON), The Juvenile Diabetes Research Foundation (New York, NY; 1-2005-1173), the Canadian Institutes of Health Research (Ottawa, ON; MOP 86702 and MOP 86759), and a New Investigator Award from the Heart and Stroke Foundation of Canada (Ottawa, ON; to D.A.H). G.I.B is supported by an Ontario Graduate Scholarship (Toronto, ON).
Author Disclosure Statement
No competing financial interests exist.