
Abstract
Adipose-derived stromal cells (ADSCs) are close relatives of bone marrow mesenchymal stromal cells (BM-MSCs). The ease of access to subcutaneous fat pad and the abundance of stromal precursors make fat tissue an attractive source of stromal cells for clinicians. However, their ability to support hematopoietic stem cells in vitro and in vivo has not been established definitively. Thus, their usefulness in supporting hematopoietic stem cell engraftment is not as clear as with BM-MSCs. In this article, we show that human ADSCs, cultured with a good manufacturing practice medium, maintain in vitro human early and committed hematopoietic progenitors and support their complete differentiation toward myeloid and lymphoid lineages. Compared with BM-MSCs, ADSCs elicit a more precocious early progenitor formation and faster proliferation and differentiation of hematopoietic progenitors. Further, in vivo, when co-injected in NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ (NSG) mice with a low number of human CD34+ cells, ADSCs enabled the higher production of immature human hematopoietic progenitors and CD45+ cells when compared with BM-MSCs. As a whole, our results indicate that human ADSCs, isolated and expanded under clinical-grade conditions, support hematopoiesis in vitro and in vivo and thus provide the rationale for their use in supporting hematopoietic reconstitution in clinical settings.
Introduction
T
More recently, Wagner et al. found that the ability to maintain human primitive hematopoietic progenitors was higher for MSCs from BM, compared with stromal cells from adipose tissue (AT) [4]. Similarly, Kilroy et al. [5] observed that cocultures with ADSCs produced half as many CD34+ cells than those observed with MSCs. On the contrary, Nakao et al. demonstrated in a heterologous system that co-injection of murine ADSCs, but not murine MSCs, with human CD34+ peripheral blood stem cells improved the survival of irradiated C57/Bl6 mice [6]. Using a few samples from patients and one in vivo experiment, Kim et al. reported that human ADSCs and MSCs similarly promote engraftment of CD34+ in NOD/SCID mice [7]. Of note, these studies were restricted to either in vitro or in vivo studies or based upon a limited number of experiments.
In the present work, using a large series of in vitro and in vivo experiments, we provide evidence that human adipose cells obtained after isolation and expansion performed using good manufacturing practice (GMP) media and reagents—a key step before pretherapeutic studies—support efficient human hematopoiesis.
Materials and Methods
Tissue preparations
Samples were obtained from fully informed hospital patients, according to French bioethics rules.
Subcutaneous AT samples were obtained from 3 men and 16 women (aged 30–81 years) undergoing elective abdominal dermolipectomy or liposuction. Stromal vascular fraction (SVF) was prepared as previously described [8]. Then, the AT was briefly digested with 2 mg/mL collagenase A (Roche Diagnostic) in 2% albumin containing DMEM:F12 medium (Invitrogen) for 45 min at 37°C, filtered through a 100-μm and then a 25-μm nylon mesh, and centrifuged at 600 g for 10 min to separate floating mature adipocytes from SVF in the pellet.
BM specimens were obtained from 9 men and 11 women (18–81 years) who underwent total hip prosthesis procedures, by milling the spongy bone from the femoral cavity. Marrow cells were extracted by high agitation in phosphate-buffered saline (PBS). Cell suspension was centrifuged 600 g for 10 min, the supernatant was discarded, and the pellet was washed with PBS.
Isolation and culture of stromal cells
Except for CFU-F assays, SVF cells were seeded (0.75×105 cells/mL medium) in plastic flasks and cultured at 37°C in a humidified atmosphere of 5% CO2. The GMP medium was α-MEM medium (Invitrogen) containing human plasma (EFS Midi-Pyrénées; enriched with human platelet factors), 0.2 mM
Except for CFU-F assays, BM mononuclear cells were seeded (3.75×105 cells/mL medium) and cultured in the same medium and under the same conditions as those used for SVF cells until confluence or up to a maximum of 21 days.
These isolation and culture procedures could be easily adapted to full GMP procedures.
Surface phenotypic analysis
Human BM-MSCs, human ADSCs, human hematopoietic cells from long-term culture (LTC), and freshly isolated mouse BM-derived cells were stained in PBS containing FcR Block reagent. Cell viability was tested using a viability marker (Live Dead; Invitrogen). Quintuple or sextuple staining was performed by direct immunoflorescence with conjugated mouse anti-human monoclonal antibodies when compared with their matched isotype control human monoclonal antibodies (BD Biosciences). Cells were washed in PBS and analyzed on a flow cytometer (FACS Canto II; Becton Dickinson). Data acquisition and analysis were performed using FACS Diva software (Becton Dickinson).
The various combinations of antibodies and conjugates used for in vivo and in vitro experiments are listed in Table 1.
Antibodies obtained from BD Biosciences, San Jose, CA.
Antibodies obtained from eBiosciences, San Diego, CA.
Antibodies obtained from Beckman Coulter, Fullerton, CA.
FITC, fluorescein isothiocyanate; PE, phycoerythrin; PerCP, peridinin chlorophyll protein; PE-Cy7, phycoerythrin cyanine 7; APC, allophycocyanin; APC-Cy7, allophycocyanin cyanine 7; BM, bone marrow; AT, adipose tissue.
CFU-F assay
To evaluate the frequency of mesenchymal-like progenitors in the human SVF, cells were cultured in T-25 flasks at a final concentration of 16 cells/cm2 in the GMP medium described earlier. The medium was renewed every 2 or 3 days. After 10 days, the cells were gently washed with PBS and fixed with methanol for 15 min. For scoring the CFU-F, flasks were stained with Giemsa 6% for 30 min. Colonies consisting of aggregates of at least 50 cells were scored under an optical microscope.
To evaluate the frequency of mesenchymal-like progenitors in human BM, cells were cultured in T-25 flasks at a final concentration of 4×104 cells/cm2 in GMP medium described earlier. The CFU-F assay was performed as described.
