
Abstract
Recently, there has been growing support for the cancer stem cell (CSC) hypothesis, which states that primary tumors are initiated and maintained by a small subpopulation of cancer cells that possess “stem-like” characteristics. CSCs have been identified in many tumor types, including hepatocellular carcinoma (HCC). The dye, Hoechst 33342, has been used to enrich CSCs into a side population. Alternatively, liver CSCs (LCSCs) can be identified by several cell surface antigens, including CD133, CD90, CD44, EpCAM, and CD13. In this review, we summarized the recent evidence regarding LCSC markers and discussed the origin and function of these markers. LCSC markers are essential to identify and isolate these cells, to develop future therapies targeting CSCs, and to predict prognosis and efficacy of these therapies. However, definite LCSC markers are still controversial, because none of these markers is exclusively expressed by LCSCs in HCC. By combining several positive or negative markers, it may be possible to isolate and identify CSC fractions beyond the ability of each individual assay. By grouping LCSC markers according to their cellular origin, the properties of LCSC markers may be better studied and new markers may be found. Lastly, markers could be used to estimate the number of LCSCs and therefore predict outcomes. From our point of view, selecting HCC tissue samples from patients with different prognoses and detecting expression patterns of marker combinations may be a new method to identify new and unique markers.
Introduction
Liver cancer
L
Cancer stem cells
Cancer stem cells (CSCs) comprise a small fraction of tumor cells that possess characteristics of stem cells, most notably the ability of self-renewal and differentiation into heterogeneous nontumorigenic cancer cells that make up the bulk of the tumor. CSCs are responsible for maintaining the growth of neoplastic tissue [3,4] and metastasis and recurrence. CSCs are also relatively resistant to anticancer drugs [5]. The existence of CSCs was first demonstrated in acute myeloid leukemia [6,7]. So far, CSCs have been demonstrated in a variety of solid tumors, including tumors of the breast, brain, prostate, pancreas, and colon [8].
Putative Markers of Liver CSCs
Recently, there has been mounting evidence in support of the existence of CSCs in HCC. These cells can be identified by several cell markers, including CD133, CD90, CD44, epithelial cell-adhesion molecule (EpCAM), CD13, OV6, and aldehyde dehydrogenase (ALDH) enzymatic activity, or by selecting for the side population (SP) cells by Hoechst dye staining. However, CSCs sorted from different laboratories have shown great heterogeneity. Cells with different markers harbor similar CSC properties, but seldom cross-express the selecting markers. Consequently, generally accepted markers of CSCs in HCC have not been identified. Putative liver CSC (LCSC) markers are summarized in Table 1.
SP, side population; ALDH, aldehyde dehydrogenase; EpCAM, epithelial cell adhesion molecule.
ABCG2
SP cells are detected by their ability to efflux Hoechst 33342 dye by an adenosine triphosphate (ATP)-binding cassette (ABC) membrane transporter, such as MDR1 and ABCG2 [21]. Recently, SP cells were used in an attempt to isolate stem cell–like fractions in cancer cells [22]. Several studies have reported that SP cells could be purified from human HCC cell lines [9,23,24]. These SP cells purified from HCC cell lines harbor CSC-like properties and may be related to metastatic potential and therapeutic resistance of HCC. The polycomb gene product BMI1 plays an important role in the maintenance of SP cells in HCC [25].
Because it is not technically feasible to detect SP cells in HCC samples in situ, examination of these ABC transporters, such as ABCG2, might help localize CSCs. Described as the determinant of SP cells, ABCG2 was found to be intrinsically expressed in HCC tissues, and its expression pattern significantly influenced the levels of drug efflux from HCC cell lines [26]. However, ABCG2+ cells might contain a larger range of cells compared with SP cells [27].
CD133 (Prominin 1)
Human CD133, a 5-transmembrane domain glycoprotein, was originally identified as a cell surface antigen present on CD34+ hematopoietic stem cells (HSCs) [28]. It is an important cell surface marker for both stem cells and CSCs in various tissues including the liver. The role of CD133 as a CSC marker has been documented in cancers of the brain, prostate, pancreas, and colon [29 –34].
