
Abstract
The inhibition of endogenous differentiation-inducing signaling or the enhancement of growth capacity and viability of preimplantation embryos, via 2i (PD0325901 and CHIR99021), dramatically improves the establishment of mouse embryonic stem cells (mESCs). Using adrenocorticotropic hormone fragments 1–24 (ACTH 1–24), which enhances survival and/or proliferation of mESCs, also increases the derivation of mESCs from single blastomeres significantly. The CHIR99021 pathway and the proposed ACTH pathway are likely different. Therefore, this study aimed to assess the synergetic effects of 2i and ACTH 1–24 on derivation of mESCs. Results in the present study demonstrate that germline-transmitted mESCs could be efficiently derived from ICR and C57BL/6J at 0.5–4.5 days postcoitum denuded zygotes to blastocysts or isolated blastomeres of 2–8-cell embryos and cultured in 10 μL droplets with human foreskin fibroblast (Hs68) or STO (a mouse embryonic fibroblast line) feeders and in knockout serum replacement (KSR) ESC medium containing 2i or ACTH 1–24. The overall success rates for C57BL/6J and ICR were 56.2% when cultured in 2i+ACTH 1–24, 26.6% in 2i, 6.7% in ACTH 1–24, and 4.8% in KSR ESC medium. These results imply that CHIR99021 and ACTH 1–24 are synergistically enhancing the establishment of mESCs. The proposed protocol also demonstrates a highly efficient and reproducible method, has a simple layout, is easy to apply, and could be used as an alternative method for routinely establishing mESC lines.
Introduction
S
Signal transduction and regulatory mechanisms of self-renewal, differentiation, proliferation, and apoptosis [10,11] as well as corresponding inhibitors of stem cells [12 –15] have been investigated. Theoretically, inhibiting endogenous differentiation and/or enhancing proliferation of preimplantation embryos can be helpful in establishing ESC lines. Recently, using the mitogen-activated protein kinase (MAPK)/extracellular-signal-regulated kinase (ERK) kinase (MEK) inhibitor PD0325901 or SU5402 to eliminate differentiation-inducing signaling from MAPK and using the glycogen synthase kinase 3 (GSK3) inhibitor CHIR99021 to enhance ESC growth capacity and viability helped in establishing mESC lines [16]. Accordingly, ESC lines of the second mammalian species (rat) with germline transmission were reported [17,18].
It was considered that FBS contains growth factors that support ESCs. However, FBS also contains potential differentiation factors of ESCs [19,20]. To circumvent interference from differentiation factors, chemically defined knockout serum replacement (KSR) was developed for use in place of FBS [21]. However, KSR did not support mES single-cell culture. In contrast, mES single cells were propagated without loss of pluripotency when adrenocorticotropic hormone (ACTH) was added to KSR ESC medium [20]. The authors proposed that ACTH signaling for mESC survival and/or proliferation might be via inhibition of adenylyl cyclase activity. Later, a simple and efficient establishment method of mESC lines from a single blastomere of 2–8-cell embryos with KSR ESC medium containing ACTH fragments 1–24 (ACTH 1–24) on mEF feeders was reported [22].
The CHIR99021 pathway and the proposed ACTH pathway are likely different. Therefore, this study aimed to assess the synergetic effects of 2i (PD0325901+CHIR99021) and ACTH 1–24 on derivation of mESCs.
It has been speculated that feeder (helper) cells, in addition to their physical structure, have other positive effects for ESCs [23]. Conventionally, mEF or STO is the most popular feeder cells for deriving mESCs. However, human foreskin fibroblasts (hFFs) feeder cells can support propagation and self-renewal of human ESCs (hESCs) [24,25] and mESCs [26]. Further, hFF feeder cells can help establish hESCs for long-term culturing [27,28]. Reports demonstrated that hFF, mEF, and STO feeder cells secrete different growth factors in supporting ESCs [26,29]. Therefore, this study aimed to assess the effects of hFF on derivation of mESCs.
Mouse teratocarcinoma stem cells and ESCs are able to condition themselves. Therefore, a conditioned medium was used to help establish mESCs [2] and propagate them in high densities [3,30]. As a part of this reason, the different small growth area in multiwell plates was adopted to initiate cultivation, to establish ESCs. To decrease the volume of expensive media, for ease of handling, and to increase disaggregation efficiency of 3-dimensional growing outgrowths or cell colonies of ICMs and blastomeres, we hypothesize that home-made microdroplets on tissue culture dishes coated with feeder cells are as effective as wells of plates. This study aimed to test this hypothesis.
This study used ICR and C57BL/6J denuded intact zygotes, preimplantation embryos, or isolated blastomeres cultured in 10 μL droplets with hFF feeder cells on a chemically defined KSR ESC medium supplemented with differentiation inhibitors (2i: 0.5 μM PD0325901+3 μM CHIR99021) or survival and/or proliferation stimulator (10 μM ACTH 1–24) to evaluate the efficiency in deriving mESCs.
Materials and Methods
All embryo-tested or cell-culture-tested chemicals were purchased from Gibco (Invitrogen) unless stated otherwise.
Mice, embryos, and blastomeres
The treatment of experimental mice in this study, including feeding, management, treatment, anesthesia, and surgery, was in accordance with “A Guidebook for the Care and Use of Laboratory Animals” (The Chinese Association of Laboratory Animal Science, 2004). The Institutional Animal Care and Use Committee of the Animal Technology Institute Taiwan approved all procedures. The lighting period (05:00–19:00) and temperature (24°C±2°C) in the clean conventional mouse room were controlled automatically.