Adipogenic and osteogenic differentiation
Adipogenic differentiation was performed when SVF or BM cells reached confluence in a culture medium with 10–3 mM dexamethasone, 450 μM iso-butyl-methyl-xanthine, and 60 μM indomethacin (Sigma) over 10 days. The medium was changed every 2 or 3 days. Then, IBMX was removed from the medium and differentiation was extended over 11 days. The extent of differentiation was noted by observation of multilocular refractive droplets in the induced cells, staining of neutral lipids by Oil Red O, and determination of triglycerides, as previously described [9]. Differentiation was quantified by cellular triglyceride content measurement by using a commercially available test combination (Triglycerides enzymatiques PAP 150; Biomerieux) after cell lysis in 0.1 N NaOH. The triglyceride content was calculated per microgram of proteins. Protein content was determined using the DC Protein Assay Kit (BioRad).
Osteogenic cell differentiation was performed under similar conditions in a medium with 10–4 mM dexamethasone, 250 μM ascorbic acid, and 3×10–3 mM NaH2PO4 − and assessed by staining of calcium-rich deposits by Alizarin Red [10]. To obtain quantitative data, 10% acetic acid was added to the stained culture dish and optical density was measured by a spectrophotometer at 405 nm.
Isolation of hematopoietic progenitors
Mononuclear cells were isolated after centrifugation of marrow cord-blood cell suspension on a Ficoll-Paque gradient (StemCell Technologies). CD34+ hematopoietic progenitors were enriched by magnetic-activated cell sorting (CD34 Microbeads Kit; Miltenyi Biotec) with a mean purity of 95.95%±1.54%, viability of 92.5%±4.3%, and yield of 36.4%±6.6%. The morphology of hematopoietic progenitors was appraised on May-Grünwald-Giemsa (MGG)–stained cytospins.
Long-term coculture, proliferation, and differentiation of CD34+ cells
Cocultures were based on the LTC method of Hogge et al. [11] for human BM, in which a stromal layer (containing a variety of nonhematopoietic cells) supports the growth and development of hematopoietic cells for 5–8 weeks. This system is considered to be the best culture system for monitoring the growth and development of immature hematopoietic progenitor cells capable of producing clonogenic cells and mature cell progeny [12,13].
Here, 2×104 CD34+ hematopoietic progenitors were seeded onto confluent adherent feeder layers (BM-MSCs or ADSCs) grown for 15 or 35 days at 33°C. These cocultures were carried out in LTC medium (Myelocult; StemCell Technologies; supplemented with 10−6 M hydrocortisone), optimal for hematopoietic development [11], and fed weekly with half-medium changes.
Nonadherent hematopoietic cells were collected every week and counted using Trypan Blue exclusion to determine the impact of the feeder layers on hematopoietic cell proliferation. In parallel, the proportions of CD34+ cells (corresponding to hematopoietic stem plus progenitor cells) and CD34+/CD38− cells [corresponding to more primitive hematopoietic stem cells (HSCs)] were determined among CD45+ LTC cells by flow cytometry.
Clonogenic assays in methylcellulose (Methocult GF+ H4435; StemCell Technologies) were performed at 2 and 5 weeks of culture to evaluate the frequency of colony-forming units–granulocyte macrophage (CFU-GM), colony-forming units–erythroid (CFU-E), and burst-forming units–erythroid (BFU-E). In the methylcellulose assay, clonogenic cells were scored in situ at 14 days as CFU-GM, BFU/CFU-E, and total CFCs (colony-forming cells). The morphology of different CFCs from 2 weeks of LTC was appraised on MGG-stained cytospins.
Depending on the hematopoietic lineage tested, cytokines (AbD Serotec) were added to the LTC medium after 2 weeks of classic LTC: erythropoietin (2 U/mL), thrombopoietin (50 ng/mL), interleukin (IL)-7 (10 ng/mL), or IL-15 (10 U/mL). After 15 days at 33°C, the morphology of nonadherent differentiating hematopoietic cells was appraised on MGG-stained cytospins. In parallel, different phenotypes of differentiated hematopoietic cells were determined in the CD45+ cell population of LTC by flow cytometry, using a panel of antibodies (Table 1) directed against monocytes, granulocytes, megakaryocytes, erythroblasts, NK cells, and B and T lymphocytes.
LTC-initiating cell assay
LTC-initiating cell (LTC-IC) frequency [13] was assessed with purified CD34+ cells cultivated upon the 2 feeder layers. Confluent layers of BM-MSCs or ADSCs were seeded in 96-well plates. CD34+ cells were plated in limiting dilutions (12, 36, and 60 replicates, respectively, per concentration: 100, 50, and 10 cells per well) on these feeder layers and cultured in LTC medium at 33°C. After 5 weeks, the cells were overlaid with clonogenic methylcellulose medium (Methocult GF+ H4435). Cultures were scored for colony-forming cells after an additional 2 weeks of growth at 37°C. Four and 6 independent experiments were performed with MSCs and ADSCs, respectively.
In vivo transplantations
NOD.Cg-Prkdc(scid) Il2rg(tm1Wjll)/SzJ (NSG) (Jackson Laboratory) immune-deficient mice were housed in pathogen-free animal facilities at the CEA (Fontenay-aux-Roses, France). All experimental procedures were performed in compliance with French Ministry of Agriculture regulations (animal facility registration No. A920322) for animal experimentation.
Adult 8-week-old NSG mice were irradiated at 3 Gy and anesthetized with isoflurane before intravenous retro-orbital injection of 5×103 viable cord blood CD34+ cells in the presence or absence of 5×105 allogeneic MSCs or ADSCs. Three weeks after transplantation, a BM aspiration was performed as a first measure of the engraftment of human CD45+ hematopoietic cells. Eleven weeks after grafting, mice were killed by carbon dioxide inhalation. BM was collected by flushing both femurs with RPMI medium (Invitrogen). Human cell engraftment was detected by flow cytometry using human CD45 antibody. Grafted hematopoietic progenitor cells (CD34+/CD38−) were assessed by flow cytometry in CD45+ Lin− cells. Phenotypes of differentiated grafted hematopoietic cells were determined by flow cytometry using a panel of antibodies (Table 1) directed against B and T lymphocytes, monocytes, and neutrophils.
