
Abstract
Mesenchymal stem cells (MSCs) can be mobilized from the bone marrow and enter the circulation. Conversely, MSCs can be recruited from the circulation and into the bone marrow. For these migratory pathways, MSCs have to traverse the bone marrow endothelium, in a basal-to-apical and apical-to-basal direction, respectively. Here we describe the migratory cues that drive MSC transendothelial migration in both directions with focus on chemokines. Live cell imaging and electron microscopy were used to examine the interaction of human MSCs with human bone marrow endothelial cells (HBMECs), and MSC transmigration analyzed. Chemokines CXCL12, CXCL13, CXCL16, CCL11, and CCL22 significantly enhanced transendothelial migration in an apical-to-basal and basal-to-apical direction, showing preferences in terms of their capacity to stimulate the direction of migration. For apical-to-basal migration CXCL16 was the most effective (6-fold stimulation), with the rank order being CXCL16>CCL11>CXCL13>CCL22>CXCL12. In the basal-to-apical direction CCL22 was the most effective (5-fold enhancement), with the remaining chemokines being roughly equal. When MSCs interacted with HBMECs they flattened, extended long microvilli (filopodia) and podosome-like protrusions that inserted into the endothelial cells. In conclusion, chemokines enhance the migration of MSCs bidirectionally across HBMECs, with directional preferences shown for different chemokines.
Introduction
M
The role of adult MSCs is to help maintain a status quo in the body, that is, to replace degenerated tissue with healthy new tissue. This is why they are multipotent cells that differentiate into the required tissue-specific cells when provided with the correct stimulus and why they are found in many adult human tissues. They also produce important hematopoietic cytokines to support hematopoiesis [11]. Since the 1860s when the German pathologist Julius Cohnheim first suggested that MSCs originated in the bone marrow this has become common knowledge. Indeed, the bone marrow is a classic source of these cells. There is evidence that MSCs can mobilize from the bone marrow and enter the circulation [12,13]. This intravazation involves migration of MSCs in a basal-to-apical direction across the bone marrow endothelium and the associated mechanisms remain to be elucidated. This is in contrast to hematopoietic stem cells whose mechanism of mobilization and migration has been comparatively well documented, involving chemokines and adhesion molecules [14,15]. It is also possible for MSCs to extravasate from the circulation into the bone marrow, involving apical-to-basal migration across the bone marrow endothelium, but as with mobilization little is known about the cues required for this migration to happen
MSCs have been reported to possess some of the relevant molecules to enable their migration, including adhesion molecules and chemokine receptors. VCAM-1 on endothelial cells and its interaction with VLA-4 with MSCs have been shown by several groups to be involved in MSC adhesion to endothelial cells [33 –35]. However, Brooke et al. [36] report that human MSCs (hMSCs) from bone marrow have low levels of VLA-4 and do not bind to VCAM-1, so the exact manner of MSC firm adhesion to the endothelium remains to be clarified. Although there have been many studies on the role of chemokines in MSC migration in vitro [37 –45], using standard chemotaxis assays, there is a lack of information concerning their ability to migrate across endothelial cells, either in an apical-to-basal direction or vice versa, and the mechanisms involved. This is despite in vivo evidence that IV-injected MSCs can extravasate across endothelial cells into tissues [29 –32]. Much more is known concerning the mechanism by which leukocytes adhere to endothelial cells and transmigrate across this layer [46,47].
The aim of the present study was to examine the migratory cues that stimulate MSC migration in a basal-to-apical direction and apical-to-basal direction across bone marrow endothelial cells (BMECs) in an in vitro model. The chemokines CXCL12, CXCL13, CXCL16, CCL11, and CCL22 were selected since these are ligands for the chemokine receptors CXCR4, CXCR5, CXCR6, CCR3, and CCR4, respectively, which have been identified on the current population of hMSCs [39].
Materials and Methods
Cell culture
Immortalized HBMECs were kindly donated by Dr. B.B. Weksler [48]. These cells were cultured in DMEM–F12 medium (Lonza) containing 10% FBS, 50 U per mL penicillin and streptomycin, and 2 μg/mL fungizone (from now on referred to as complete medium), in a humidified environment at 37°C, 5% carbon dioxide in air. All other cells were cultured in this medium and environment unless stated otherwise.