Sexually mature ICR and C57BL/6J females were intraperitoneally injected with 10 U of pregnant mare serum gonadotrophin followed by 10 U of human chorionic gonadotrophin injected 40–48 h later at approximately either 09:30 or 16:00. The females were then paired with stud ICR or C57BL/6J males, respectively. Each vaginal plug was checked the next morning [0.5 day postcoitum (dpc)]. The 0.5–4.5-dpc embryos were flushed from oviducts and/or uterine horns with KSOM medium supplemented with 20.85 mM HEPES (HK).
The 1-cell zygotes to hatched blastocysts were washed and placed in HK at room temperature for ∼15 min until the next treatment. Zona pellucida of the embryos was removed (denuded, naked) in seconds using the acidic Tyrode's solution. The blastomeres of 2–8-cell embryos were separated by incubating denuded embryos in 0.25% trypsin–EDTA for ∼4 min at 37°C in a 5% CO2 incubator, followed by gentle pipetting. The isolated blastomeres and denuded embryos were washed with KSR ESC medium and then plated to feeders in different culture media. Single blastomeres that originated from 2- and 8-cell embryos were designated as 1/2 and 1/8, respectively.
Establishment and maintenance of mESCs
The STO and Hs68 (hFF; ATCC No. CRL-1635; BCRC No. 60038) medium was DMEM (glucose, 4.5 g/L; Sigma) containing 10% FBS (standard; HyClone), penicillin (50 U/mL), and streptomycin (50 μg/mL). After 10 μg/mL mitomycin C-inactivated cultivation for roughly 3 h, single trypsinized cells were frozen until thawed for feeder preparation. Hatched blastocysts and denuded embryos, zygotes, or blastomeres were cultured individually on the STO or Hs68 feeder layers seeded in 10 μL droplets (not gelatin-coated), under paraffin liquid (Code 26137-85; Nacalai Tesque, Inc.), on 60-mm tissue culture dishes (thirty-two 10-μL droplets) at 1 or 2 days before embryos or blastomeres were plated.
The KSR ESC medium (control medium) contained knockout DMEM (glucose 4.5 g/L) supplemented with 20% KSR, 0.1 mM nonessential amino acids, 1.75 mM GlutaMAX™-I supplement, 0.1 mM β-mercaptoethanol, penicillin (15.62 U/mL), streptomycin (15.62 μg/mL), and home-made LIF (∼0.3%, v/v) roughly 1,000 U/mL [31]. To evaluate the derivation of mESC lines, denuded embryos and blastomeres were cultured in control KSR ESC medium or supplemented with 0.5 μM PD0325901 (MEK inhibitor; Stemgent, Inc.), 3 μM CHIR99021 (GSK3 inhibitor; Stemgent) (2i) [16], and/or 10 μM ACTH 1–24 (adenylyl cyclase inhibitor; American Peptide Co., Inc.) [22].
Approximately 2 h before denuded embryos and blastomeres were seeded on feeders, fresh ESC media were exchanged. Embryos and blastomeres were cultured in humidified atmosphere comprising 5% CO2 in air in a 37°C incubator. Following attachment to the feeder, the media were exchanged every 2–3 days. After the initiation of cultivation for 2–4 days, the developmental patterns of embryos and blastomeres were recorded. After culturing for 5±1 days (intact embryos) or 9±2 days (blastomeres and zygotes), individual 3-dimensional outgrowth or cell colony was identified and then feeder cells and flat-growing cells were mechanically removed via a mouth pipette. The outgrowth or cell colony, in the same droplet, was washed with TrypLE Express once and then incubated with TrypLE Express in a 37°C incubator for ∼9 min. The outgrowth or cell colony was washed with KSR ESC medium once and then disaggregated into clumps and single cells, which were reseeded onto fresh 20 μL droplets [twenty-four 20 μL droplets in one 60-mm dish; passage No. 1 (P1)] with corresponding feeders and media. After 5±2 days, morphologically ES-like colonies were then passed (via TrypLE Express) to fresh 20 μL droplets or 4-well plates (P2) and thereafter either 4-well plates or 35-mm cell culture dishes (P3) depending on the number of cells. Once ES-like cells were growing in 4-well plates, KSR ESC medium was used for all subsequent propagation. At passages 5±2, mES-like cells were frozen in FBS supplemented with 10% (v/v) dimethyl sulfoxide.
Chimeric embryos generated via ESCs microinjected into 2.5-dpc embryos in 0.2 M sucrose hypertonic medium
Chimeric embryos were generated via hypertonic microinjecting of ESCs to embryos, a novel developing method at our laboratory. The double-plating protocol to enrich mESCs for microinjection has been previously described [32]. The pellet of enriched, thawed or fresh C57BL/6J single mESCs was resuspended in ∼5 μL of 0.2 M sucrose EHK (37.5% KSR ESC medium, 62.5% HK mixture, and 285±10 mOsm/kg H2O) and stored at 4°C until microinjection. Thousands of enriched single mESCs (B6) and dozens of ICR×ICR 2.5-dpc 3–8-cell embryos were transferred to the same droplet of 0.2 M sucrose EHK (500±10 mOsm/kg H2O) medium for hypertonic microinjection. After thousands of mESCs were sucked into the injection pipette (outer diameter, 17–20 μm; inner diameter, 13–16 μm) in minutes, ∼10–20 mESCs were injected at room temperature into the large subzonal cavity of 3–8-cell embryos. After 50–70 embryos were injected within 1 h, the injected embryos were washed with droplets containing 37.5% KSR ESC medium and 62.5% KSOM-AA (EK) for ∼1 h and then washed with fresh EK or KSOM-AA for culturing overnight until transfer to uterine horns of 2.5-dpc or oviducts of 0.5-dpc pseudopregnant ICR recipients. Natural birth followed. The black and white coat color of pups was checked for evidence of chimerism. Chimeras confirmed that mES-like cells were mESCs. Phenotypically normal male chimeras were naturally mated with female ICR. The coat color of pups revealed the germline transmission.
