
Abstract
There has been a recent increase in our understanding in the isolation, culture, and differentiation of mesenchymal stem cells (MSCs). Concomitantly, the availability of MSCs has increased, with cells now commercially available, including human MSCs from adipose tissue and bone marrow. Despite an increased understanding of MSC biology and an increase in their availability, standardization of techniques for adipogenic differentiation of MSCs is lacking. The following review will explore the variability in adipogenic differentiation in vitro, specifically in 3T3-L1 and primary MSCs derived from both adipose tissue and bone marrow. A review of alternative methods of adipogenic induction is also presented, including the use of specific peroxisome proliferator-activated receptor-gamma agonists as well as bone morphogenetic proteins. Finally, we define a standard, commonly used adipogenic differentiation medium in the hopes that this will be adopted for the future standardization of laboratory techniques—however, we also highlight the essentially arbitrary nature of this decision. With the current, rapid pace of electronic publications, it becomes imperative for standardization of such basic techniques so that interlaboratory results may be easily compared and interpreted.
Introduction
T
This potential variation in technique is only compounded by the commercial availability of MSCs derived from a variety of species and tissue types. Although the fact that one can order overnight as many viable human stem cells as desired is an amazing accomplishment of science, communication, and transportation, it brings with it several problems. For example, companies often send with their MSCs a proprietary medium whose contents are highly controlled but not reported to the customer. This practice is clearly driven by economic realities but not scientifically justified.
It is on the backdrop of these obvious shortcomings that this concise review article will explore the extreme variability in adipogenic differentiation medium between institutions, specifically looking at 3T3-L1 cells and primary MSCs of adipose tissue and bone marrow origin. Presumptively, the adipogenic supplements for a cell line (3T3-L1) would be highly controlled in comparison to primary cells; however, we found significant variability among both cell types. A review of alternative methods of adipogenic induction is also presented, including the use of specific peroxisome proliferator-activated receptor-gamma (PPARγ) agonists as well as bone morphogenetic proteins (BMPs). Finally, we define a standard, commonly used adipogenic differentiation medium for each cell type with the hopes that this may prompt a standardization of basic laboratory practices. Although the standardization of basic in vitro assays is necessary, there exists by no means a “quick-fix” for this dilemma. However, our hopes are that this and similar reviews will bring attention to this pressing scientific problem.
Materials and Methods
An exhaustive literature review was performed using PubMed. Search terms included “adipogenic differentiation 3T3-L1,” “bone marrow mesenchymal stem cell adipogenic differentiation,” “adipose stem cell adipogenic differentiation,” “rosiglitazone adipogenic differentiation,” and “bone morphogenetic protein adipogenic differentiation.” Results were stratified by species of origin, focusing only on those articles describing culture of either mouse or human cells. All articles in 2010 with full text available were examined. Finally, those companies that sell propriety adipogenic differentiation medium were contacted in the hopes that they would share their standardized recipes. No commercial entities were willing to share their medium components.
Results
Adipogenic differentiation of 3T3-L1 cells
3T3-L1 cells are the most commonly studied adipogenic cell line that is available through American Type Culture Collection (American Type Culture Collection No. CL-173). The L1 substrain of 3T3 was developed through clonal isolation. Generally, 3T3-L1 cells undergo adipogenic differentiation rapidly, within 1 week in most instances. In the last year, 45 articles have been published across United States, European, and Asian academic centers (Fig. 1A) [1 –45]. As demonstrated in Fig. 1B, articles using 3T3-L1 cells have been published in a wide range of journals including Biochemistry, Cell Biology, Endocrinology, and others (Fig. 1B). The majority of articles use standard Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum; however, this varies. Additional components are numerous; however, generally 3 components are used for nearly all differentiation of 3T3-L1 cells. These standardly include insulin, dexamethasone, and IBMX. For a cell line, all components were found to vary considerably in concentration from article to article (Fig. 1C). Insulin is widely used to induce proliferation and differentiation of preadipocytes [46]. At high concentrations, insulin is known to mimic insulin-like growth factor-1, activating mitogen-activated protein kinase pathways [47,48]. Dexamethasone is an anti-inflammatory steroid molecule that stimulates both osteogenic and adipogenic differentiation in a cell-, time-, and concentration-dependent manner [49,50]. However, when MSCs experience either prolonged exposure or increased concentrations of dexamethasone, they yield higher number of adipocytes in cultures while inhibiting osteogenic differentiation [51]. IBMX in combination with dexamethasone regulates PPARγ, promoting adipogenesis [52]. IBMX is a competitive, nonselective phosphodiesterase inhibitor, raising intracellular cAMP and protein kinase A (PKA). PKA signaling pathway is required for transcriptional activation of PPARγ and thus adipogenic gene expression [18]. In addition, both dexamethasone and IBMX are inducers of C/EBPδ and C/EBPβ, which are transcription factors for growth and differentiation [53]. Despite the consistent use of these 3 components, concentrations for each vary widely (Fig. 1C). For example, the concentration of insulin varies from 0 to 1,800 nM depending on the article. A list of additional components occasionally added, such as PPARγ agonists, is given in Table 1. A comprehensive breakdown of major components by article is available in Table 2.