In 2006, Suetsugu and colleagues investigated CD133 in 3 liver cell lines and found that CD133 only expressed in the Huh7 cells. Functional studies with sorted CD133+ and CD133− cells demonstrated that CD133+ cells from Huh7 manifested a higher in vitro and in vivo tumorigenic potential [10]. Yin et al. found similar results in a CD133+ fraction isolated from the HCC cell line SMMC-7721 [11]. Studies of Ma et al. also found that the CD133+ cells had significantly greater tumorigenicity in immunodeficient mice and higher colony-forming efficiency and proliferation ability in vitro, and these cells could be induced to differentiate into nonhepatocyte-like lineages, supporting CD133 as a marker of liver cancer stem/progenitor cells [12].
Further studies found CD133+ cells to be more resistant to conventional chemotherapeutic drugs, such as doxorubicin and 5-fluorouracil, than their CD133− counterparts; the underlying mechanism of which required the preferential activation of the Akt/PKB and Bcl-2 survival pathway [5]. The CD133+ population in HCC has also demonstrated relative resistance to transforming growth factor-β (TGF-β)–induced apoptosis compared with CD133− cells. One mechanism of resistance to TGF-β–induced apoptosis in CD133+ CSCs is an activated mitogen-activated protein kinase pathway [35].
Oxygen levels have been shown to modulate CD133 expression in cultures of medulloblastoma and glioblastoma cells [36,37]. However, it has not been confirmed in HCC. Recent studies demonstrated that CD133 expression was regulated by DNA methylation in HCC. In this study, You et al. found that TGF-β1 was able to upregulate CD133 expression through inhibition of DNMT1 and DNMT3 expression and subsequent demethylation of promoter-1. The mechanism by which TGF-β induces CD133 expression is partially dependent on the Smads pathway [38].
The function of CD133 in HCC remains unknown. It may play an important functional role in the growth of tumor cells. Yao et al. showed that knockdown of CD133 decreased the colony-forming ability and altered the cell cycle distribution in Huh-7 human HCC cells [39]. Expression of CD133 was found to confer malignant potential by regulating metalloproteinases in human HCC [40].
According to CSC theory, only a minority of cells are believed to be stem cells in a population. Although several studies have found CD133 to be expressed in only a minute proportion of HCC cells [11,12], further analysis by flow cytometry revealed that the percentage of cells expressing CD133 significantly varied among several HCC cell lines (from less than 1% to more than 90%). CD133 was also present in ductular reactions in both acute and chronically damaged livers [41]. CD133 could be a biliary marker and hepatic progenitor cell marker in vivo [42]. Thus, CD133 alone is not sufficient to detect tumor-initiating cells in cell lines.
It should be noted that almost all CD133-related experiments carried out to date have detected the expression of AC133 and AC141 epitopes, rather than total CD133 protein. The monoclonal antibodies used against AC133 and AC141 were CD133/1 and CD133/2, respectively [10 –13]. Recent evidence suggests that CD133 mRNA seems to be constant, whereas the AC133 epitope is lost upon CSC differentiation in colon cancer [43]. Therefore, CD133 should be used with caution as an LCSC marker.
CD90 (Thy-1)
CD90 is a 25–37-kDa glycosylphosphatidylinositol-anchored glycoprotein expressed on many cell types, including T cells, thymocytes, neurons, endothelial cells, and fibroblasts, and is involved in cell–cell and cell–matrix interactions [44]. As a marker expressed on hepatic stem/progenitor cells during liver development [45], its expression has been identified in murine breast CSCs, prostate cancer, and primarily cultured CD133+ glioblastoma cells [46 –48]. Recent studies have suggested CD90 as another putative marker for liver CSCs.
Yang et al. found that the expression of CD90 correlated with tumorigenic potential of the cell lines tested [14,15]. To exclude lymphocytes from among all the cells marked by CD90, the authors combined its use with CD45 to isolate nonlymphoid CD90+ cells. CD90+ cells sorted from HCC cell lines and CD90+/CD45− cells isolated from tumor tissues and blood samples of liver cancer generated tumor nodules in immunodeficient SCID/Beige mice and could be serially transplanted into secondary and tertiary animals.
CD44
CD44 is a member of a family of transmembrane glycoproteins, which act mainly as receptors for hyaluronan. It is an adhesion molecule that helps tumor cell invasion and migration [49]. CD44 has been identified as a CSC marker in breast, pancreatic, colorectal, and gastric cancer [30,50 –52].