Ethics approval was obtained to obtain hMSCs from cancellous bone samples taken from consented patients undergoing total hip replacements for osteoarthritis (n=9, 5 male and 4 female, aged 61–85). Patients were healthy apart from having osteoarthritis and had no other medical conditions. The cancellous bone was flushed through with medium until the bone samples were a pale creamy color [39]. The bone was then discarded. The medium containing the cells was then put through a 70 μm filter into a centrifuge tube and spun down at 2,000 rpm for 8 min. The supernatant was then poured off and the cells resuspended in 10 mL of medium and layered over 10 mL of lymphoprep and spun at 2,400 rpm for 20 min. The cell layer was then removed and placed in a 175 cm2 flask containing complete medium. The medium was changed 24 h later, the nonadherent cells were removed, and the remaining cells were grown until confluent, and passaged as required. Cells were used between passages 2 and 5 and their phenotype was tested; the cells were plastic adherent, CD105 positive (<95%) and negative (0%) for CD45 and CD34 by flow cytometry, and when incubated in osteogenic and adipogenic medium stained for alkaline phosphatase and lipid, indicating that they could differentiate down osteogenic and adipogenic lineages (data not shown). The MSCs were used between passages 2 and 5 because MSC chemokine receptors and their migratory responses to chemokines are down-regulated with prolonged culture, at 12 and 16 passages [38]. All hMSCs were stored and used according to human tissue act guidelines.
Live cell video recording of MSC migration
About 1×105 HBMECs were seeded in a 24-well tissue culture-treated plate and grown until confluent in the medium. The medium was then removed and replaced with fresh serum-free medium containing the required chemokine (Peprotech) for 30 min; 100 ng/mL of chemokine was used since this was found to be the optimum concentration to stimulate MSC migration from preliminary dose responses (0–500 ng/mL). After this 30 min incubation the medium was removed and replaced with serum-free medium containing 10,000 hMSCs. The cells were then observed using a phase-contrast microscope in a heated chamber, at ×10 objective magnification and maintained at 37°C. Cell-F imaging software was used to take a photograph of the stem cells on top of the endothelial layer every 3 min for 15 h. These images were then linked together at a rate of 10 frames per second to make a video. Each stem cell was closely watched and registered as either migrated or not migrated. A cell was classed as migrated if it settled down onto the endothelial layer at this stage looking phase bright, the cell would then flatten, put out microvilli, become phase dark, and disappear. On average, 50 MSCs per experiment were counted.
MSC transendothelial migration using transwells
Eight micrometer polyethylene terephthalate (PET) membranes in hanging well inserts (Millipore) were coated on both the basal and apical surface with 4 μM fibronectin in PBS for 1 h, the fibronectin was then removed, and the filter was left to dry for a further hour. To examine apical-to-basal migration the filters were then placed in a 24-well tissue culture-treated plate containing 800 μL of serum-free medium per well. About 1×105 HBMECs in 400 μL of serum-free medium were then added to the apical surface and incubated for 48 h. To examine basal-to-apical migration the filters were turned upside down in a Petri dish, which was then filled with serum-free medium until it was just touching the apical surface, but not covering the basal surface of the filter. About 1×105 HBMECs in 80 μL of serum-free medium were then added to the basal side of the filter, and left to adhere for 1 h. The filter was then turned upright and hung in a 24-well plate and incubated for 48 h with 800 μL serum-free medium in the basal well and 400 μL in the apical well. The apical surface was then scraped and washed.
The required number of flasks containing hMSCs were then trypsinized, spun down, and resuspended in serum-free medium containing 8 μM calcein AM. The hMSCs were then incubated for 1 h to allow uptake of the calcein. During this incubation the medium was removed from the basal and apical surfaces of the transwell filters containing HBMECs and replaced with serum-free medium containing the required chemokine (at 100 ng/mL) for 30 min. The medium was then removed from the apical surface of the transwell and replaced with 400 μL of serum-free medium containing 50,000 calcein labeled hMSCs. These were then left to migrate overnight for approximately 15 h; this time point was chosen since live cell video-recording at the apical endothelial surface showed that it was more than sufficient time for transendothelial migration to occur (around 8–10 h, Fig. 1B). The filters were fixed in 70% methanol for 20 min, the apical surfaces were then scraped to remove any endothelial layer and excess hMSCs that had not migrated. The filters were then gently rinsed in PBS. The total number of fluorescent hMSCs over the entire basal surfaces of the filters were then counted down a microscope at ×100 magnification and recorded as the number of migrated cells.