Observation and record of images
A Zeiss Axiovert 35 microscope fitted with Nikon D1 digital camera body was used for photographing embryos and cells. For pups, a Nikon D1 camera body with an AF MICRO NIKKOR (55 mm, 1:2.8) lens was used to take color images. Adjustments of brightness and contrast applied to the entire digital images were processed using the Photoshop 7.0 software (Adobe Systems).
Statistical analysis
All data were subjected to an analysis of variance using the General Linear Model procedure in SAS version 9.2 (SAS Institute) followed by the Student–Newman–Keuls test. Percentile data were arcsine square root transformed before analysis of variance. A P value of <0.01 was considered statistically significant. Each treatment included replicates in all experiments.
Results
STO or Hs68 feeders on 10 μL droplets supported efficient derivation of mESCs using KSR ESC medium containing differentiation inhibitors (2i)
The first experiment tested the efficacy of deriving mESCs with 2i and STO or Hs68 feeders on 10 μL droplets. ICR and C57BL/6J 1.5–4.5-dpc 2-cell embryos to blastocysts and blastomeres isolated from 2- to 8-cell embryos were plated on STO or Hs68 feeders in KSR ESC medium supplemented with or without 2i (Figs. 1 and 2). Experimental results demonstrated that the 2i and embryo status (denuded intact embryo vs. blastomere) affected the establishment of ESC line (P<0.01; Table 1). Results indicated that the KSR ESC medium containing 2i significantly increased derivation efficiency for mESCs in denuded intact embryos from 10.8%±5.8% to 40.8%±7.6% (mean±SEM) regardless of mouse strains and feeders (P<0.01). For blastomeres, it was 1.1%±1.1% to 14.0%±4.8% (P<0.01; Table 1). No difference existed between STO and Hs68 feeders in the establishment of mESCs (P>0.01; Table 1). However, as the STO feeders occasionally detached and curled up during cultivation, Hs68 feeders were adopted for subsequent experiments. Results also confirmed that the 10 and 20 μL (P1–P2) droplets and 4-well plates (P2–P3) sequential cultivation was very easy to handle and reduced the need of expensive media to efficiently establish mESCs.
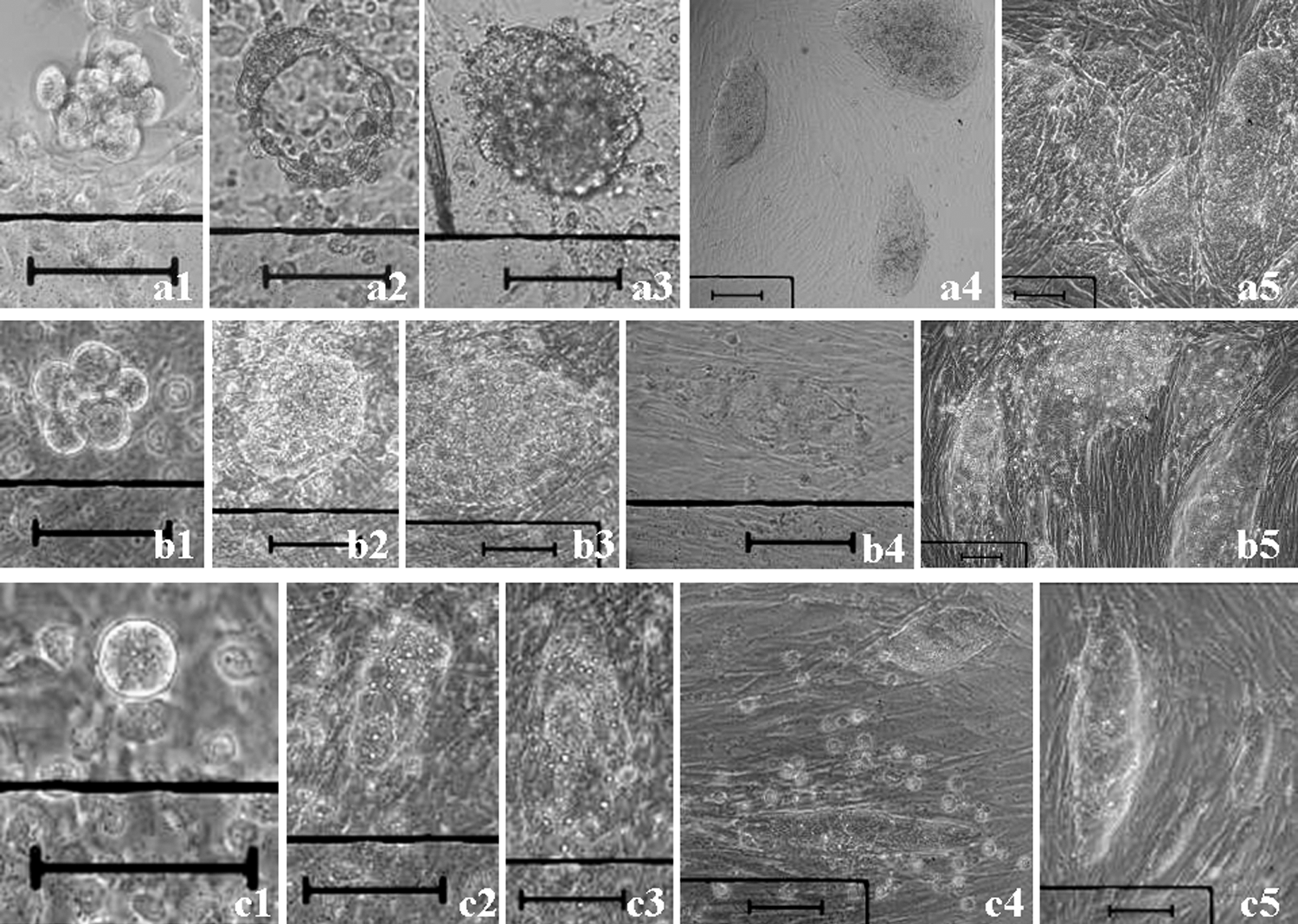
C57BL/6J embryonic stem cells (ESCs) derived from 2.5-day postcoitum (dpc) denuded 8-cell embryos or single blastomeres cultured in 10 μL droplets.