Data analysis
Results were expressed as means±SEM or medians±SEM for in vivo experiments. Comparisons between groups were conducted using the 2-tailed unpaired t-tests for in vitro experiments and using the 2-tailed Mann–Whitney tests for in vivo experiments, using GraphPad Prism 4.0 software (GraphPad Software). Significance was defined as *P≤0.05 and **P≤0.01.
Results
MSC and ADSC characterization
We evaluated the capacity of human BM and subcutaneous AT to generate mesenchymal progenitors, using the CFU-F assay (Fig. 1A). The number of CFU-F was significantly higher in SVF (64,375±16,563 equivalent CFU-F for 1×106 SVF cells seeded) compared with BM (53±16 equivalent CFU-F for 1×106 BM cells seeded), confirming that human AT is a major source of stromal progenitors.
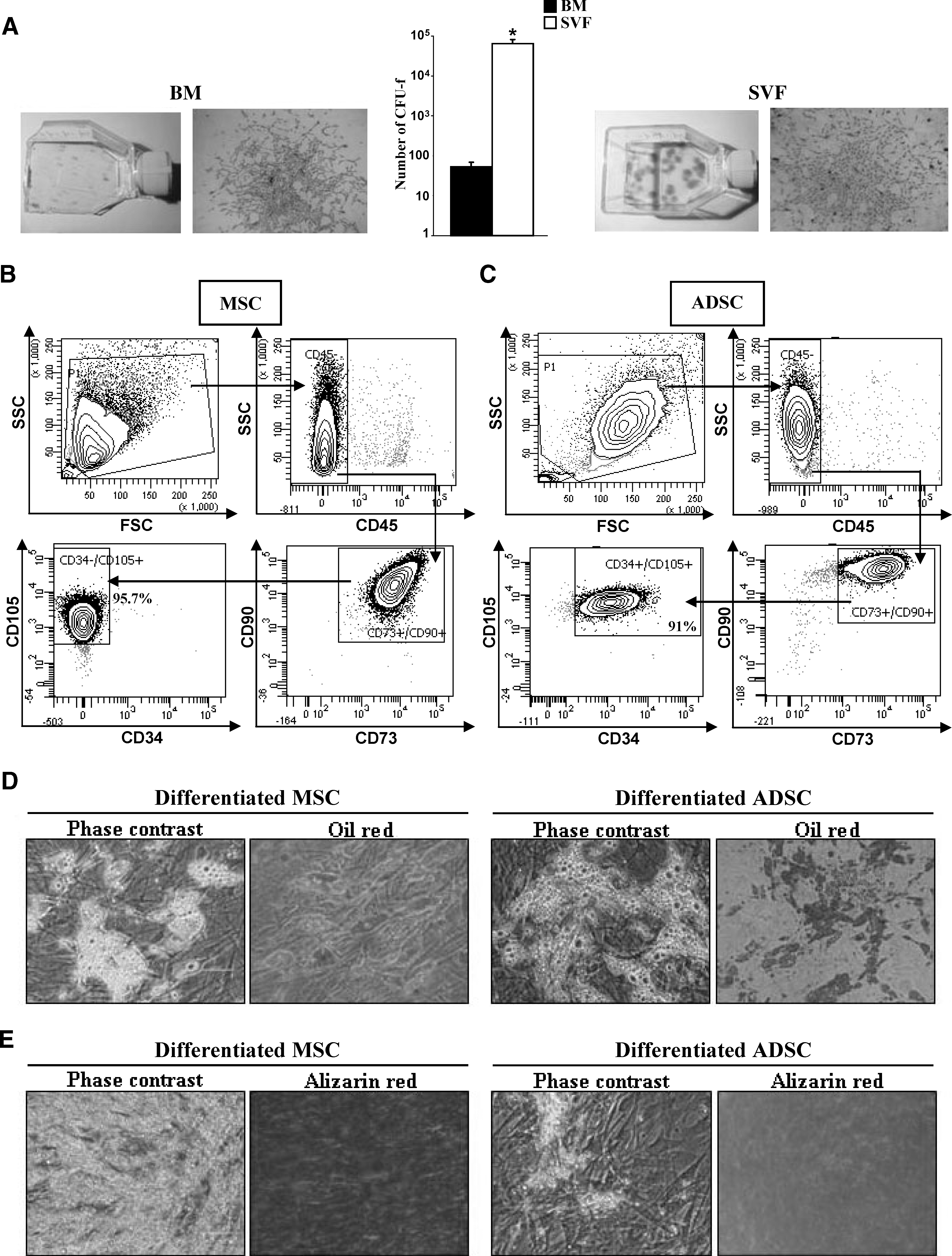
Mesenchymal stromal cells (MSCs) from bone marrow (BM) or adipose stromal vascular fraction. CFU-F quantification: Giemsa staining; n=4; *P≤0.05
Previous studies have suggested that although stromal MSCs and ADSCs are very similar, they do display important differences. Flow cytometry analysis showed that human MSCs and ADSCs, cultured in human plasma, shared some cell surface markers, such as CD105, CD90, and CD73, and were negative for CD45, as expected for nonhematopoietic cells, but differed in the expression of CD34. Indeed, MSCs were mainly CD34−/CD45−/CD105+/CD90+/CD73+ (96.9%±0.6%; Fig. 1B), whereas ADSCs were CD34+/CD45−/CD105+/CD90+/CD73+ (88.1%±1.9%; Fig. 1C).