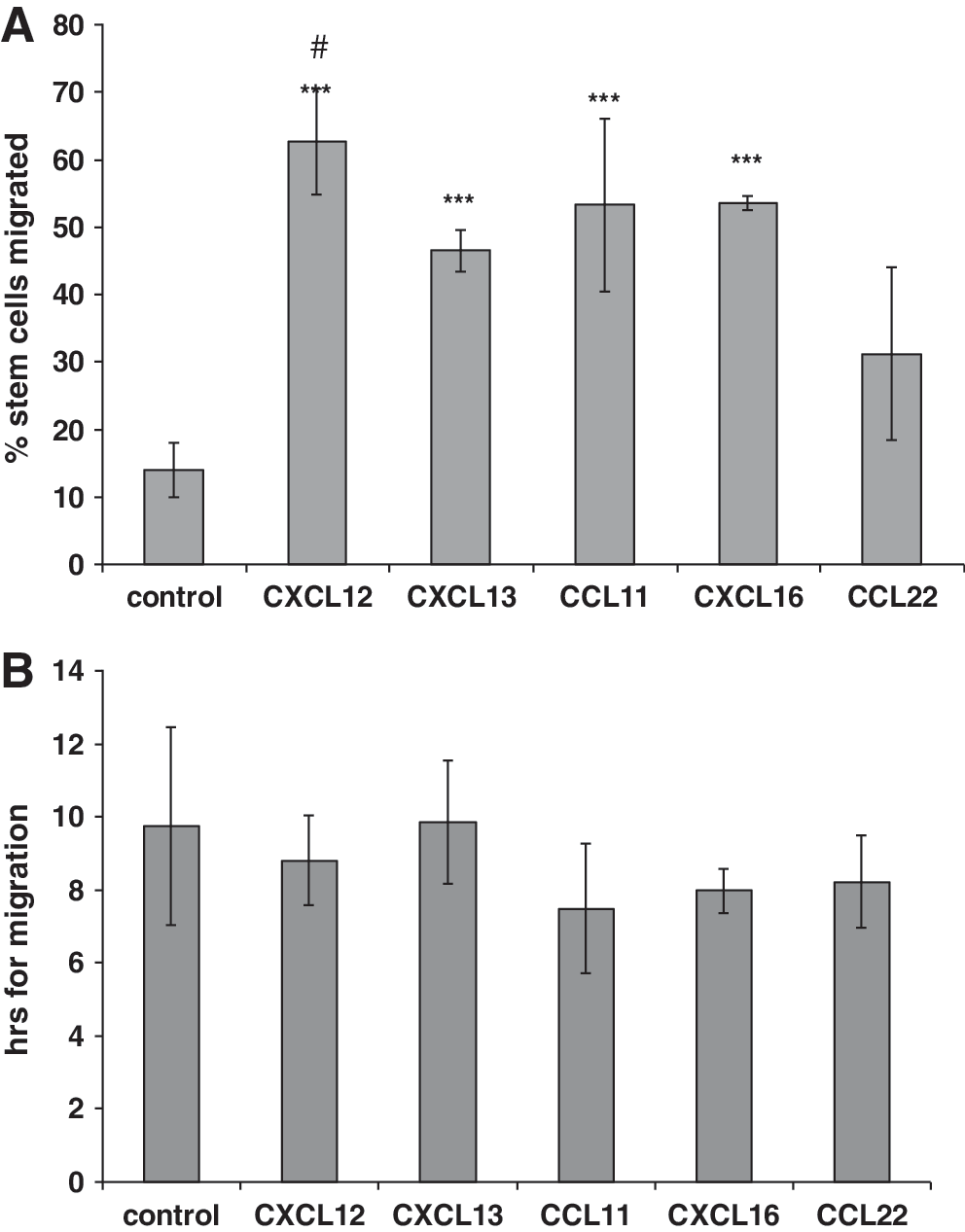
Chemokine-stimulated migration of human mesenchymal stem cells (hMSCs) across human bone marrow endothelial cells (HBMECs) using live cell video recording. HBMECs were cultured as plastic-adherent monolayers and treated with chemokines CXCL12, CXCL13, CXCL16, CCL11, and CCL22 at 100 ng/mL for 30 min or left untreated (control). hMSCs were then added and photographed every 3 min for 15 h under phase contrast, and videos made. Cells that migrated across the HBMECs were counted and expressed as a percentage of total cells and the time taken for migration to occur was noted. On average, 50 MSCs were counted per experiment. CXCL12, CXCL13, CXCL16, and CCL11 significantly increased (***P<0.001) the percentage of MSCs that migrated compared to control
Transmission electron microscopy
MSCs (not labeled with calcein) were incubated on fibronectin-coated 8 μm pore transwell filters precultured with a layer of HBMECs as for apical-to-basal transendothelial migration assays (see above). The filters were then fixed for 2 h at room temperature in 0.1 M sodium cacodylate and 2 mM CaCl2 (buffer A) containing 2.5% glutaraldehyde, before being stored in buffer A, containing 0.1% glutaraldehyde. The samples were then washed 3 times for 5 min in buffer A, before postfixation using 0.1% osmium tetroxide in buffer A for 1 h. The samples were then washed again and stored overnight in 70% ethanol. The samples were dehydrated in increasing concentrations of ethanol (80% and 100%) for 15 min followed by 15 min in 100% dry ethanol. Spurr's resin and dry ethanol was used to infiltrate the samples with ratios of 1:3 for 1 h, 1:1 for 1 h, and 3:1 overnight, followed by pure Spurr's resin for three 1-h incubations and polymerization at 60°C for 16 h. Sections 100 nm thick were cut and collected on to copper grids, which were then stained with 2% lead citrate and 2% uranyl acetate before observation with a JEOL electron microscope.