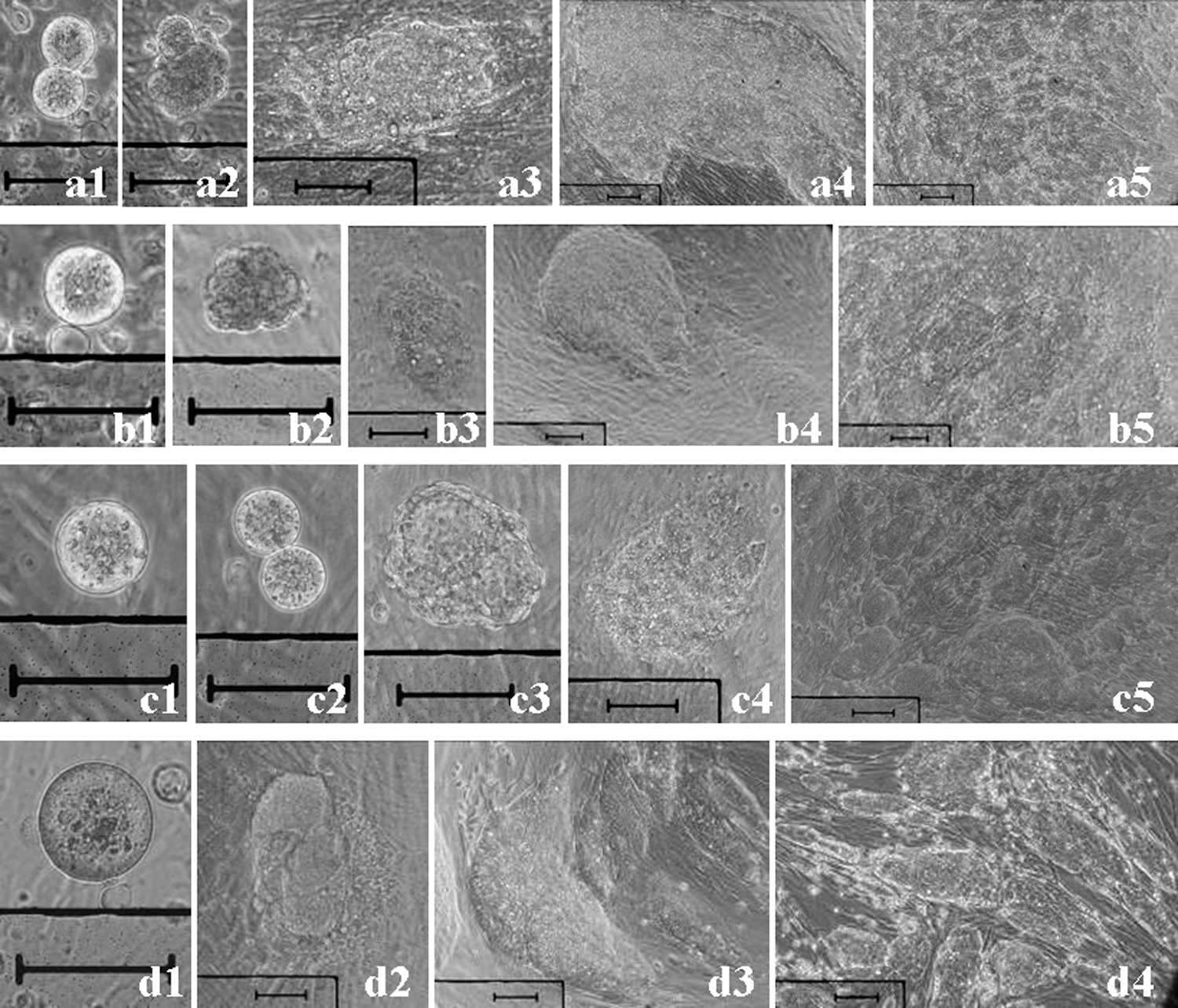
C57BL/6J ESCs derived from 1.5-dpc denuded 2-cell embryos or isolated single blastomeres and 0.5-dpc denuded zygotes cultured in 10 μL droplets with Hs68 feeders.
Blastomeres isolated from 2- to 8-cell embryos of 1.5–2.5-dpc ICR×ICR and C57BL/6J (B6)×C57BL/6J were used.
Two-cell embryos to blastocysts from 1.5- to 4.5-dpc ICR×ICR and B6×B6 were used.
