
Abstract
Skin samples taken from 6 female patients receiving allogeneic bone marrow transplants (BMT) from male siblings (n=5) or from unrelated human leukocyte antigen (HLA)-matched male donor (n=1) due to hematological malignancies were studied for the presence of donor cells. One nontransplanted male and 1 female control that received female BM were used as further controls of the technique. Skin biopsies were taken from the scalp and the back from each patient 12–16 years after the successful BMT. We have found donor chimerism in all of the 6 patients in both of their biopsies. Using single and double immunostainings in combination with Y chromosome hybridization, we observed that there are cytokeratin-expressing donor-derived cells in the epidermis of all the 6 patients, the numbers being slightly higher in the scalp (0.37%–1.78%) than in the back (0.32%–1.08%) biopsies. The indication for BMT, and the age of the patient did not seem to have any effect on the numbers found. A few of the double-labeled cells also stained for Ki67, a marker of cellular proliferation, suggesting that the engrafted cells were able to further divide in the epidermis. In 2 patients we observed patches of donor keratinocytes within the epidermis, suggesting a clonal origin. We conclude that in agreement with some and in contrast to other published studies, BM-derived circulating cells are able to engraft in the human skin and to further proliferate there and thus contribute to tissue renewal. These data raise the possibility to use BM cells in regenerative medicine to help in extended injuries, large surface burns, or lack of skin due to other reasons.
Introduction
I
Methods
All patients participated voluntarily in the study and were enrolled in a protocol approved by the local ethics committee (TUKEB, 214/2004). Incisional biopsies (1 cm×0.5 cm×0.5 cm) were taken under local anesthesia (Lidocain-Adrenalin USP) and frozen in embedding media. We enrolled 6 patients (for individual data see Table 1) with gender-mismatched transplants, 1 female patient who received female BM and 1 nontransplanted male control. From all patients, 3 sections were taken from the back skin biopsies and 3 samples from the scalp biopsy. The biopsies were cut in a cryostat at 10 μm thickness, and the sections were placed on poly-l-lysine-coated slides and stored frozen at −80°C until ready for processing. For processing, the tissue was removed from the freezer and without thawing, placed into 4% paraformaldehyde/1× phosphate-buffered saline (PBS) for 10 min. The slides were then rinsed 3×3 min in 1×PBS followed by 10 min in 1× Universal Power Block (Cat. No. HK085-5K; BioGenex).
ALL, acute lymphoid leukemia; AML, acute myeloid leukemia; CML, chronic myeloid leukemia; TBI+Cy, total body irradiation and cyclosporine; bu, busulphan; sib, sibling; MUD, matching unrelated donor; BMT, bone marrow transplants.
Combined double immunofluorescence and Y chromosome fluorescence in situ hybridization
Three antibodies were used to identify keratinocytes (Pancytokeratin against cytokeratin 1,5,10 and 14), hematopoietic cells (CD45), and proliferating cells (Ki67) in combination with the Y chromosome that was labeled using in situ hybridization and a human Y chromosome-specific probe [4].
Cytokeratin immunofluorescence
After rinsing the slides quickly in deionized water followed by a 1×PBS rinse, the slides were processed as indicated above except using a mouse anti-human cytokeratin (Pan-CK against cytokeratin 1,5,10 and 14) antibody (Cat. No. VP-C417; Vector Labs, Inc.) at a dilution of 1:150. This was followed by incubation in mouse SuperPicture (Cat. No. 87–9163; Invitrogen Corp.). Positive staining was observed in green with a tyramide plus FITC system (Cat. No. NEL741001KT; Perkin-Elmer, Inc.). Before continuing with the second immunostaining, the slides were briefly microwaved in 10 mM Citrate pH 6.1 for 5 min at 50% power. This blocks any unreacted peroxidase molecules (potentially leftover from the first immunostaining), which could possibly give a nonspecific staining for the second primary antibody.
Ki67 immunofluorescence
The slides were then incubated with a cell proliferation antibody, rabbit anti-Ki67 (Cat. No. 15580; GeneTex, Inc.), at a 1:200 dilution, overnight at 4°C. The next day the slides were rinsed 3×3min in 1×PBS. Endogenous peroxidase was suppressed for 10 min using a commercially prepared solution (Cat. No. S2001; Dakocytomation). Rabbit SuperPicture (Cat. No. 87-9263; Invitrogen Corp.) was applied to the sections for 30 min at room temperature. After rinsing 3×3 min in PBS, positively stained cells were observed in far red (yellow color) using a tyramide plus CY5 system (Cat. No. NEL745001KT; Perkin-Elmer, Inc.) at a dilution of 1:1000 for 10 min. The slides were then rinsed well in several changes of 1× PBS.