Next, we evaluated adipogenic and osteogenic potentials in both cell types. Lipid droplets and Oil Red O staining were not observed in undifferentiated MSCs or ADSCs because consistent cytoplasmic vacuolation was evident in the induced cells (Fig. 1D). The quantification of triglyceride content (Supplementary Fig. S1A; Supplementary Data are available online at
During osteogenic differentiation, morphological changes appeared during the second week of subculture. At the end of the 21-day induction period, some calcium crystals were visible in the culture. Cell differentiation was confirmed by Alizarin Red staining (Fig. 1E) and quantification (Supplementary Fig. S1B), revealing higher levels in MSCs, as previously reported [14].
Early hematopoietic progenitors
CD34+ cells were isolated from BM with a mean purity of >95%, as determined by flow cytometry analysis (Fig. 2A) and MGG staining. They were cultured with stromal cells in LTC conditions.
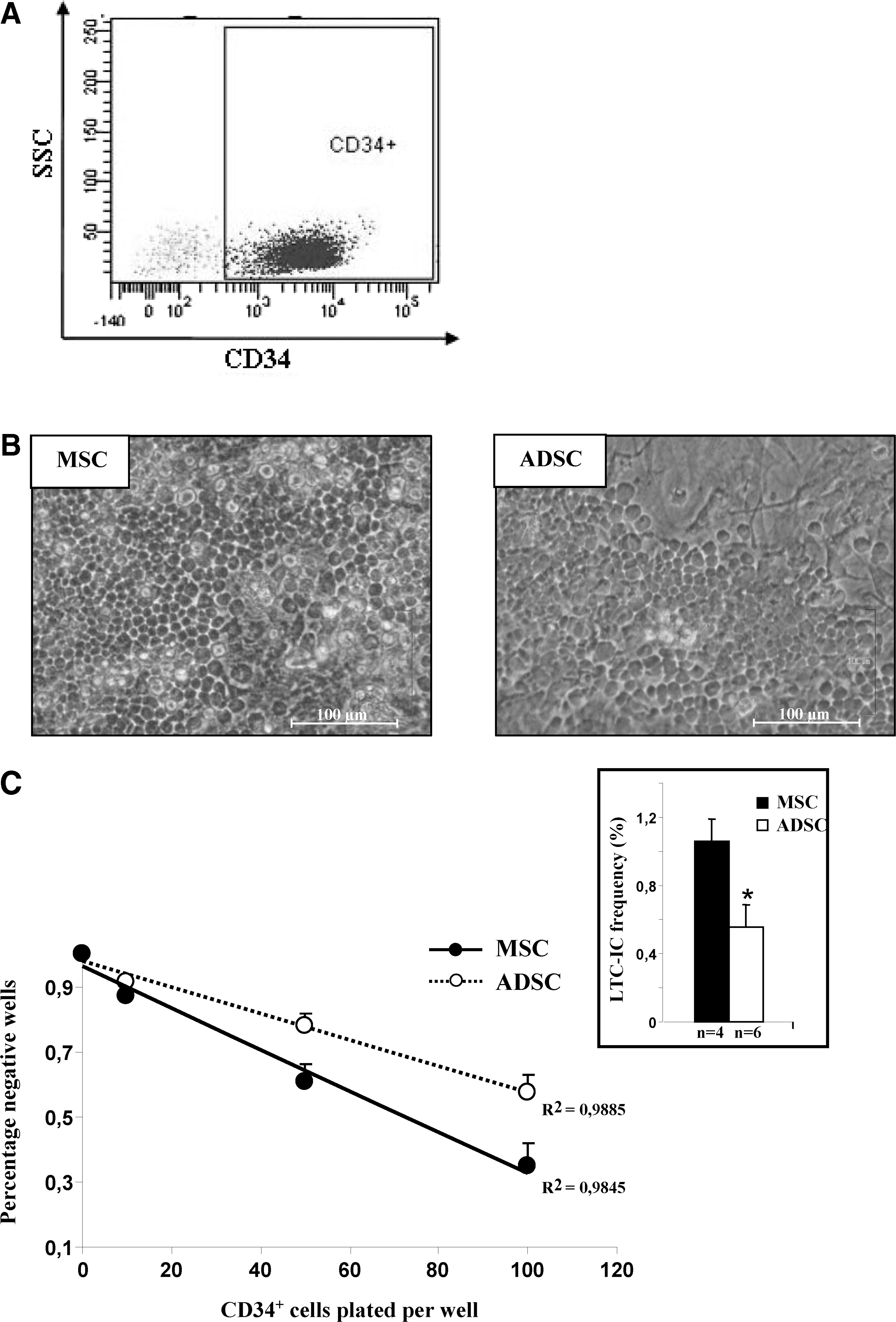
Characterization of CD34+ hematopoietic progenitors and analysis of early hematopoietic progenitors. Phenotypic analysis of viable CD34+ cells isolated from BM (representative experiment with 95.4% purity)
The formation of a cobblestone area (Fig. 2B) was observed earlier (day 14) during LTC performed with ADSC layers than with MSC layers (days 21–28).
Quantitative evaluation of very immature hematopoietic progenitors (LTC-IC) was performed using LTCs in limiting dilution conditions. The frequency of LTC-IC was about half as great with ADSC feeder layer as with MSCs after 35 days of culture (Fig. 2C; 0.56% vs. 1.06%). These results show that if ADSCs are able to maintain long-term survival of very immature hematopoietic cells, MSCs exhibit a higher capacity.
We followed up the proportions of CD34+ and CD34+ CD38− cells in the whole hematopoietic CD45+ population during the first 3 weeks of culture. Our results show that the proportions of hematopoietic CD34+ (Fig. 3A) or CD34+CD38− (Fig. 3B) cells were significantly higher in the presence of MSCs than with ADSCs at 7, 14, and 21 days of culture. The proliferation of hematopoietic cells was increased on the ADSC layer compared with MSCs during the first 14 days of culture (Fig. 3C). In the same way, after 21 days of culture, the hematopoietic cell proliferation on the ADSC layer seemed higher than on MSCs. This would indicate a faster differentiation of HSCs into committed progenitors on ADSCs than MSCs.