KSR ESC medium: DMEM containing 20% KSR and LIF without fetal bovine serum.
2i: 0.5 μM PD0325901 (MEK inhibitor)+3 μM CHIR99021 (GSK3 inhibitor).
Embryo status (denuded intact embryo vs. blastomere) and c,d2i affect the establishment of ESC line significantly (P<0.01).
dpc, day postcoitum; ESC, embryonic stem cells; GSK3, glycogen synthase kinase 3; KSR, knockout serum replacement; LIF, leukemia inhibitory factor; MEK, mitogen-activated protein kinase/extracellular-signal-regulated kinase kinase.
Differentiation inhibitors or proliferation stimulators significantly increased derivation efficiency for mESC lines
To establish mESCs, experimental results showed that KSR ESC medium supplemented with 2i and ACTH 1–24 (2i+ACTH 1–24) was significantly superior to medium containing only 2i, which was in turn better than KSR ESC medium supplemented with only ACTH 1–24. The KSR ESC medium was the poorest for deriving mESCs. The overall successful efficiency for C57BL/6J and ICR was 56.2%±11.8% (2i+ACTH 1–24), 26.6%±5.5% (2i), 6.7%±4.4% (ACTH 1–24), and 4.8%±3.6% (KSR ESC medium) (P<0.01; Table 2). For C57BL/6J, it was 48.8%±10.8% (2i+ACTH 1–24), 21.6%±6.2% (2i), 11.9%±7.5% (ACTH 1–24), and 2.2%±2.2% (KSR ESC medium) (P<0.01; Supplementary Table S1; Supplementary Data are available online at
Zygotes and 2-cell embryos to blastocysts from 0.5- to 4.5-dpc ICR×ICR and C57BL/6J × C57BL/6J were used. 1/2: One blastomere isolated from 2-cell embryos (2/2); 1/8: one blastomere isolated from 8-cell embryos (8/8).
2i+ACTH 1–24: 0.5 μM PD0325901 (MEK inhibitor)+3 μM CHIR99021 (GSK3 inhibitor)+10 μM ACTH fragments 1–24 (survival and/or proliferation stimulator).
One B6 ESC line derived from four 1/8 in KSR ESC media without LIF.
Two B6 ESC lines derived from 2 morulae in KSR ESC media without LIF but supplemented with 2i, and 1 ESC line derived from 1 denuded morula treated with acidic Tyrode's solution for 30 s.
Three B6 ESC lines derived from 4 denuded blastocysts treated with acidic Tyrode's solution for 30–40 s.
Culture media with different letters differ significantly (P<0.01).
Embryo status with different letters differ significantly (P<0.01).
ACTH, adrenocorticotropic hormone.
Further, this is the first study demonstrating that zygotes can be used to establish mESCs (Table 2; Fig. 2d). Notably, this study found that mESCs were established at very high rates from denuded morula and blastocysts treated with acidic Tyrode's solution for 30–40 s to denature or partially damage cells on the surface of embryos (Table 2). Results also showed that mESCs could be established in media without LIF (Table 2).
The relationship between embryo, blastomere developmental patterns and successful establishment of mESCs
The 2i or ACTH 1–24 could not completely inhibit differentiation of embryos and blastomeres to develop into typical blastocysts or small blastocysts that have a cavity with or without visible ICM-like cells (Table 3 and Figs. 1a and 2c). The developmental patterns of embryos and blastomeres following cultivation were not affected by treatments (inhibitor × stimulator), but were affected by embryo status (P<0.01) except those embryos developed to cell colonies directly (Table 3). However, blastomeres tend to have a higher proportion developed to small blastocysts, whereas denuded intact embryos tend to have a higher proportion developed to typical blastocysts or no development at all (Table 3). The embryos or blastomeres directly developed to cell colonies were significantly affected by embryo status, embryo development stage, and treatments (P<0.01; Table 3). Those embryos or blastomeres developed to cell colonies directly and typical blastocysts have a higher proportion of mESCs established than embryos or blastomeres developed to small blastocysts or no development (P<0.01; Table 4). Addition of both inhibitor and stimulator resulted in a higher proportion of ESCs established than those without inhibitor and stimulator (P<0.01; Table 4).
Blastomeres isolated from 2- to 8-cell embryos of 1.5–2.5-dpc ICR × ICR and C57BL/6J (B6) × C57BL/6J were used. Two-cell embryos to blastocysts from 1.5- to 4.5-dpc ICR × ICR and B6 × B6 were used.
After the initiation of cultivation for 2–4 days, the developmental patterns of embryos and blastomeres were recorded.
Developing small blastocysts show cavity and with or without visible ICM-like cells.
The developmental patterns of embryos and blastomeres following cultivation were not affected by treatments (inhibitor×stimulator), but were affected by embryo status (P<0.01) except those embryos developed to cell colonies directly. Blastomeres tend to have a higher proportion developed to small blastocysts, whereas denuded intact embryos tend to have a higher proportion developed to typical blastocysts or no development at all.
ICM, inner cell mass.
The embryos or blastomeres developed to cell colonies directly and typical blastocysts have a higher proportion of ESCs established than the embryos or blastomeres developed to small blastocysts or no development (P<0.01). Addition of both inhibitor and stimulator resulted in a higher proportion of ESCs established than those without inhibitor and stimulator (P<0.01).
Blastomeres isolated from 2- to 8-cell embryos of 1.5–2.5-dpc ICR×ICR and C57BL/6J (B6) × C57BL/6J were used. Two-cell embryos to blastocysts from 1.5- to 4.5-dpc ICR × ICR and B6 × B6 were used.