As mentioned earlier, Yang et al. found that most CD90+ cells also expressed CD44. Blockade of CD44 by a neutralizing antibody induced apoptosis of CD90+ cells, which prevented local and systemic tumor formation in mice. Further treatment of CD90+ CSCs with the anti-human CD44 neutralizing antibody induced apoptosis in vitro and suppressed CD90+ cell–induced local and systemic tumor engraftments in vivo [14,15]. Because expression of CD44 in HCC is related to a higher frequency of extrahepatic metastasis and shortened survival [53,54], the CD90+/CD44+ phenotype of CSC might explain the aggressive growth pattern of HCC. Zhu et al. demonstrated that an LCSC phenotype could be precisely defined by coexpression of CD133 and CD44 cell surface markers [13]. Taken together, CD44 appears to be more useful combined with other markers instead of being an independent marker of CSCs in HCC.
Epithelial cell-adhesion molecule
Recently, the EpCAM has been identified as a potential marker of LCSCs. EpCAM has been previously identified as a marker for stem/progenitor cells of adult liver and oval cells [55,56]. It is highly expressed in fetal hepatoblasts, hepatic stem cells, bile duct epithelium, and premalignant hepatic tissues and HCC, but not in most normal adult hepatocytes [16,57]. EpCAM plays a role in cell proliferation, migration, and mitogenic signal transduction [58].
EpCAM+ HCC cells were found to be more tumorigenic and invasive when compared with EpCAM− cells [17]. Yamashita and colleagues proposed a classification system defined by EpCAM and α-fetoprotein (AFP) and found that EpCAM+/AFP+ cells seemed to present characteristics of CSCs [16]. When xenografted in nonobese diabetic, severe combined-immunodeficient (NOD/SCID) mice, only EpCAM+ cells could efficiently initiate the development of invasive tumors, even after serial transplantation, whereas EpCAM− cells did not show these properties. Further, EpCAM has been shown to be a direct transcriptional target in the Wnt/β-catenin signaling pathway, which has been suggested to play an important role in governing the self-renewal of cancer cells [59]. EpCAM may serve as a biomarker for the activation of Wnt/β-catenin signaling, suggesting that Wnt/β-catenin signaling inhibitors may have a selective effect on EpCAM-positive HCC [60].
CD13 (aminopeptidase N)
Recently, CD13 has been identified as a novel cell surface marker for semiquiescent CSCs in human liver cancer cells [20]. CD13, also known as aminopeptidase N, is a membranous glycoprotein that plays important roles in cancer progression including cell proliferation, invasion, and angiogenesis [61,62].
Haraguchi et al. explored SP cells and found that CD13+ cells were closely associated with SP cells. They demonstrated that CD13 was a marker for semiquiescent CSCs in human liver cancer based on its predominant distribution during the G1/G0 phase. The CD13+ cells possessed a high tumorigenic potential in NOD/SCID mice and also induced chemoresistance to doxorubicin and 5-fluorouracil treatment. This result could be explained by the low levels of intracellular reactive oxygen species that protected the cells from DNA damage and induced apoptosis in the CD13+ fraction. Combining a CD13 inhibitor with regular chemotherapy may improve survival of patients with HCC [20]. However, because CD13 expression is not specific for HCC and is present in a variety of cells other than CSCs, caution should be exercised in treating HCC with CD13 as a CSC target [61,63].
Other candidate markers
Several reports have described additional possible LCSC markers. ALDH is a cytosolic enzyme including more than 17 human ALDH isoforms. ALDH activity is important for multiple biological activities including drug resistance, cell differentiation, and oxidative stress response [64]. Isolated lung and breast cancer cells with high ALDH1 activity display features of CSCs [65,66]. Ma et al. found that ALDH1A1 expression and activity positively correlated with CD133 expression in liver cell lines [19]. Subsequent functional studies revealed the CD133+/ALDH+ cells bearing higher tumorigenic potential than CD133+/ALDH− and CD133−/ALDH− counterparts, which confirmed that ALDH activity is a more specific marker for the CD133+ liver CSC population.