CD45 immunofluorescence
The slides were then incubated in a mouse anti-human CD45 antibody (BD Biosciences; Cat No. 555485) at a 1:200 dilution, overnight at 4°C. The next day the slides were rinsed 3×3 min in 1×PBS. Endogenous peroxidase was suppressed for 10 min using a commercially prepared solution (Cat. No. S2001; Dakocytomation). Rabbit SuperPicture (Cat. No. 87-9263; Invitrogen Corp.) was applied to the sections for 30 min at room temperature. After rinsing 3×3 min in PBS, positively stained cells were observed in far red (yellow color) using a tyramide plus CY5 system (Cat. No. NEL745001KT; Perkin-Elmer, Inc.) at a dilution of 1:1000 for 10 min. The slides were then rinsed well in several changes of 1× PBS.
Y chromosome fluorescence in situ hybridization
Following the double immunostaining, the slides were processed for nonradioactive in situ hybridization to detect the Y chromosome. The template for this probe was generated from human genomic DNA by using primers that amplified a 1.3-kb long DNA piece of the human Y chromosome. The probe was labeled with digoxigenin-uridine 5′–triphosphate (Cat. No. 1 175 025; Roche Applied Science). The digoxigenin-labeled probe was then applied to the slides and hybridized at 81°C for 12 min followed by 30 min at 55°C. Slides were next washed in SSC, including a 65°C strigency wash. A peroxidase-conjugated anti-digoxigenin antibody (Cat. No. 11 207 733 910; Roche Applied Science) was then applied at 1:400. Positive Y chromosome signal was observed in red by applying the high affinity substrate of HRP, Alexa-tyramide-594 (Perkin-Elmers). Finally, DAPI (Cat. No. D1306, Invitrogen, Corp.) was applied as a general chromosomal stain of all cell nuclei.
Quantitation
We used 3 nonconsecutive sections (at least 300 μm apart) from each biopsy sample and took pictures of either the whole available section or every second field depending on how large the sections were. In general, we ended up with at least 20 fields from each sample and counted all the cell nuclei (stained with DAPI) using the NIH ImageJ program (Mac Vs 1.43). We then manually counted all Y-positive cells as well as the double-Y and CK-positive cells.
Results
We first established—similarly to our previous studies—that the sensitivity of the Y chromosome hybridization is over 90% (ie, the percentage of cells in a male section that are labeled with the Y chromosome) (Fig. 1A) and the specificity of the probe is 100% (ie, no positive nuclei were found in the female patient samples). In all samples studied, we observed Y chromosome-positive nuclei in epithelial layers and hair follicles (Fig. 1B) as well as in the epidermal layers, and used the CD45 immunostaining to exclude cells of hematopoietic origin. Most of the Y-positive cells in the epithelial layer also stained for CK confirming the epithelial nature of those cells (Fig. 1C). When Ki67 was added as a nuclear proliferation marker, we observed rare occasional cells that were triple positive (Y chromosome, cytokeratin, and Ki67) (Fig. 1D–F, G–J). In 2 patients we observed a specific distribution of Y-positive epithelial cells: the Y-positive cells seem to group in one area across the heights of the epidermal layer leaving long stretches of epithelium on both sides containing exclusively host-derived cells. We used regular microscopic images for the quantification, but performed Z series at the interval of 0.5 μm in our images to demonstrate the exact localization of the Y label, the CK, and the Ki67 to a certain cell.