Developing small blastocysts show cavity and with or without visible ICM-like cells.
Germline-transmitted chimeric mice
After ESCs were hypertonic microinjected into 2.5-dpc embryos and cultured overnight, embryos were transferred to oviducts or uterine horns. After natural birth, pups born alive showed the high level of black and white coat color chimerism (Figs. 3a–c). After backcross breeding, coat color of pups demonstrated the germline transmission (Figs. 3d, e).
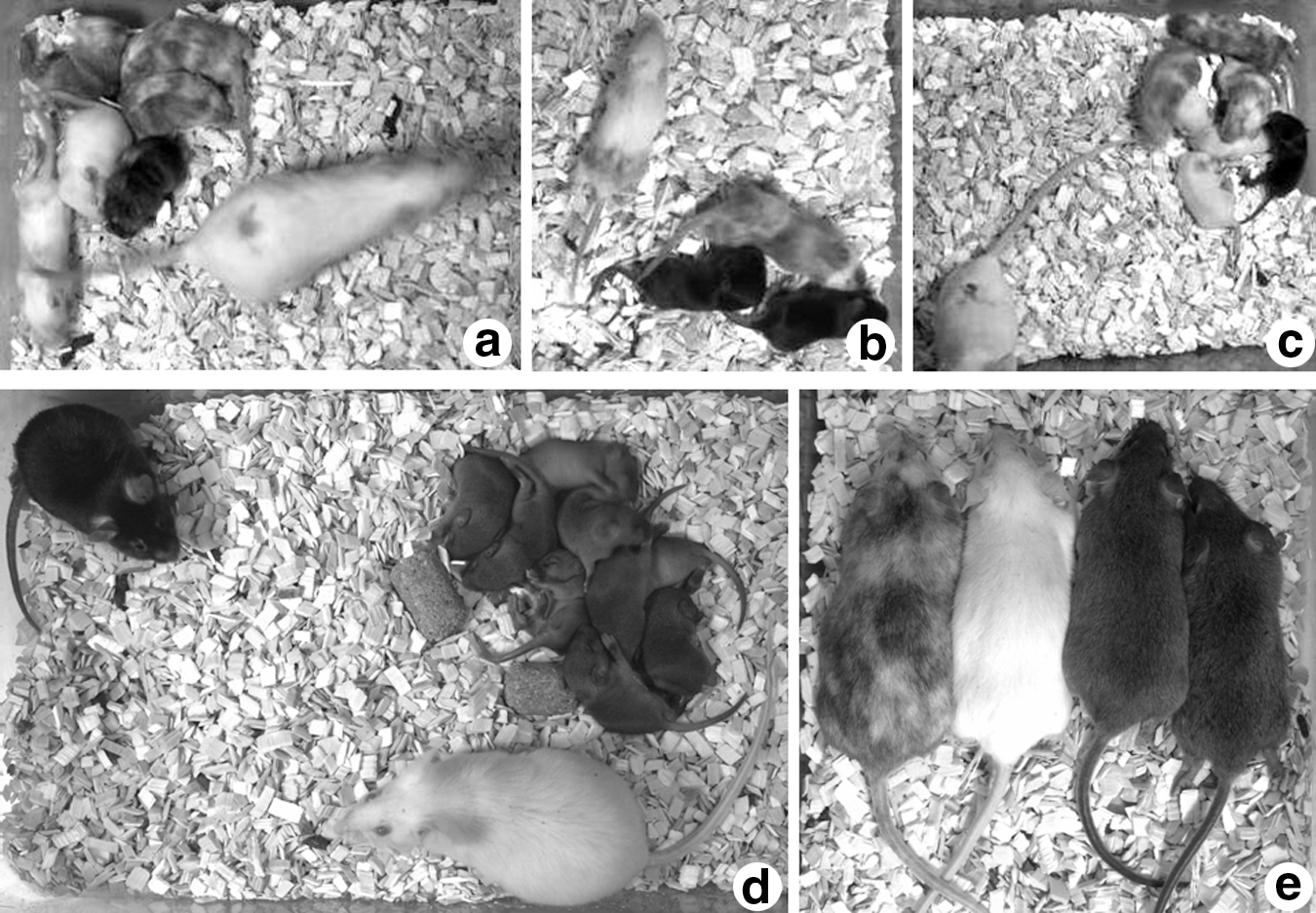
Chimeric mice with germline transmission generated via hypertonic microinjection. The C57BL/6J ESCs in 0.2 M sucrose EHK were microinjected into ICR × ICR 2.5-dpc embryos and washed into EK or KSOM-AA cultured overnight. Injected embryos were transferred to the pseudopregnant ICR 2.5-dpc uterine horns. The ESC98B33
Discussion
In the present study, neither STO nor Hs68 feeder cells coated on 10 μL droplets and cultured in KSR ESC medium supported derivation of ESCs from embryos or blastomeres of ICR or C57BL/6J effectively. However, by supplementing KSR ESC medium with 2i or ACTH 1–24, the efficiency in establishing mESC lines increased dramatically. Results also indicated that to inhibit cell differentiation and increase cell growth/viability (in the presence of 2i) simultaneously tends to be better than only increasing cell survival and/or proliferation (with ACTH 1–24) when deriving mESCs. In addition, to our knowledge, this is the first report of ESCs derived from zygotes. These zygote-derived mESCs are morphologically indistinguishable from mESCs derived from fertilized embryos and blastomeres. Moreover, the generation of germline-transmitted chimeras confirmed that the established mES-like cells are mESCs.