Oval cells are one of the important progenies of liver stem cells. OV6 is widely used as the oval cell marker of choice [67]. It is a mouse monoclonal antibody that recognizes a common epitope in CK14 and CK19 in rat liver [68]. Absent in normal human liver tissue, OV6 positivity is present in the ductular proliferative cells and periseptal hepatocytes in diseased liver [69].Yang et al. have demonstrated that OV6+ HCC cells possess greater tumorigenic ability and chemoresistance to standard chemotherapy when compared with OV6− cells [18]. In addition, the CD133+ population was found to be significantly enriched in cells positive for OV6, indicating that OV6+ is a potential LCSC marker. Another oval cell marker, c-Kit (CD117), is not significantly overexpressed in HCC [70]. The proportion was less than 0.1% of total tumor cell volume in most HCCs [71]. To date, there is not enough evidence supporting c-Kit or other oval cell markers to be accepted as an LCSC marker.
As a biomarker of HCC, AFP is positive in human hepatoblasts, fetal hepatocytes [72], and rat oval cells [73], but negative in mature hepatocytes and human hepatic stem cells [56]. As stated above, AFP could be used with EpCAM to isolate cells with stem cell features [17]. There are no studies supporting AFP as an independent LCSC marker.
LCSC Markers in Action
Until now, fluorescence-activated cell sorting and magnetic-activated cell sorting analyses have been frequently used to isolate CSCs. Definite LCSC markers are essential to identify and isolate these cells, which could enhance our understanding of the initiation, progression, metastasis, or relapse of HCC at molecular, genomic, or epigenetic level. In addition, LCSC markers may be predictors of clinical outcome and long-term survival.
As CSCs are difficult to treat by conventional methods because of their chemoresistant and radioresistant properties, specific targeting and eradication of CSCs is one of the most therapeutically important challenges. The cell surface LCSC markers CD133 and CD44 may be used as potential molecular therapeutic targets in HCC [14,74]. Future therapies are likely to be directed toward targeting CSCs by these LCSC markers or those pathways involved in CSC self-renewal. There are 2 major hypotheses regarding the cellular origin of cancer: from stem cells due to maturation arrest or from dedifferentiation of mature cells. The cellular origin of HCC is currently under debate. HCC may arise from cells at various stages of differentiation in the hepatic stem cells lineage (Fig. 1): (1) mature liver cells, which proliferate after normal liver tissue renewal and respond rapidly to liver injury; (2) liver progenitor cells or oval cells as bipotential stem cells, which are activated to proliferate when the liver damage is extensive and chronic; and (3) bone marrow stem cells, including HSCs and mesenchymal stem cells (MSCs), as multipotent liver stem cells. During liver regeneration and repair, replication of mature hepatocytes is the quickest and most efficient way to generate hepatocytes. Oval cells usually replicate and differentiate into hepatocytes only when replication of mature hepatocytes is delayed or entirely blocked. Bone marrow cells can generate hepatocytes in transplanted livers, but the frequency of hepatocytes produced by this route is very low [72]. According to the cell origin, present LCSC markers can be divided into different groups: (1) HSCs: CD133−; (2) MSCs: CD90, CD44; (3) oval cells: OV6, EpCAM; (4) others: ABCG2, CD13, ALDH. This classification helps to explore the properties of current LCSC markers and to find new markers.

Possible cellular origins and markers of LCSCs. HCC may arise from cells at various stages of differentiation in the hepatic stem cell's lineage: (1) mature liver cells; (2) liver progenitor cells or oval cells as bipotential stem cells; and (3) bone marrow stem cells, including HSCs and MSCs as multipotent liver stem cells. HCC could originate from stem cells because of maturation arrest or from dedifferentiation of mature cells. HSCs, hematopoietic stem cells; MSCs, mesenchymal stem cells; LCSCs, liver cancer stem cells; HCC, hepatocellular carcinoma.
Available evidence indicates that poorly differentiated HCC originates from bone marrow stem cells, whereas well-differentiated HCC originates from mature hepatocytes [75]. Thus, we can speculate on the characteristics of LCSCs from the origin of HCC, which can be divided into different subgroups using several LCSC markers. HCC with LCSCs originating from HSCs may have poor differentiation and higher malignant potential, whereas those derived from dedifferentiation of hepatocytes have the opposite property. A study of LCSC markers can help to identify the source of HCC.