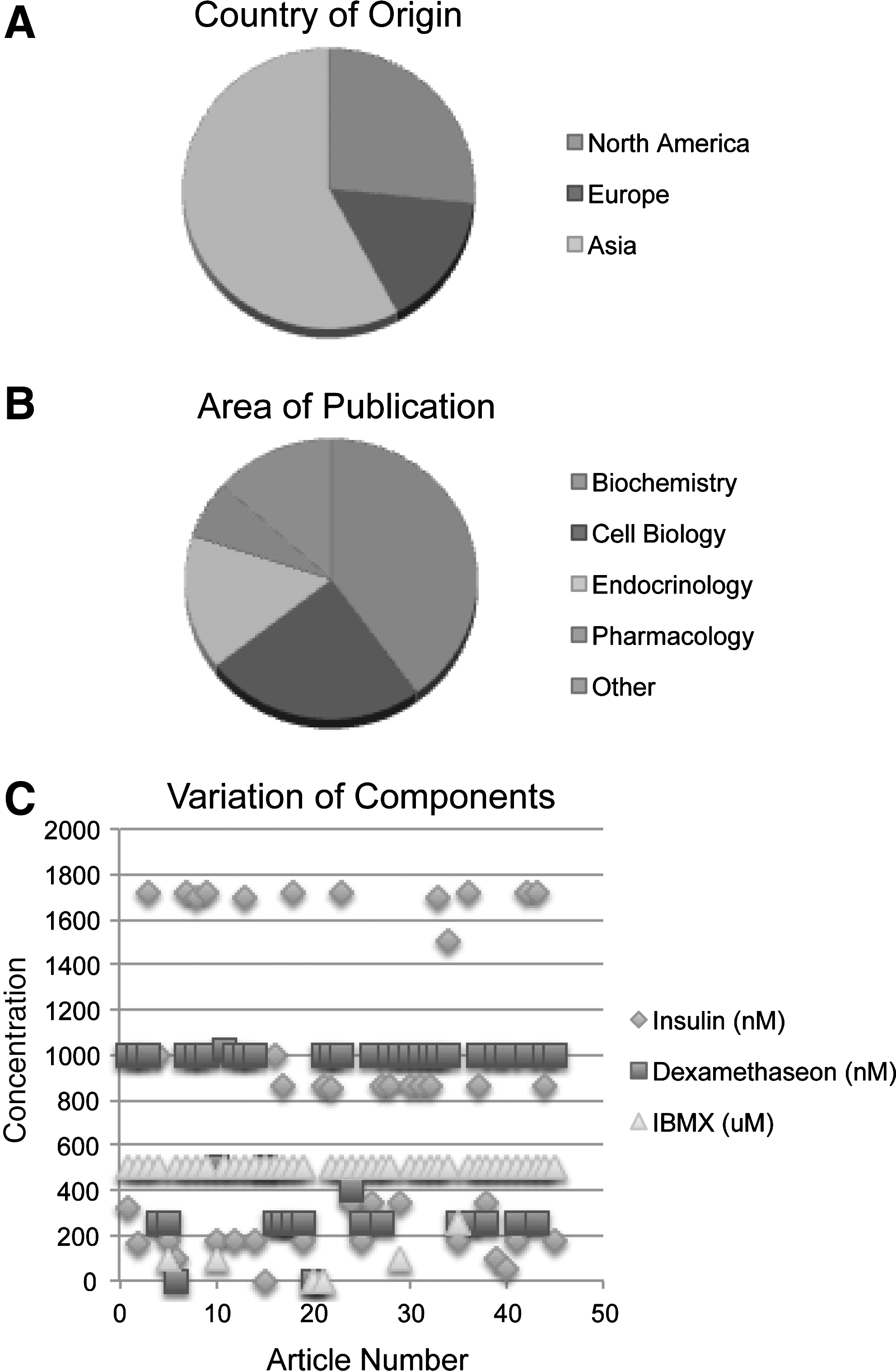
Variation in adipogenic differentiation of 3T3-L1 cells. A literature review for articles within 2010 was performed for any publication examining the adipogenic differentiation of 3T3-L1 preadipocytes.