Theoretically, inhibiting differentiation and/or enhancing proliferation of preimplantation embryos can be helpful in establishing ESC lines. Serum and serum substitutes contain various inductive stimuli that may activate commitment and differentiation programs [17]. Therefore, simple withdrawal of serum or other exogenous stimuli cannot prevent differentiation of ESCs because of endogenous autoinductive differentiation of fibroblast growth factor 4 via the MEK/ERK pathway [16,33]. However, to suppress differentiation sufficiently and to maintain high viability and growth rates, one must provide LIF or restrict the activity of GSK3, which acts mainly via the Wnt/β-catenin signaling pathway to suppress cellular biosynthetic capacity and then cell growth and viability [34,35]. Recently, using the MEK inhibitor PD0325901 or SU5402 to eliminate differentiation-inducing signaling from MAPK and using the GSK3 inhibitor CHIR99021 to enhance ESC growth capacity and viability enables derivation and propagation of germline-competent mESCs from CBA and 129 strains in a chemically defined medium (N2B27) [16]. Accordingly, ESC lines of the second mammalian species (rat) with germline transmission have been reported [17,18]. However, the efficiency was not reported [16]. Experimental results obtained in this study showed that the efficiency in establishing C57BL/6J and ICR ESCs in KSR medium containing 2i could be as high as 66.7% for morulae and 54.0% for blastocysts. The efficacy of 2i to help establish mESCs is confirmed.
A previous report showed that KSR ESC medium did not support the mES single-cell culture, likely because this medium lacked some important growth factors or factors such as “stem-cell autocrine factor” are secreted by mESCs themselves [20]. However, when the KSR ESC medium was supplemented with ACTH, single mESCs were propagated and their pluripotency was maintained. The authors hypothesized that ACTH may be integrated via a weak cross interaction with an unknown, nonphysiological inhibitory G protein-coupled receptor. The signaling system other than the cAMP-PKA pathway or PKA may play an important role in propagation of mESCs [20]. Accordingly, mESC lines have been efficiently established using single blastomeres from 2- to 8-cell embryos in KSR ESC medium containing ACTH 1–24 [22]. In the present study, results suggest that the GSK3 inhibitor (CHIR99021) and ACTH 1–24 likely have different pathways in synergistically enhancing the establishment of mESCs, because 2i+ACTH 1–24 are much better than 2i or ACTH 1–24 alone.
In 1 trial in this study, 2 C57BL/6J ESCs were derived from 2 morulae on Hs68 feeder cells with KSR ESC medium supplemented with 2i but without exogenous LIF. A previous study showed that the combination of PD98059 (an inhibitor of MEK) with LIF enhances derivation of ESC lines from the refractory CBA strain [36]. Combining the MAPK-activated protein kinase 1/2 (MEK1/2) inhibitor U0126 [37] or GSK3 inhibitor 6-bromoindirubin-3′-oxime (BIO) [35] with LIF significantly increased the efficiency of mESC isolation of strains other than the 129 strains. Previous reports indicated that the inhibitors that block the MEK/ERK differentiation pathway can support self-renewal of mESCs even without LIF signaling [12,16]. However, these small molecules only play positive roles in mESC self-renewal, and cannot maintain the pluripotency of mESCs during long-term culturing without LIF [16,38]. Our results revealed that mESCs could be derived using feeder cells and 2i but LIF is option.
In the present study, results showed that mESC establishment rates were 0%, 12.5%, 8.3%, and 50.0% for blastomeres derived from C57BL/6J 2-cell embryos cultured in KSR ESC medium, 2i, ACTH 1–24, and 2i+ACTH 1–24, respectively (Supplementary Table S1). To date, most available ESC lines were derived from outgrowth of ICM of blastocysts. However, because of ethical concerns on the derivation of hESCs for regenerative medicine, a single blastomere (usually from 8-cell embryos) has been utilized to generate ESCs. In 1996, mES-like cells were first reported from single blastomeres [39]. Since then, no ESCs were established from blastomeres until 2006 [40]. Although overall successful rates were <5% [39 –41], it suggests the possibility of deriving personalized hESCs.
Wakayama et al. developed a protocol and achieved the best results to date for establishing mESCs via blastomeres. In their experiment, the zona pellucida was removed with acidic Tyrode's solution, and naked B6D2F2 embryos were pipetted several times to separate blastomeres. After culturing on KSR (20%) ESC medium containing ACTH 1–24 in 96-well plates coated with mEF, the mESC establishment rates were 33%, 8%, and 8% for blastomeres derived from 2-cell, late 4-cell, and 8-cell embryos, respectively [22]. The authors did not aggregate blastomeres with other mESCs, suggesting that the success was likely attributable to the KSR ESC medium containing ACTH 1–24 [22]. Recently, Lorthongpanich et al. demonstrated that CD-1 ESC lines can be established from 1/2 and 1/4 single blastomere-derived blastocysts in ACTH 1–24 [42]. Both studies demonstrated that a single blastomere does not need to aggregate with a small clump of mESCs at the beginning of cultivation to establish mESCs [40]. These experimental results suggest that positive and synergetic effects of 2i and ACTH 1–24 could be useful reagents in supporting ESC derivation from blastomeres.