To date, several markers have been reported for HCC. However, definite expression patterns of LCSC markers are still controversial. HCC is typically comprised of morphologically diverse cells that express various hepatic lineage markers. Therefore, there may be functionally diverse LCSC groups that express different markers. Although these LCSCs have similar “stem-like” characteristics, their differences are not clear. What is the relationship among various LCSC markers? Yang et al. found that only a proportion of CD45−/CD90+ cells concomitantly expressed CD133, whereas all the CD45−/CD133+ cells concomitantly expressed CD90, suggesting that CD45−/CD90+ was more sensitive than CD45−/CD133+ in identifying LCSCs [15]. Haraguchi et al. found that coexpression of CD133 or CD90 was detected in CD13+ HuH7 or CD13+ PLC/PRF5 cell lines, respectively [20]. Tomuleasa et al. also isolated hepatic cancer stem-like cells expressing CD133 and CD90 [76]. Yamashita and colleagues found that CD90 expression was limited to HCC cell lines that were EpCAM−/AFP−. However, this result has not been verified in other studies. Combinations of 2 or more markers may help to identify more invasive LCSCs, such as CD133+/CD44+ or CD90+/CD44+ (Fig. 2).
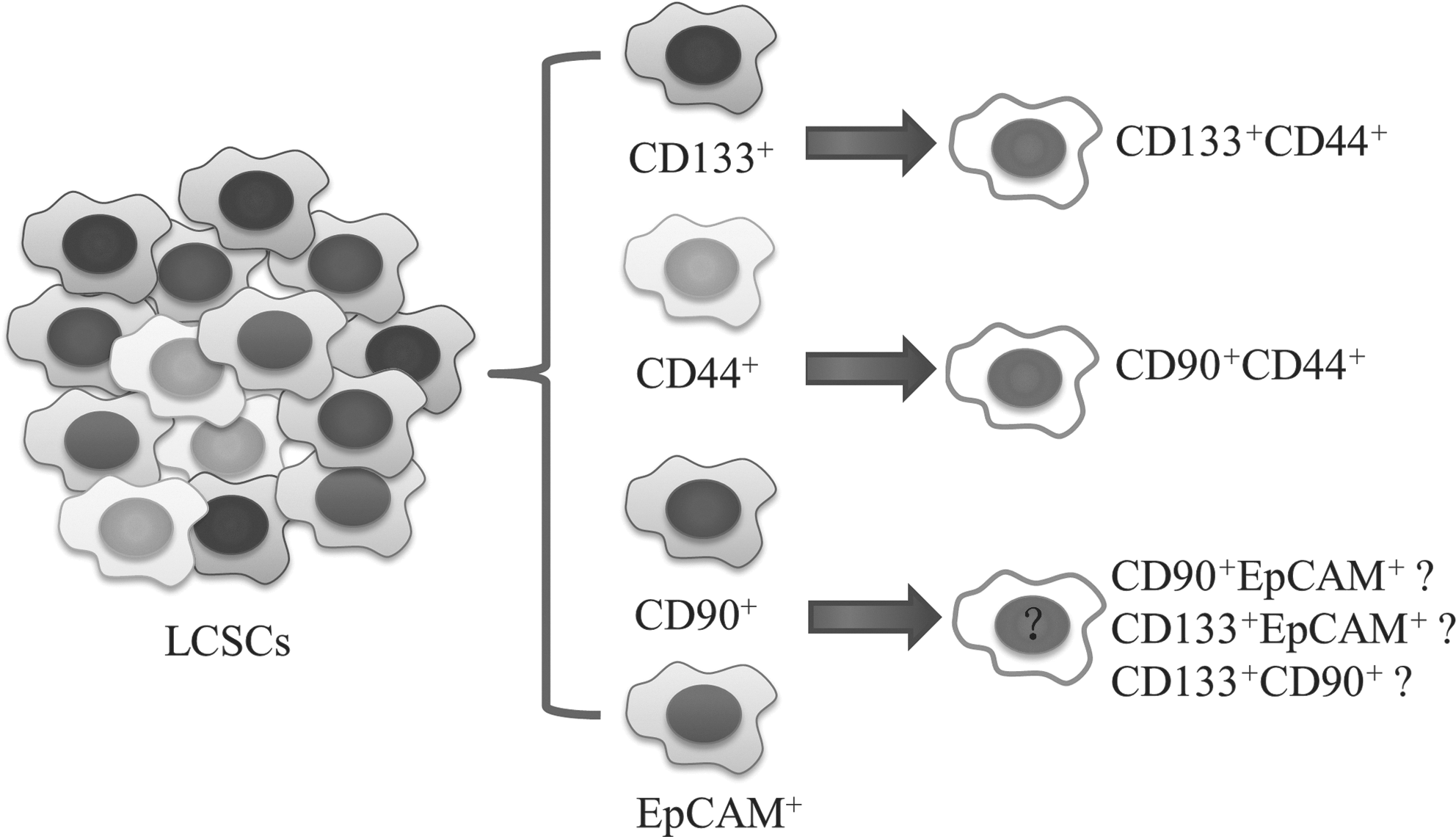
Possible relationships between various LCSC markers. Present research suggests the heterogeneity of LCSCs. Combinations of 2 or more markers may help to identify more invasive LCSCs such as CD133+/CD44+ or CD90+/CD44+. More studies are needed to define the coexpression pattern of different markers.