FBS, fetal bovine serum.
Adipogenic differentiation of primary BMSCs
Unlike an immortalized cell line, primary cells are a heterogenous mixture of MSCs, unipotential and bipotential cells, and fibroblastic cells among numerous other cell types. With this variability in cell population, one would expect that the adipogenic differentiation of primary MSCs is, expectedly, even more variable (Fig. 2). For the purposes of this literature review, 2 of the most commonly studied MSC populations were examined: bone marrow mesenchymal stem cells (or BMSCs) and adipose-derived mesenchymal stem cells (most commonly abbreviated ASCs).
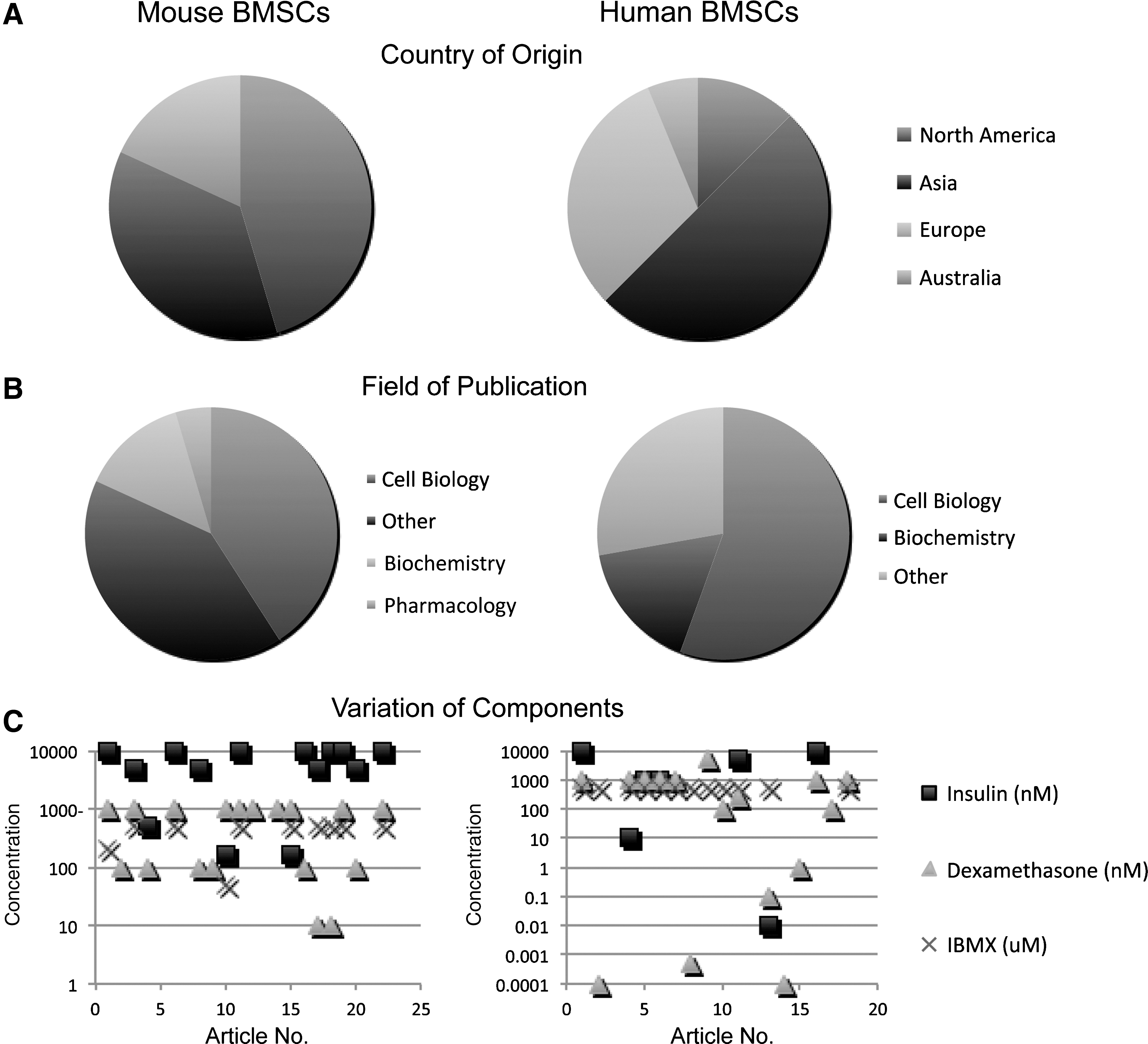
Variation in adipogenic differentiation of BMSCs. Again, a literature review for articles within 2010 was performed for any publication examining the adipogenic differentiation of BMSCs—broken down by either mouse (left) or human (right) origin.