Statistics
To compare the effects of chemokines on migration of MSCs across HBMECs ANOVA was performed followed by Dunnett's and Tukey's post-tests to compare chemokine treatments and with control (P<0.05 deemed as significant).
Results
hMSC migration in an apical-to-basal direction across BMECs
Since the presence of chemokine receptors has been variously reported on MSCs [37 –45], initial experiments were performed to characterize the spectrum of chemokine receptors expressed by the hMSCs used in the present study. The MSCs expressed a broad range of chemokine receptors. Using flow cytometry the percentage positive cells were as follows (mean±SD): CXCR1 55%±37%; CXCR2 22%±13%; CXCR3 43%±19%; CXCR4 96%±4%; CXCR5 94%±5%; CXCR6 95%±3%; CCR1 43%±17%; CCR2 40%±16%; CCR3 98%±2%; CCR4 13%±12%; CCR5 78%±12%; CCR6 17%±9%; CCR7 29%±7%; CCR8 10%±12%; CCR9 64%±1% CCR10 48%±37%. These results and associated methods have already been fully reported in a previous study using the same preparations of hMSCs [39]. In the present study it was decided to focus on chemokine receptors CXCR4, CXCR5, CXCR6, and CCR3 since these were the most highly expressed and to include a chemokine receptor that was expressed to a lesser extent, namely, CCR4. The chemokine ligands chosen for these receptors were CXCL12, CXCL13, CXCL16, CCL11, and CCL22, respectively.
HBMECs are a human bone marrow endothelial cell line that expresses endothelial cell markers and adhesion molecules [48]. They express E-selectin, V-CAM, and I-CAM and support rolling and firm adhesion of CD34+ hematopoietic progenitor cells, and have provided a model to study hematopoietic stem cell homing. In the current study HBMECs were treated with 100 ng/mL CXCL12, CXCL13, CXCL16, CCL11, or CCL22 and MSCs added to their apical surfaces. This model is similar to that used by others who used apically applied chemokines to endothelial cells to study leukocyte transendothelial migration [49,50]. Using live cell video imaging the numbers of MSCs that migrated across the HBMECs in the presence or absence of chemokines were counted (Supplementary Video S1; Supplementary Data are available online at
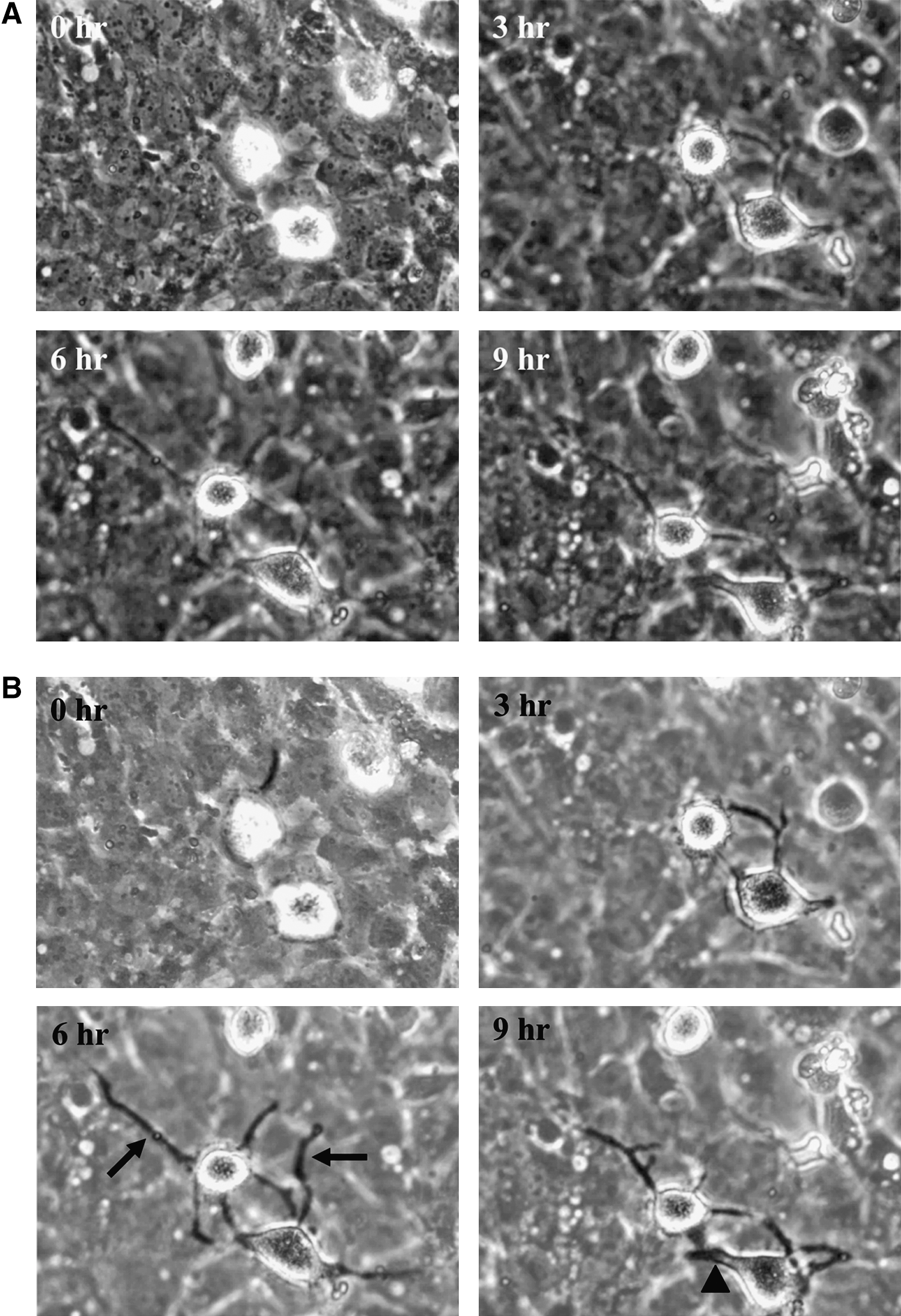
Live cell images from a video recording showing hMSC adhesion to HBMECs treated with CXCL12. HBMECs were cultured as plastic-adherent monolayers and treated with CXCL12 at 100 ng/mL for 30 min (as Fig. 1). hMSCs were then added and photographed every 3 min for 15 h under phase contrast, and videos made.
After the increase in MSC migration with chemokines using HBMECs in flat culture dishes, the same chemokine treatments were repeated in an 8 μm pore transwell system to confirm that the stem cells were fully transmigrating and not integrating into the endothelial layer as previously described [34]. HBMECs were grown on transwell filters, treated with chemokines, and MSCs added to the apical endothelial surfaces. In the presence of CXCL12, CXCL13, CXCL16, CCL11, or CCL22 (100 ng/mL), there was a significant increase in the number of stem cells that had migrated across the HBMECs and adhered to the underside of the filter compared with control (P<0.05, P<0.01, P<0.001, P<0.01, and P<0.001, respectively) (Fig. 3A). This amounted to increases of 2-, 3.6-, 5.6-, 4.8-, and 3.1-fold for CXCL12, CXCL13, CXCL16, CCL11, and CCL22, respectively, compared to the no chemokine control. There were several differences between the chemokines; CXCL16 increased the migration of MSCs significantly more than CXCL12 (P<0.001), CCL22 (P<0.01), and CXCL13 (P<0.05).