LCSC Markers as Prognostic Parameters
According to the CSC theory, CSCs may influence the prognosis of patients by promoting the recurrence, metastasis, and multidrug resistance of tumors. Recent findings suggest that the presence of CSCs could be linked with patients' survival [77,78]. However, the prognostic value of a single marker is still controversial. CD133+ and CD133− fractions of Huh7 cells did not differ in migratory properties [79]. EpCAM+/AFP+ HCCs had a significantly shorter survival, with a higher frequency of portal vein invasion, than EpCAM−/AFP− HCCs [16]. The number of LCSCs may impact malignant characteristics in HCC, such as tumor stage, histological grade, and AFP level, and therefore may be relevant to patients' prognosis. It is supposed that the greater the size of the CSC population, the poorer will be the prognosis. Present research has focused on positive or negative CSC markers in HCC. Few studies have been conducted on the size of CSC populations with positive markers. Song and coworkers first showed that increased CD133 expression was an independent prognostic factor in patients with HCC [78]. Further studies are still needed to determine the association between LCSC markers and pathologic features with the clinical outcome of HCC.
Because of the high degree of heterogeneity of HCC, the predictive range of a single marker is limited to a very small subpopulation. A combination of several LCSC markers may provide greater specificity and reliability. Yang et al. constructed a simplified predictive model using CD133, CD44, Nestin, and microvessel density (determined by CD34), which were found to be independent predictors of recurrence-free survival in multivariable analysis with clinicopathological characteristics [80].
Future Directions
The currently identified markers for LCSCs have their limitations. First, most investigations on CSCs or the SP phenotypes to date have been performed using cultured cell lines. Only Yang and colleagues have isolated CSCs from tissue and blood of patients with HCC. More studies are needed for validation of CSCs in tumor tissue.
Second, none of these markers is exclusively expressed by CSCs in HCC. They are often expressed by other stem/progenitor cell populations in patients. CD133, CD44, and CD90, as well as ALDH, are expressed in HCC tissue, suggesting stem/progenitor cell properties, which are also expressed in inflamed or nearly normal liver tissue [81]. In targeting cells that express these markers, there is the risk of depleting normal stem/progenitor cells simultaneously. More studies are needed to identify markers that are more specific or to use several markers in combination to differentiate CSCs from their differentiated counterparts and normal stem cells. Application of negative markers, such as CD45, may be helpful in isolating LCSCs.
Third, increasing evidence supports the notion that the presence and number of LCSCs are related to clinical outcome. However, the prognostic significance of LCSC markers remains controversial, especially over a long-term follow-up period. Thus, the study of the relationship between CSC markers and prognosis represents a promising new research direction.
Conclusions
There is increasing evidence to support the existence of liver CSCs. To date, LCSC can be identified by several markers, including CD133, CD90, CD44, OV6, and EpCAM, or by selecting for the SP cells. However, no single marker is exclusively expressed by CSCs in HCC. It is possible that various combinations of markers may be of value in identifying LCSCs and in determining prognoses of patients with HCC.
Footnotes
Acknowledgments
The authors thank members of Prof. Shen Xi-Zhong's laboratory for their helpful discussion and critical reading of the manuscript. This work was partially supported by the National Basic Research Program of China (2007CB936000), Ministry of Health (2009ZX10004-301), China National Key Projects for Infectious Diseases (2008ZX10002-017), National Nature Science Foundation of China (30872503), and China Postdoctoral Science Foundation–funded project (20100480542).
Author Disclosure Statement
The authors declare that there are no conflicts of interest.