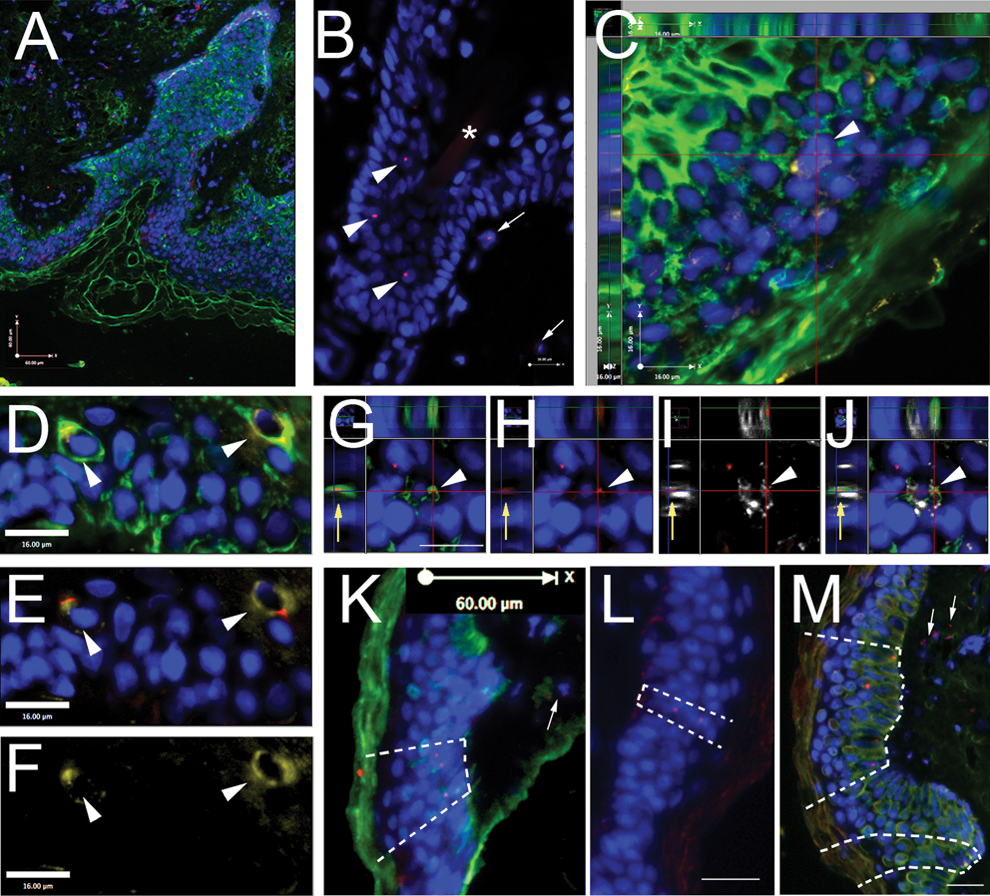
Presence of donor-derived cells in the skin of patients 12–16 years following gender-mismatched BM transplant. In all panels blue color is DAPI (chromosomal marker for nuclei), green labels cytokeratin (fluorescein isothiocyanate), red dots label Y chromosome (Alexafluor-594), and yellow
Our results demonstrated the presence of double-positive (Y and CK) in all samples examined (Fig. 1), with a relatively small patient-to-patient variation. The percentage of Y-positive epidermal cells ranged between 0.37% and 1.78% (scalp) and 0.32% and 1.08% (back), indicating a slightly, but consistently higher number in the scalp than in the back samples. In addition to the double-positive cells we also counted the number of epidermal donor-derived cells that were not epithelial in nature (ie, not CK positive). The number of these cells, most of them CD45 positive, ranged between 7.5% and 17.5% (scalp) and 3.7 and 13.6 (back) (Table. 1 and Fig. 2). The histology of the hybridized sections showed a columnal distribution of groups of Y-positive keratinocytes in between long stretches of cells of exclusively host origin (Fig. 1. K–M).

Columnar graph showing the percentage of Y chromosome-positive cells among the cytokeratin-positive (left panel) and the cytokeratin-negative (right panel) cells. Data regarding the scalp are represented by black columns, and data representing the data regarding the back skin biopsies are represented by gray columns. Standard error of the mean of 3 samples is shown.