Chemokines enhance bidirectional migration of hMSCs across HBMECs in transwells. For luminal-abluminal migration, HBMECs were grown on the apical surfaces of 8 μm pore transwell filters. For abluminal-luminal migration, filters were inverted and seeded with HBMECs on the basal filter surfaces; the filters were then turned upright and placed in the transwell. For both migration directions, calcein-labeled hMSCs were placed in the upper wells and 100 ng/mL CXCL12, CXCL13, CXCL16, CCL11, or CCL22 added, with control being in the absence of chemokine. After 15 h the total number of fluorescent MSCs over the entire basal surface was counted.
These transwell experiments involved adding chemokines to the basal and apical chambers. This was to reflect live cell imaging experiments in which apical chemokine was applied, as well as to providing a basal source of chemokine. In addition, classical transmigration experiments were performed with chemokines CXCL12, CXCL13, CXCL16, and CCL11 added only to the basal chamber (ie, basal endothelial surface) and MSCs to the apical chamber to compare with the above experiments. Chemokines CXCL12, CXCL13, and CXCL16 gave significant increases in luminal to abluminal migration at 100 ng/mL of chemokine, whereas for CCL11 500 ng/mL was required for a response (Fig. 3C). In addition, the fold-changes at 100 ng/mL were 1.2, 1.4, and 1.1 for CXCL12, CXCL13, and CXCL16, respectively, when the chemokine was only applied basally (Fig. 3C), which were much lower compared to when the chemokine was applied basally and apically (Fig. 3A).
Filters from the same apical-to-basal migration method, after having been seeded with HBMECs, treated with chemokines and MSCs, and incubated for 6 h or overnight, were imaged by transmission electron microscopy (TEM). This showed that in the presence of chemokine the stem cells at 6 h were putting out many long and fine microvilli all over the MSC when at a distance from the endothelial layer, and mainly at the cell periphery and ventral surface of the MSC when in close contact with the endothelial layer (Fig. 4A). The MSCs also put out thicker podosomes, which appeared to probe the endothelial surface (Fig. 4B). These thicker protrusions were only seen when the MSC was in close proximity of the endothelial layer. After overnight migration MSCs were seen to be flattened on the endothelial cells with a high degree of cell–cell contact (Fig. 4C), and podosome-like protrusions were observed penetrating the endothelial cell layer. At this stage the MSCs that were actively migrating did not have large amounts of fine microvilli. In the absence of chemokine the MSCs rested on the endothelial cell layer but did not extend microvilli or podosome-like structures (Fig. 4B inset).