In the last year, ∼20 articles have been published on the in vitro adipogenic differentiation of mouse and human BMSCs, respectively (see Tables 3 and 4 for a complete listing). The majority of these articles were published in the United States or Asia (Fig. 2A) and were primarily in the fields of cell biology and biochemistry (Fig. 2B). Generally, a 3-component cocktail is used for BMSC adipogenic induction, including insulin, dexamethasone, and IBMX (Fig. 2C). Vast inconsistency exists, however, between published protocols. Notice that a logarithmic scale is used for Fig. 2C, illustrating the extreme variability from 1 protocol to another. Moreover, a 1- or 2-component cocktail is sometimes used, with the other drugs simply omitted (Fig. 2C). A comprehensive breakdown of major components by article and by species is available in Tables 3 and 4.
Interestingly, the concentrations of adipogenic components used for mouse and human BMSC culture differ significantly. Figure 3 demonstrates the clear difference in the standard concentration of supplements when taking into account species of derivation. Insulin was not used in the majority of publications in human BMSCs, whereas a near 100% increase in dexamethasone and a presence of indomethacin was observed in human compared with mouse BMSCs (Fig. 3). Collectively, these results suggest overall that a different stimulus may be needed for human compared with murine BMSC adipogenic differentiation.
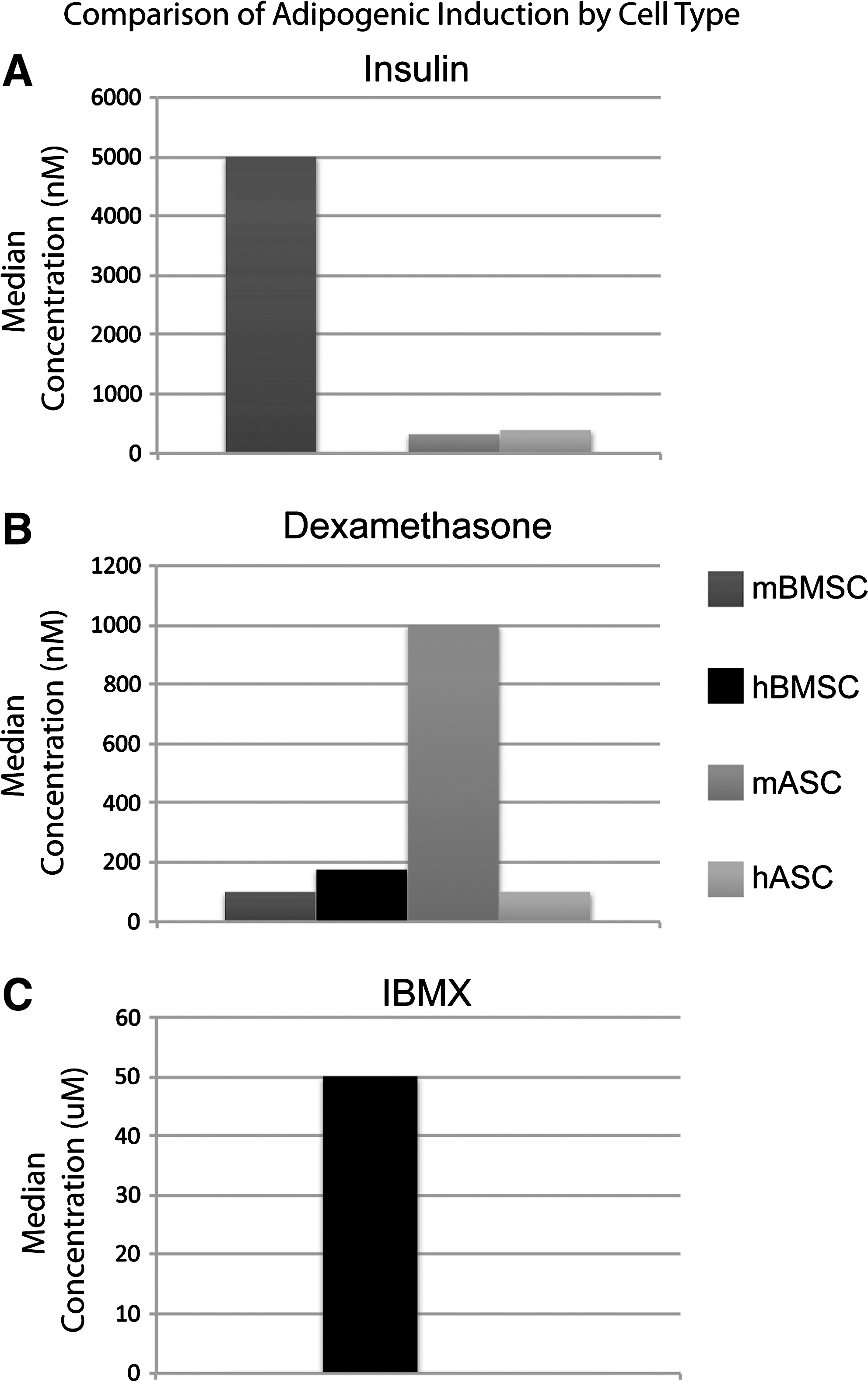