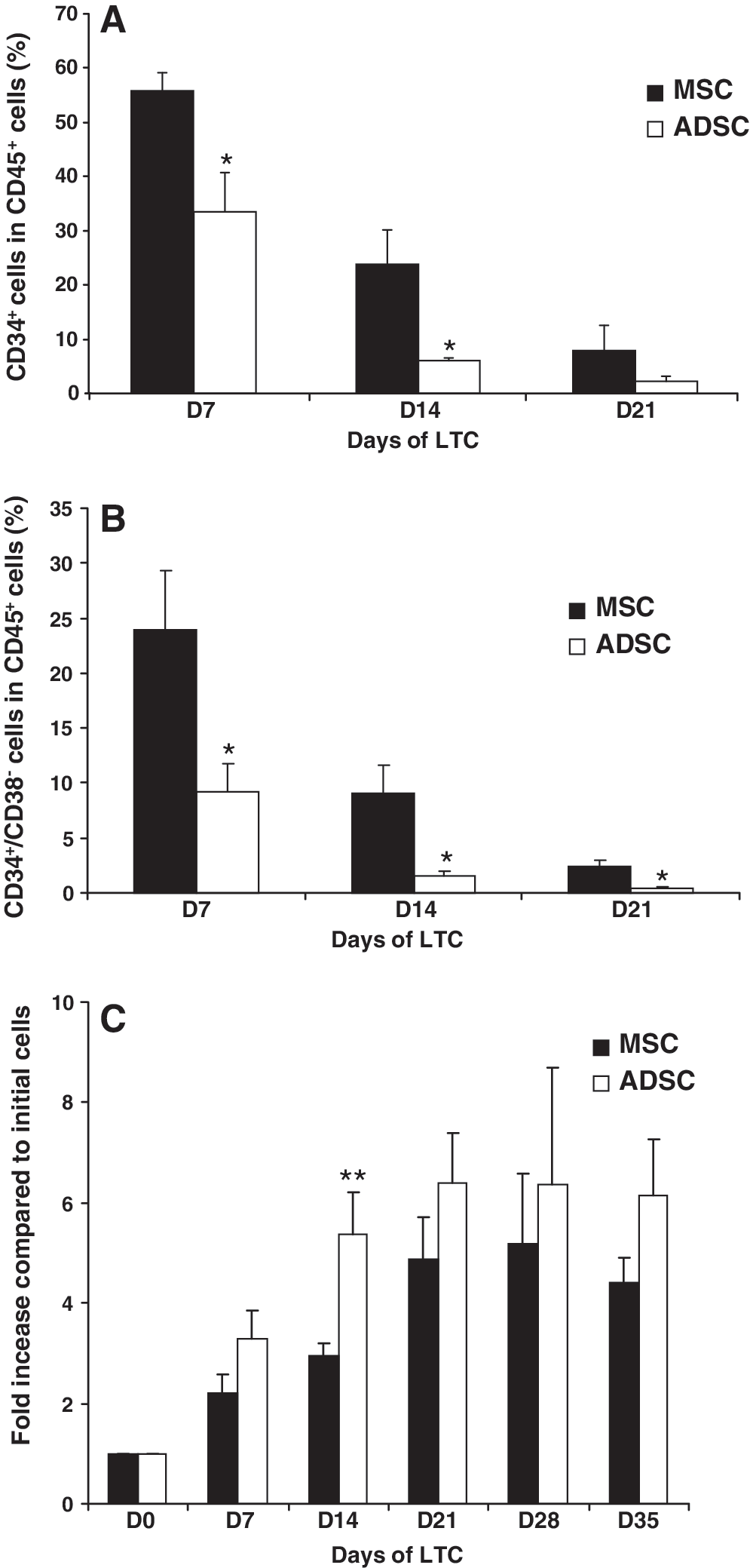
Proliferation of CD34+ hematopoietic progenitors. Proportion of CD34+
Hematopoietic-committed progenitors and precursors
Human CD34+ cells were seeded onto ADSC or MSC layers for 2 or 5 weeks. Then, the cells were cultured in semisolid methylcellulose for 15 days to generate hematopoietic colonies (Fig. 4A). After 2 weeks of LTC, a higher number of CFU-GM was obtained with the ADSC layer (286.19±67.7 vs. 447.56±86.25), the opposite being observed after 5 weeks (48.8±11.18 vs. 26.82±6.3; Fig. 4B). A similar number of erythroblast colonies (BFU-E plus CFU-E) was observed after 2 weeks of culture either on the ADSC or MSC layer, but it was higher with the MSC layer after 5 weeks of culture (Fig. 4C). A similar pattern was observed when pooling the different types of hematopoietic colonies (Fig. 4D).