A previous study demonstrated that blastomeres from 2- to 8-cell embryos developed to blastocysts in 3–5 days in KSR ESC medium containing ACTH 1–24 [22]. Another report also indicated that 2–4-cell embryos and blastomeres, which developed into blastocysts in medium containing ACTH 1–24, PD98059 (MAP2K1 inhibitor), or SB203580 (MAPK14 inhibitor), yielded developmental rates comparable to those of the control [43]. This study showed that ∼60% of 1.5–2.5-dpc denuded intact embryos developed into typical blastocysts or small blastocysts and ∼80% of corresponding blastomeres developed into small blastocysts in 2–4 days in KSR ESC medium containing 2i or ACTH 1–24. The results implied that 2i and ACTH 1–24 had little or no adverse effect on cell proliferation and development of embryos and blastomeres. Further, these observations indicate that 2i, MAP2K1, and MAPK14 inhibitors are not strong enough to inhibit differentiation of embryos and blastomeres entirely. We also observed that 0.3–3 μM pluripotin (SC-1; inhibitor of RasGAP and ERK1 pathways) [12] did not inhibit differentiation of ICR 2.5-dpc embryos and blastomeres completely (unpublished data). All these experimental findings suggest that other more powerful inhibitors via RAS-MEK-ERK or other pathways may be worth developing to further enhance the success rate of deriving ESCs. Theoretically, cocktail chemicals that can completely inhibit differentiation, increase propagation, and decrease apoptosis of pluripotent cells should maximize the derivation of ESCs.
Feeder cells support embryos and ESC attachment through the physical matrix. Further, they may release embryo trophic factors, reduce inhibitory or toxic factors in FBS, may be beneficial by lowering concentrations of ions and/or glucose in medium and overcoming developmental blockage of embryos mediated by the release of growth factors essential for activation of the embryonic genome and for normal embryonic development, or may protect embryos from oxygen toxicity [23]. Recently, reports indicate that they also enhance the survival and propagation of ESCs through the production of growth factors [26,29,44]. Various feeder cells, which differ in their capability to support ESCs, have been utilized for the establishment, propagation, and maintenance of ESCs of pluripotency. Recently, hFFs showed excellent traits that support hESCs and mESCs [25,26,45]. In the present study, we demonstrated that the hFF (Hs68) feeder was as good as the mouse STO feeder for derivation of ICR and C57BL/6J ESCs.
Experimental data obtained by the present study showed that C57BL/6J blastocysts in KSR ESC medium did not lead to the derivation of ESCs effectively; however, the success rate in 2i medium was as high as 61%. Further, 8-cell embryos (38%) also demonstrated the efficacy of 2i in efficiently deriving mESCs. Generally, the efficiency in establishing C57BL/6 (B6) ESCs via FBS or KSR ESC medium containing LIF varies and is typically <10% [9,19,46 –55]. When establishing C57BL/6J ESC lines using FBS (15%) ESC medium containing LIF, the highest success rate was 40% (36/89) [52]. Notably, this study used mEF feeder cells coated on 96-well plates for initiation of blastocysts and thereafter digested ICM outgrowths cultured at 39°C under 5% CO2 in an incubator. In establishing B6 ESCs using KSR (20%) ESC medium containing LIF, the rate of (C57BL/6N × C57BL/6J) F1 ES-like colony generation can be as high as 40% (10 of 25 blastocysts) [55]. Previous results also demonstrated that the efficiency of establishing ESCs was significantly higher for the C57BL/6N strain (53%) than for the C57BL/6J strain (20%) [55]. Our results demonstrated that C57BL/6J denuded intact embryos cultured in 10 μL droplets with Hs68 feeders and in KSR ESC medium supplemented with differentiation inhibitors (2i) is efficient to establish mESCs.
Practically, the first 1–3 passages are critical for successful derivation of mESCs. This study only used KSR ESC medium containing 2i or ACTH 1–24 for the first 10 μL droplets (for embryo outgrowth) and second (for single cells and clumps of digested outgrowths) and/or third 20 μL (for ES-like cells) droplets (P1–P2). Once ES-like cells started to grow on 4-well plates (P2–P3), KSR ESC medium was used for all following treatments. The feeders, LIF, and the high density of ESCs may explain these promising experimental results. By this simple protocol, the success rates in establishing B6 ESCs are comparable to [52,55] or much better than other studies [9,19,46 –51,53,54].
The capability for ESC incorporation into developing embryos and chimeras delivery is an important trait of authentic ESCs. In this study, chimeras were generated via hypertonic microinjection of mESCs into subzonal cavity of 2.5-dpc embryos (manuscript in preparation). Germline-transmitted chimeras demonstrated that established ES-like cells are mESCs.
To summarize, this study shows that ESC lines could be efficiently derived from ICR and C57BL/6J 0.5–4.5-dpc denuded intact zygotes, embryos, or isolated blastomeres from 2- to 8-cell embryos and cultured in 10 μL droplets with hFF or STO feeders and in a chemically defined KSR ESC medium supplemented with differentiation inhibitors (2i) and/or proliferation stimulator (ACTH 1–24). The proposed protocol has a simple layout, is easy to operate, is highly efficient, is reproducible, and could be an alternative method for establishing ESC lines routinely.
Footnotes
Acknowledgments
The authors thank the National Science Council of the Republic of China, Taiwan, for financially supporting this research under Contract Nos. NSC99-2324-B-059-001, NSC97-2317-B-059-004, and NSC95-2317-B-059-001. H.R. Chang, T.L. Hsu, and M.S. Liu at Animal Technology Institute Taiwan (ATIT) are thanked for their technical assistance and routine maintenance in the laboratory and mouse facility. The authors thank Prof. Kong-Bung Choo, Taipei Veterans General Hospital, for providing comments on the manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.