Discussion
In addition to the constant epidermal renewal, the so-called physiological turn over, the cutis also frequently needs repair due to injuries. The cutaneous stem cell reservoir evoked the interest of basic biologists and dermatologists and there are several good reviews describing these stem cell pools [5 –7]. The epidermal stem cells seem to maintain the skin integrity under physiological turnover, whereas the bulge stem cells contribute more to the everyday repair [3]. Krause showed the presence of male cells in the skin at a frequency of 1.2%–2.7% using fluorescence in situ hybridization analysis after she transplanted female mice with a single male hematopoietic stem cell (HSC) [8]. Following wound healing the mouse BM-derived cells (BMDC) were reported to contribute to epidermal regeneration and differentiate into keratinocytes [9] as well as migrate into the hair follicle, the sebaceaous gland, and the epidermis, and their engraftment is significantly upregulated in wounded skin [10]. Due to the high cutaneous turnover rate, one wonders if these skin stem cell pools need renewal themselves, such as replacement by circulating cells. In the murine epithelium, the hematopoietic progenitor marker, CD34, is a specific marker of bulge cell keratinocytes [11], but in the human, CD34 is present in stem cells of the outer root sheath [12 –14]. Since these cells lack CK15, a type I keratin, they were suggested to be transit amplifying (TA) cells, progeny of the bulge stem cells [13,14]. In a groundbreaking study, Deng et al. transplanted fluorescently labeled mesenchymal stem cells from white male mice into lethally irradiated black female mice and observed that the recipient mice grew white hair all over the body. The authors concluded that BMDCs give rise to functional skin cells in an injury setting [15]. In a human study, Korbling identified donor cells in the skin of female recipients of male HSC stem cells using a combination of immunohistochemistry for specific epithelial antigens and fluorescence in situ hybridization [16] and found 2%–7% chimerism in the biopsies. Similar numbers (1.6%–7.8% of donor-derived keratinocytes) were reported by Murata et al. in patients with acute graft versus host disease (GVHD) who previously received HSC transplant from male donors [17], in addition to 2.2%–9.4% chimeric endothelial cells, which were also shown in human samples by Suzuki [18]. On the contrary, Hematti et al., who used similar samples but when performed polymerase chain reaction on cultured keratinocytes, could not detect any donor-derived cells [19]. Using fingernail shavings from gender mismatched transplant patients, 2 independent publications demonstrated high chimerism (9%–73%) using short tandem repeat polymerase chain reaction [20,21]. These data suggest that—especially in injury settings—hematopoietic progenitors do indeed contribute to skin regeneration in both mice and humans.
Our group of patients (aged between 27 and 48 years) included 6 women, all of whom underwent BM transplantation due to malignant hematological diseases. All of them received BM (as opposed to granulocyte colony-stimulating factor [GCSF]-induced peripheral blood stem cells) and the skin biopsies were taken between 12 and 16 years after transplant. The treatment protocols before BM suppression and following the transplant were almost identical in all of them. Five patients received transplants from siblings, whereas one had a matching unrelated donor. This 1 patient had the highest level of chimerism in the scalp, which might be a simple coincidence given the restricted number of samples studied. It has been established that the epidermis contains 2 major types of stem cells: the epidermal proliferative unit and the stem cell niche in the bulge region. The observed patchy distribution of the donor-derived cells in our patients suggests that a circulating cell might have become an epidermal proliferative unit stem cell and gave rise to a lineage of keratinocytes in that area. A theoretical possibility should also be mentioned that the circulating BMDCs might become TA cells without becoming stem cells first, as already suggested in mouse studies [22]. This would explain that although there are columns of Y-positive cells, these do not spread over time, that is, the TA cells, unlike the mother stem cells, lose their ability to generate new stem cells. It seems that the majority of the donor-derived keratinocytes have a limited proliferation capacity based on the colocalization of Ki67, an accepted marker of proliferation, with the Y chromosome and CK.
The observations in this study strongly support the idea that circulating cells after BM transplantation can arrive at the skin and participate in its regeneration by establishing themselves as skin stem cells. The toxicity of chemotherapy resulting in hair loss constitutes an injury-induced need to regenerate hair. In Hematti's earlier negative study [19], all patients received blood progenitors collected from the peripheral blood after GCSF induction. In our study all patients received BM. A possible explanation of the discrepancy might be that other than hematopoietic cells in the BM participate in the process we observed. These other cells could be mesenchymal cells, or a unique as yet unknown skin progenitor that lives in the BM. The possibility of donor and recipient cell fusion seems to be very unlikely; for example, in his mouse studies Brittan excluded fusion as a cause for donor-derived epithelial cells by in situ hybridization for both Y and X chromosomes [10]. In addition, in our previous study in cheek epithelial cells from gender-mismatched BMT patients, we also demonstrated the lack of fusion by X and Y chromosomal hybridization, as well as the donor origin of some [23]. It is theoretically possible that keratinocytes might phagocytose Y chromosome-positive circulating blood cells, and then appear to be Y chromosome-positive themselves. Although keratinocytes have been shown to phagocytose pathogens and melanosomes [24,25], there is no evidence that they take up blood cells, and this seems unlikely to have occurred.
Our results suggest that, indeed, there is a contribution of circulating BMDCs to the skin stem cell population. This contribution is modest, but might be boosted by danger signals, when more repair is needed. The exact nature of such signals should be further explored, since these cells might be helpful when regeneration is needed quickly and in large quantities, like in case of trauma or burn disease.
Footnotes
Acknowledgments
The authors appreciate the help provided by G. Szombath, M. Sardy, and M. Krepuska in coordinating the biopsies and contributing technical help. This research was supported by the Division of Intramural Research program of NIDCR, in the Intramural Research Program, NIH, DHHS.
Author Disclosure Statement
No competing financial interests exist.