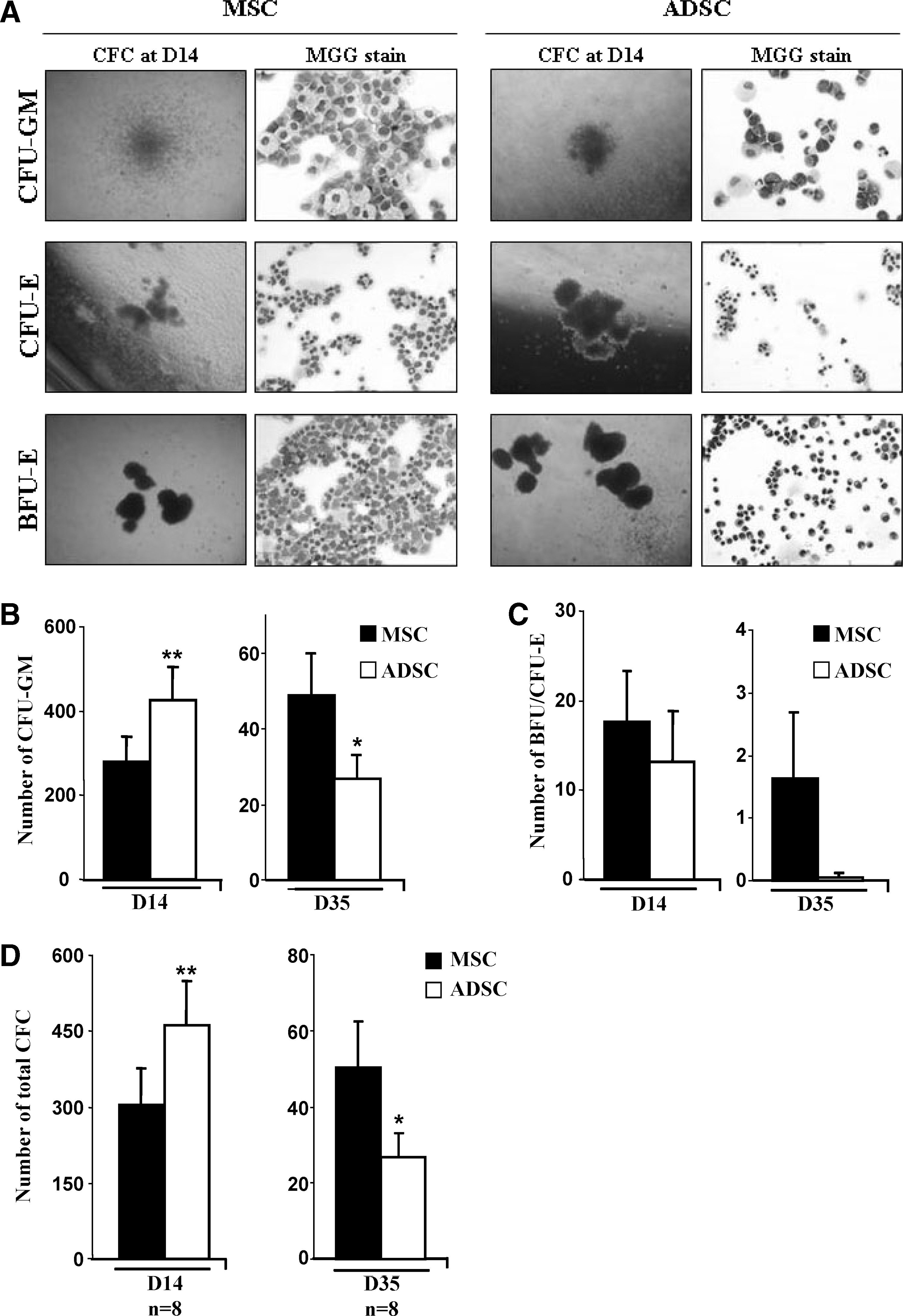
Maintenance of hematopoietic-committed progenitors and precursors on MSCs or ADSCs. Clonogenic tests in methylcellulose. Formation and May-Grünwald-Giemsa staining of CFU-GM, CFU-E, and BFU-E clusters at day 14
These results could indicate that human ADSCs are efficient for maintaining committed progenitors and that MSCs are more effective at maintaining earlier progenitors.
Differentiation of CD34+ cells toward myeloid and lymphoid lineages
Using clinical-grade conditions, we confirmed our previous results concerning the differentiation of CD34+ cells cultured on human ADSC layers [3]: human hematopoietic progenitors can differentiate into granulocytes, monocyte-macrophages, megakaryocytes, erythroblasts, and lymphocytes as assessed by MGG staining and flow cytometry analysis. Differentiated cells (ie, segmented neutrophils, monocytes, lymphocytes, orthochromatic erythroblasts, and megakaryocytes) were obtained earlier on the ADSC layers than on the MSC layers. Feeder layers of MSCs or ADSCs did not generate human hematopoietic cells when cultured in the absence of CD34+ cells. Moreover, in addition to our previous observations [3], we found that CD34+ cells cultured on the ADSC layers could differentiate into not only B and NK but also T lymphocytes as assessed by the presence of CD19, CD56, and CD3, respectively (Fig. 5).