Differences in adipogenic differentiation of BMSCs and ASCs based on species. Median values for each component of BMSC/ASC adipogenic induction medium was calculated and compared between mouse and human cells.
Adipogenic differentiation of primary ASCs
In the last year, ∼20 articles have been published on the in vitro adipogenic differentiation of ASCs [22,54 –79]. The majority of these articles were published in the United States (Fig. 4A) and were primarily in the fields of biochemistry and cell biology (Fig. 4B). Generally, a 3–4-component cocktail is used for ASC adipogenic induction, including indomethacin, insulin, dexamethasone, and IBMX (Fig. 3C). As with BMSCs, vast inconsistency exists between published protocols (Fig. 4C). A few articles use propriety, undisclosed components for adipogenic induction [75]. A comprehensive breakdown of major components by article is available in Tables 5 and 6. Interestingly, and in similarity to BMSCs, the differences in induction components between ASCs of mouse or human origin differ significantly (Fig. 3). For example, insulin concentration is approximately equivalent between mouse ASCs (mASCs) and human ASCs, whereas an ∼10-fold increase in dexamethasone concentration was observed in mASCs in comparison to their human counterpart (Fig. 3). These results again suggest clear differences in adipogenic induction between mouse and human MSCs.
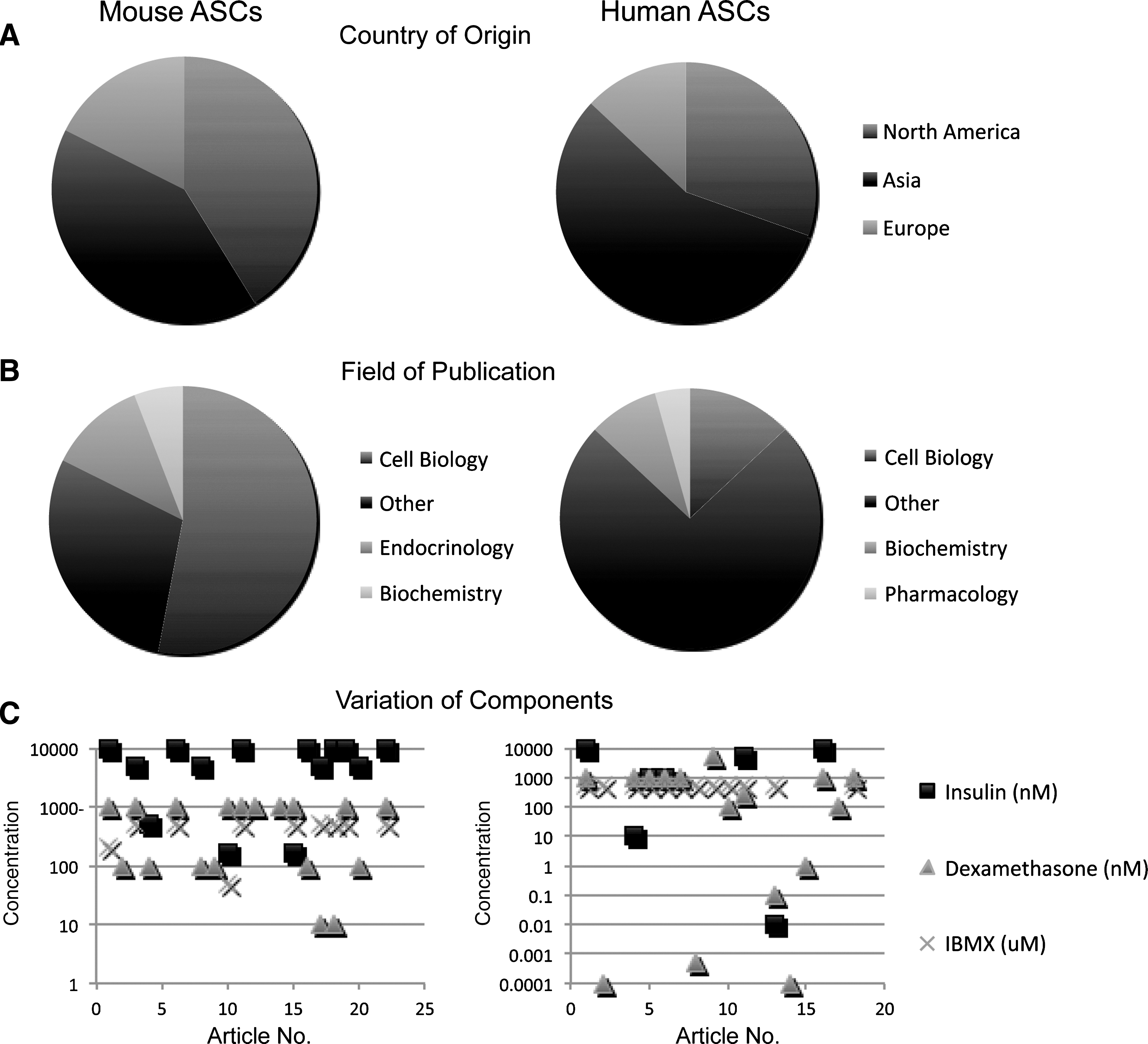