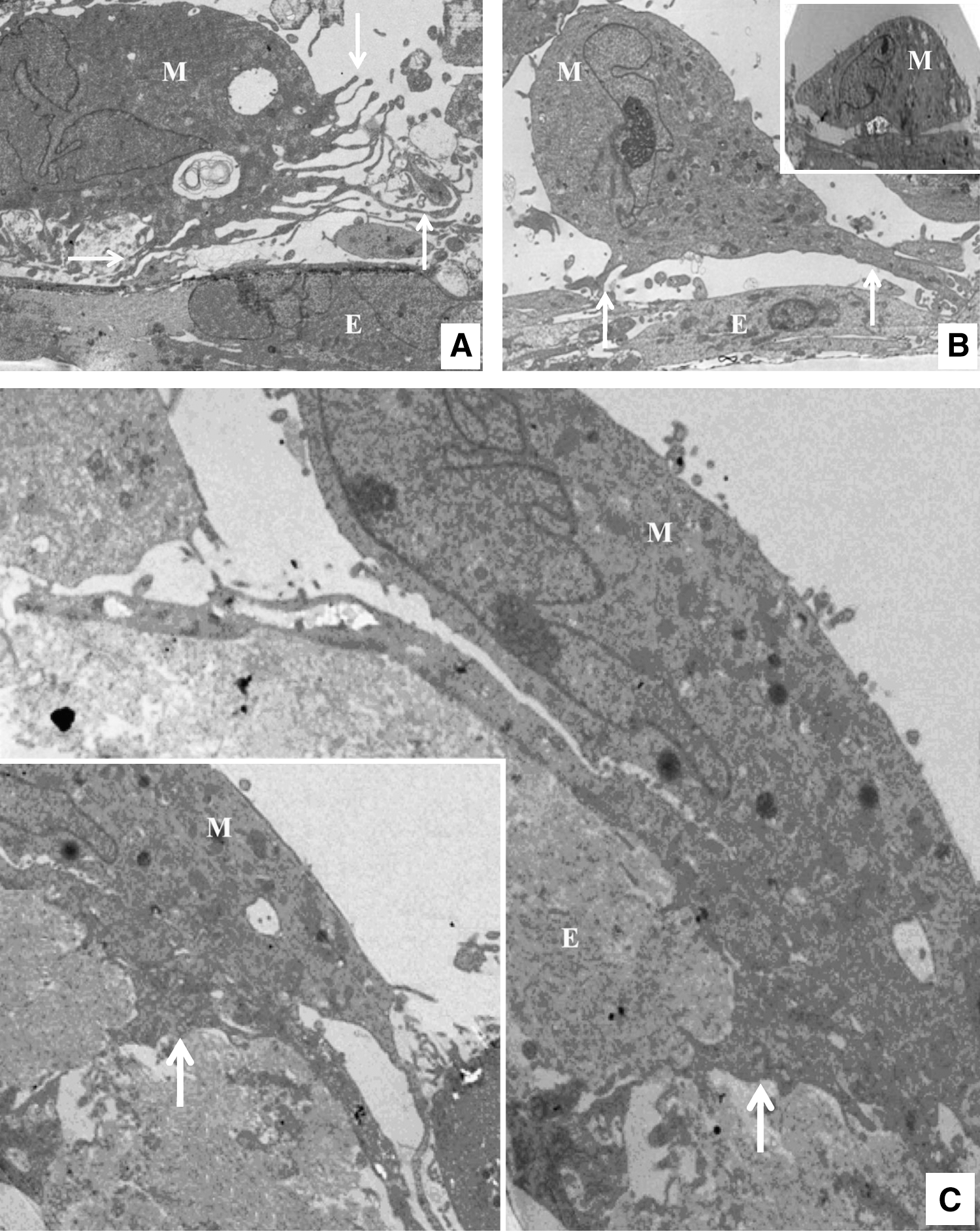
Electron microscopy images of hMSCs and their interaction with HBMECs in the presence of CXCL16 after 6 h
hMSC migration in a basal-to-apical direction across BMECs
Next the migration of MSCs in the opposite basal-to-apical direction was examined by developing a reverse transmigration model. To do this the transwell filters were inverted and the HBMECs were seeded on the underside of filter, allowed to adhere for 1 h and then turned the right way up and grown as normal, hanging in a 24-well plate. Before treatment with any chemokines the apical surface of the filter was carefully scraped and washed with PBS to remove any cell layer that may have gone through the filter. Analysis of the filters showed that treatment with CXCL12, CXCL13, CXCL16, or CCL11 all significantly increased transendothelial stem cell migration (P<0.01), with 2-, 1.7-, 2.4-, and 2-fold increases, respectively, compared to control (Fig. 3B). However, the most significant increase was with CCL22, which gave a large significant difference (P<0.001) and a 5.3-fold increase compared to control. There were several differences between the chemokines; CCL22 significantly increased the migration of MSCs compared to CXCL12, CXCL13, CXCL16, and CCL11 (P<0.001 in each case).