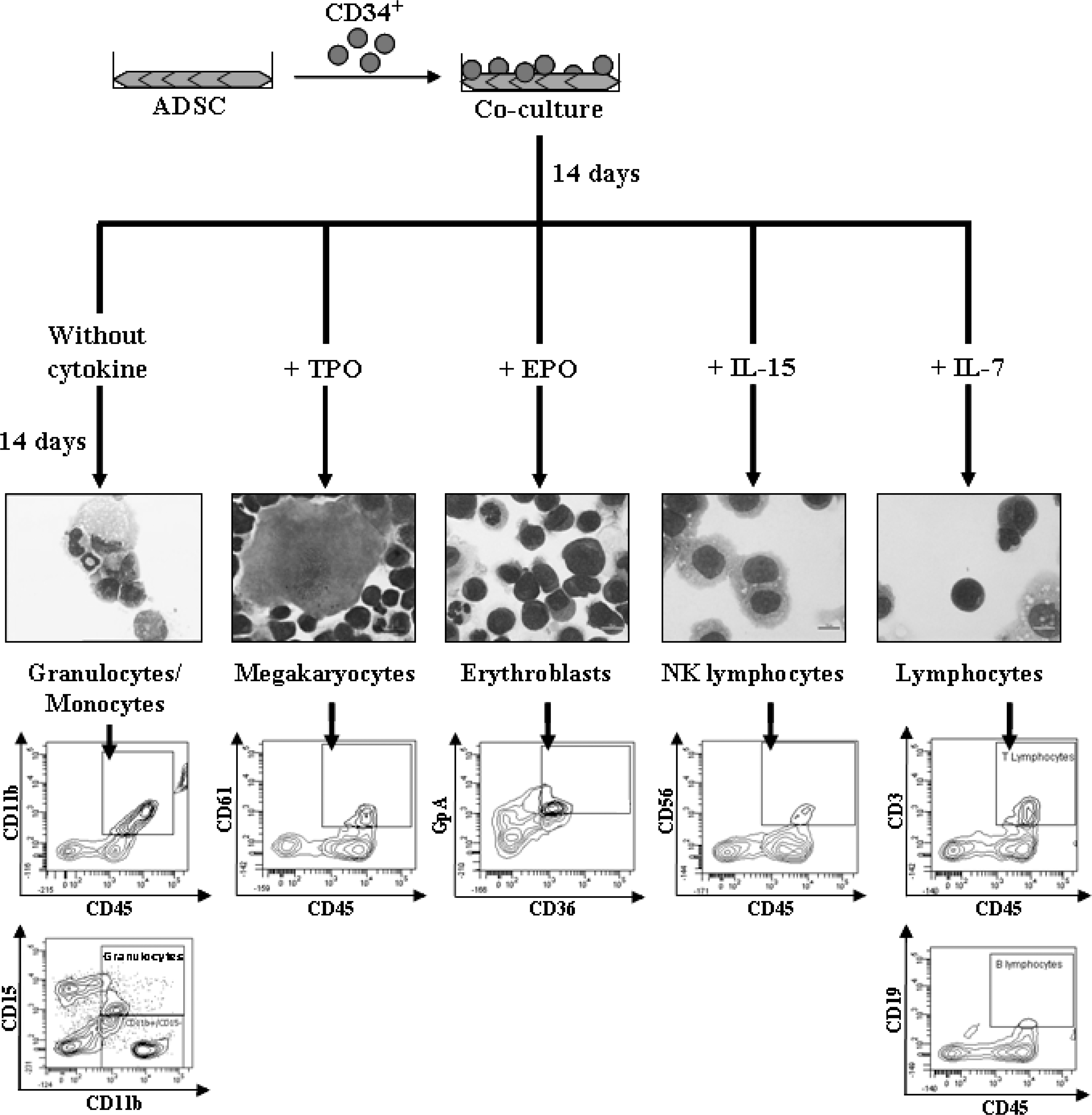
Differentiation of CD34+ in myeloid and lymphoid lineages: May-Grünwald-Giemsa staining and surface phenotype by flow cytometry analysis.
Hematopoietic reconstitution in NSG mice
To assay whether ADSC support was beneficial to human hematopoietic reconstitution after transplantation, CD34+ cord-blood cells were injected into immune-deficient (NSG) mice with human ADSCs or MSCs or alone (Fig. 6A). As shown in Fig. 6B, 3 weeks after transplantation, the number of human CD45+ cells was significantly higher in the BM of mice cografted with CD34+ cells and ADSCs (CD34++ADSCs: 1.6% vs. CD34++MSCs: 0.3% and CD34+ alone: 0.26%). This difference in hematopoietic recovery was no longer observed at 11 weeks after the infusion of the cells into 3 different groups of mice (CD34++MSCs: 4.4%; CD34++ADSCs: 7%; CD34+ alone: 2.8%). However, immature human hematopoietic CD34+/CD38− progenitors were more frequent in human Lin−/CD45+ population cells (2.5% vs. 0.55%; P=0.0927) in mice injected with CD34+ cord-blood cells and ADSCs (Fig. 6C). Conversely, we observed no significant difference in the proportion of human differentiated hematopoietic cells (Fig. 6D, E).
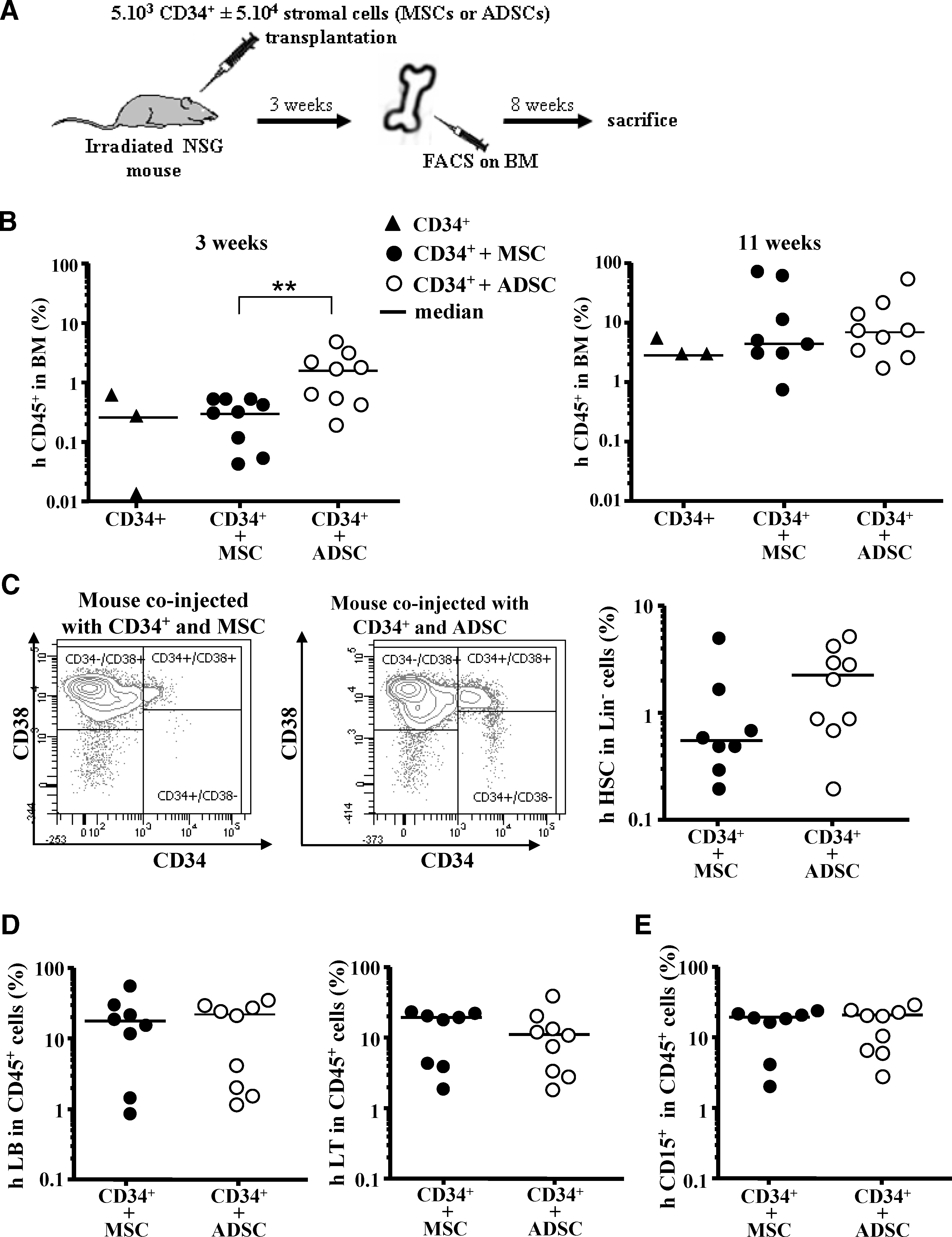
In vivo studies. Grafting protocol
Altogether, these results suggest that ADSCs are at least as efficient as MSCs at promoting the grafting and in vivo production of human hematopoiesis in immune-deficient mice.