Variation in adipogenic differentiation of ASCs. Again, a literature review for articles within 2010 was performed for any publication examining the adipogenic differentiation of ASCs—broken down by either mouse (left) or human (right) origin.
Use of PPARγ agonists
Other specialized components have been used to induce adipogenesis, either in addition to the standardized cocktail of agents or alone. One of the most commonly studied is the PPARγ agonist rosiglitazone, as well as similar agents (troglitazone, etc.). PPARγ agonists (thiazolidineldiones or glitazones) are not only a boon to those under treatment for diabetes but also, in general, work to speed up the differentiation of preadipocytes or adipoprogenitor cells in vitro. Rosiglitazone binds to PPARγ, thus “sensitizing” fat cells to insulin. It is known that glitazones reduce bone mineral density—postulated to be via diverting MSCs to adipogenesis rather than osteogenesis in vivo [80,81]. Glitazones are also known to increase bone loss via stimulation of osteoclasts and promotion of bone resorption [80]. There exists some debate as to the extent that rosiglitazone is able to induce MSC adipogenesis as a single agent [82]; however, in general, rosiglitazone both speeds and increases the degree of differentiation of adipoprogenitor cells. Thus, addition of rosiglitazone may be a useful addition to the standard adipogenic induction cocktail if cells are of late passage or otherwise resistant to speedy differentiation. A standard dose of 1 μM rosiglitazone is suggested.