Discussion
The results of the present study show that chemokines can enhance the migration of MSCs bidirectionally, in an apical-to-basal direction and basal-to-apical direction across BMECS. Chemokines CXCL12, CXCL13, CCL11, and CXCL16 had significant effects stimulating transmigration in both directions.
There is evidence that MSCs can migrate from the circulation and into the bone marrow [16 –21]. MSCs have been used in bone marrow transplantation and graft-verses-host diseases; in these situations the MSCs have shown some degree of engraftment into the bone marrow and clinical benefit [24,51,52]. Clinical trials that utilize MSCs to treat graft-versus-host disease have been started and suggest that MSCs will improve cell and solid organ transplantation by reducing rejection and may eliminate the present need for lengthy immunosuppressive drugs regimens [51]. In addition, following total body and local radiation there is elevated engraftment of MSCs to the bone marrow [53]. Systemic administration of MSCs into patients with osteogenesis imperfecta leads to increased bone formation and reduced frequency of bone fracture, indicating that MSCs can migrate into the patient's bone giving rise to osteoblasts [22,23]. In mice intravenous administration of MSCs results in recruitment of these cells to the bone, resulting in their differentiation to osteoblasts and osteocytes [18,19]. There is also evidence that MSCs can mobilize in response to injury and inflammation and enter the circulation [12] and recent evidence suggests this is regulated by VEGF and CXCR4 [54]. For example, MSCs can be mobilized from the bone marrow into the blood leading to their recruitment to the heart following myocardial infarction, and resulting in MSC differentiation into cardiomyocytes [13]. Mansilla et al. [55] found significantly higher numbers of MSCs in peripheral blood taken from acute burns patients when compared to healthy controls, amounting to a 20-fold increase. Further, in response to muscle injury there is increased mobilization of MSCs into the blood [56]. Thus, the increased numbers in peripheral blood may be required to home to sites of injury, trauma, and inflammation to aid regeneration of the damaged tissues. The results of the present study suggest that chemokines and their receptors may stimulate luminal-to-abluminal transendothelial migration (and vice versa) during recruitment from the blood to the bone marrow and mobilization of MSCs from the bone marrow to the circulation. These mechanisms have been widely examined for other cell types such as hematopoietic stem cells and leukocytes. For example, CXCL12/CXCR4 mediates the mobilization hematopoietic stem cells from the bone marrow and into the circulation, and homing to this tissue [57]. In addition, ELR+ CXC chemokines and CXCR2 mobilize neutrophils from the bone marrow into the circulation, and CXCL12/CXCR4 plays a role in the trafficking of senescent neutrophils back to the bone marrow [58].
Some differences between the chemokines and their effects on the direction of MSC transendothelial migration were apparent. In terms of their ability to stimulate apical-to-basal migration, CXCL16 was the most effective (6-fold stimulation), with the rank order being CXCL16>CCL11>CXCL13>CCL22>CXCL12. Whereas in the basal-to-apical direction CCL22 was the most effective (5-fold enhancement) with the remaining chemokines being roughly equal. This indicates that chemokines differ in their abilities to stimulate the direction of MSC migration across BMECs.
The chemokines CXCL12, CXCL13, CXCL16, and CCL11 all significantly enhanced MSC migration across BMECs in an apical-to-basal direction using live cell imaging at the apical endothelial surface and by counting the numbers that had transmigrated across the endothelial cells to the basal surface of the transwell insert. Both methods gave broadly similar results for CXCL13, CXCL16, and CCL11. However, when imaging at the apical endothelial surface CXCL12 gave the greatest effect compared to other chemokines, whereas counting the transmigrated MSCs CXCL12 gave the lowest effect. This suggests that in the presence of CXCL12 some MSCs may not fully transmigrate but remain associated with the endothelial cell layer. It has been shown that MSCs can integrate into the endothelial cell layer or associate with the abluminal endothelial surface [34,59] and CXCL12 may enhance this effect. The other chemokines CXCL13, CXCL16, and CCL11 appear to differ from CXCL12 in this respect.
There have been several reports documenting the presence of chemokine receptors on hMSCs and their respective chemokine ligands stimulated migration across filters in standard chemotactic chambers in the absence of endothelial cells. For example, expression of CXCR4 on MSCs has been demonstrated by several groups and CXCL12 stimulated MSC chemotaxis [37,38,41 –44]; the percentage of cells reported to express CXCR4 by these groups ranges from 1% to 43% of cells. In contrast, Von Lüttichau et al. [40] found no expression of CXCR4 on the cell surface of MSCs. In our study, there was high expression of CXCR4 on MSCs and CXCL12 enhanced transendothelial migration. Expression of CXCR5 has been shown by Von Lüttichau et al. [40], on a small proportion of cells, and by Honczarenko et al. [38], on 80% of MSCs and CXCL13 enhanced MSC migration in both studies. Functional CXCR6 expression has been shown by 3 groups [37,38,44] on 22%, 43%, and an unspecified percentage, respectively. In the present study, CXCR5 and CXCR6 were highly expressed on MSCs, on 94% and 95% of MSCs, respectively, and their ligands CXCL13 and CXCL16 enhanced transendothelial migration. Two groups have reported CCR3 expression by MSCs [43,44] and when these cells were stimulated with inflammatory cytokines, such as TNF-α, the expression increased. In the present study, CCR3 was detected on basically all MSCs (98%) and its ligand CCL11 was effective in driving transmigration across HBMECs. CCR4 has been reported on MSCs at low/intermediate level and CCL22 enhanced MSC migration [40,43]. Our data are in agreement showing low expression of CCR4 and there was a transendothelial migratory response to CCL22. In fact, CCL22 was the most effective chemokine at enhancing basal-to-apical migration, agreeing with other studies showing that even though expression of chemokine receptor may be low there is still a migratory response. Other reports, however, have found CCR4 to be absent on MSCs [38,45]. Differences in the level of expression of chemokine receptors between different studies may be due to differential receptor internalization and degradation or storage, the source of MSCs, different culture conditions, and the methods used for their detection; it could also be a feature of ex vivo highly expanded cells [38,60].
The MSCs showed various morphological changes when they interacted with the HBMECs before their transendothelial migration. They flattened, put out microvilli (or filopodia) and podosome-like protrusions, which inserted into the endothelial cell layer. These microvillous structures resemble the filopodia that have been seen on leukocytes that interact with the endothelial cell surface; such filopodia are proposed to promote lymphocyte crawling and probing before transendothelial migration [61]. They also resemble the long plasmic podia that have been described on hMSCs interacting with unstimulated endothelial cells [34]. Other structures have been described on leukocytes that interact with endothelial cells. These include invadosome-like protrusions or podosomes that probe the endothelial surface, and are suggested to play a role in identifying and exploring locations permissive for diapedesis [62]. In the present study TEM showed similar podosome-like structures on the ventral surface of MSCs that had inserted into the endothelial cell layer. Therefore, MSCs appear to possess several similar microstructures identified for leukocytes that have been shown to play a role in transendothelial migration.
In the present study, the HBMECs were treated with chemokines for 30 min, and MSCs added, which were then left to migrate for 15 h. Live cell imaging indicated that transendothelial migration took on average around 8–10 h to occur. During this time, chemokines would be bound by the HBMECs and presented to chemokine receptors on MSCs resulting in enhanced migration. However, it cannot be ruled out that some of the stimulatory effects on MSC migration may be due to chemokines acting directly on the HBMECs, which may provide pro-migratory signals.
The chemokines studied in the current study are present in the bone marrow. There has been particular study of CXCL12 and this chemokine is produced in the bone marrow and occurs in its endothelial cells in situ [63 –65]. CXCL13 can be produced by HBMECs [65], with expression also occurring in osteoblasts and bone marrow stromal cells [67]. CXCL16 is produced in the bone marrow, including stromal cells [64]. There is evidence that CCL11 can be expressed by bone marrow cells [68] and CCL22 can be produced by mast cells from bone marrow [69]. Therefore, the chemokines shown to enhance MSC transendothelial migration can be present in the bone marrow environment and occur on the endothelial cells.
In conclusion, chemokines can enhance the migration of MSCs bidirectionally across HBMECs, with directional preferences shown for different chemokines. These molecules and their receptors may be exploited therapeutically in the future to enhance the recruitment of MSCs to the bone to stimulate hematopoiesis and in pathologies such as graft-versus-host disease, and enable more MSCs to enter the circulation from the bone marrow to help the regeneration remote tissues in the body.
Footnotes
Acknowledgments
We wish to thank the orthopedic surgeons at the RJAH Orthopaedic Hospital for providing the bone marrow tissue samples, and Karen Walker, EM Unit Keele University, for performing the TEM. This work was supported by a grant from the Biotechnology and Biological Sciences Research Council (UK).
Author Disclosure Statement
There are no commercial associations of the authors that create conflicts of interest in connection with the article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.