Use of BMPs
BMPs are a subset of the transforming growth factor-β superfamily, so named as they were first observed to induce osteogenic differentiation when implanted in muscle pouch model. BMPs are powerful osteoinductive agents, and they have clear pleiotropic effects, including the induction of chondrogenesis [83], adipogenesis [84 –86], and angiogenesis [87]. In some specific scenarios, the stimulation of BMP2 on adipogenesis results in the formation of cyst-like bone voids filled with lipid [88,89]. For example, in a recent study, BMP2 was implanted at high doses in a femoral defect in rats [90]. It was observed that there exists a dose-dependent increase in the formation of cyst-like bone voids with escalating doses of BMP2. Similar observations have been made in an ectopic bone formation model (nude mouse muscle pouch) by 2 independent investigators—in which various BMPs were observed to induce “lipid-laden” bone cysts [84,89]. BMP7 in particular (otherwise known as OP-1, which is also approved for human use for bone tissue regeneration) has been associated with adipogenic differentiation [91 –93]. These observations bring up troubling questions regarding the lack of specificity of BMPs for skeletal tissue regeneration, but also whether BMPs may be appropriate induction agents for in vitro adipogenesis. In essence, should BMPs be standardly supplemented to adipogenic differentiation medium? BMP-induced adipogenesis, however, is a relatively newly described phenomenon and may have as-yet undescribed, potentially biologically relevant differences from so-called “standard” adipogenic differentiation. Thus, we would extend caution to those supplementing BMPs to “standardized” adipogenic medium, unless specifically studying this interesting phenomenon in cell signaling.
Discussion
In summary, while the use of in vitro adipogenic differentiation of MSCs has increased in recent years, a lack of clear standardization is clear from the present review. Overall, improved standardization of basic laboratory techniques such as adipogenic differentiation will vastly improve the interpublication comparability. In examining the averages and medians of adipogenic induction medium, we suggest the following formulas (see Tables 2 –6 and 7 for a summary). Noteably, these are based on a compromise between all available techniques for the past year and not the authors' current laboratory practices.
ASCs, adipose-derived mesenchymal stem cells; BMSCs, bone marrow mesenchymal stem cells.
For 3T3-L1 cells, 1,000 nM insulin, 700 nM dexamethasone, and 500 μM IBMX are used. For mBMSCs, 5,000 nM insulin and 100 nM dexamethasone are used. For hBMSCs, 175 nM dexamethasone and 50 μM indomethacin are used. For mASCs, 320 nM insulin and 1,000 nM dexamethasone are used. For human ASCs, 393 nM insulin and 100 nM dexamethasone are used. All induction components should include 10% fetal bovine serum and no other components unless specifically being tested. Although no single recipe is the definitive “cocktail” for adipogenic differentiation, we suggest these concentrations as a reasonable starting point for new experiments. Such attempts at standardization will improve interlaboratory comparisons.
At least for primary MSCs, a certain degree of heterogeneity in adipogenic induction supplements is understandable—as in fact there is still debate about the exact identity of MSCs. For example, the stem cell surface markers characteristic of ASCs are still being examined, and so a precise identity and purity of these cell populations are still forthcoming. Despite our evolving definition of an MSC, the clear lack of standardization of adipogenic differentiation is quite apparent based on our review. To this end, we propose the aforementioned standardized components, which is a compromise based on available studies. Importantly, these adipogenic differentiation conditions represent by no means an ideal or maximal stimulation condition, but rather a simple average of recently published articles. Thus, these values have an essential arbitrary nature to them and should be simply used as a “starting-off” point rather than a “gold standard.”
One additional surprising result from our study besides the clear interlaboratory variation was the difference in adipogenic stimuli used for mouse and human cells (Fig. 3). For example, a 10-fold difference in dexamethasone concentration was observed between mouse and human ASCs (Fig. 3B). Such a difference could be in part anticipated, as species of derivation seems to impart basic biologic differences onto ASCs. For example, we have previously observed that mouse and human ASCs differ significantly in their ability to differentiate down an osteogenic lineage, both in vitro and in vivo [94 –96]. In addition, cytokine responsiveness seems to differ as well. For example, transforming growth factor-β1 appears to repress osteogenic differentiation in mouse ASCs; however, it has a significantly muted effect among human cells [96]. Such observations are indeed curious, and the basic interspecies differences among MSC populations have yet to be identified.
Footnotes
Acknowledgments
The authors thank Ms. Donna Soofer and Asal Askarinam for their helpful assistance. Benjamin Levi was supported by the National Institutes of Health, National Institute of Arthritis and Musculoskeletal and Skin Diseases grant 1F32AR057302-02. AWJ was supported by T32 grant number 5T32DE007296-14.
Author Disclosure Statement
The authors have no conflicts of interest.