Discussion
We have described here, for the first time, the in vitro and in vivo hematopoietic-supporting activity of human ADSCs cultured in clinical-grade media.
Isolation procedures for stromal cells vary extensively, and this may influence their properties. It has been reported that culture conditions have a major impact not only on the composition of human stromal cell preparations [15] but also on the proteome of cultured cells [4,16]. Extended culture of MSCs alters their capacity to support hematopoietic progenitor cells without a change in their phenotype or differentiation ability [17]. The need for accurate and standardized isolation procedures seems obvious for a clinical utilization.
In many reported clinical trials, human MSCs are expanded in fetal bovine serum (FBS). However, FBS may elicit an immune response, particularly in patients receiving repeated cell infusions [18,19]. Therefore, replacing FBS with human serum or plasma appears a prerequisite to safer therapeutic assays. Conflicting results have been reported for MSCs cultured in human serum [20 –22], concerning proliferation or differentiation properties. Here, we observed no alteration of proliferation, phenotype, and differentiation properties of MSCs and ADSCs after expansion in human plasma (data not shown).
Concerning our own results, modifications to CD34+ culture conditions probably explain the differences between present and previous works [3], wherein we observed neither the generation of T lymphocytes from CD34+ cells nor the maintenance of hematopoietic progenitors seeded onto ADSCs.
A number of publications demonstrate that cotransplantation of human stromal cells favors engraftment of hematopoietic progenitors. Generally, unrelated MSCs isolated from BM have been used with CD34+ cells obtained directly or mobilized from marrow or from umbilical-cord blood units [23 –30]. This raises the problem of availability of MSCs as a source for routine utilization: leftover material from total hip arthroplasty or iliac crest aspirate from healthy donors giving BM for allogeneic transplantation purpose. Noort et al. [31] used culture-expanded human fetal lung cells, but the limited access to such material and ethical concerns exclude this source of stromal cells for therapeutic studies. In this respect, subcutaneous AT, easily accessible by liposuction and particularly rich in stromal progenitors, is an attractive candidate for reconstructive therapy.
Few publications are available concerning the hematopoietic-supporting activity of human ADSCs. The closest data to our own results were reported by Kim et al. [7]. They found that human ADSCs expanded in 10% human serum maintain their proliferative capacity and ability for differentiation into adipocytes and osteoblasts. These authors did not perform in vitro progenitor assays. In irradiated NOD/SCID mice, they established that ADSCs and MSCs promote a similar engraftment of mobilized human hematopoietic progenitors at 6 weeks after transplantation. Using fewer CD34+ cells (5×103 vs. 5×105) and fewer ADSCs (5×105 vs. 106), we also observed an equivalent percentage of human CD45+ hematopoietic cells in the BM of transplanted mice as previously described [32]. Here, we also demonstrate the persistence of human multilineage engraftment at 11 weeks after infusion.
For cotransplantation studies, we used retro-orbital injections. This method prevents the cell collapse that is frequently observed in the heart when cells are infused via tail vein [33]. It has also been described that MSCs could be trapped in the lung following intravenous injection. Further studies will be necessary to track the distribution of ADSCs in different tissues after retro-orbital injection.
Compared with their BM counterpart, MSCs, we found that human ADSCs elicit a more precocious formation and proliferation of early progenitors (cells forming cobblestone areas and LTC-IC), even if their number is lower. We also observed an earlier generation of differentiated blood cells. These observations are in line with the results obtained by Wagner et al. [4] with a different experimental design (hematopoietic progenitor cells isolated from umbilical cord blood and culture medium supplemented with 10% fetal calf serum).
The ability of ADSCs to allow rapid ex vivo expansion and differentiation of CD34+ cells could be valuable to quickly obtain a large number of committed progenitors for short-term hematopoietic reconstitution.
Taken together, our in vitro and in vivo results indicate that human ADSCs elicit an important proliferation of HSCs with rapid generation of committed hematopoietic progenitors. These progenitors are maintained for a long time in the grafted animals and give rise to different blood lineages. Moreover, efficient engraftment was obtained with a very low number of injected CD34+ cells. From a clinical perspective, these properties could be very valuable in stimulating a rapid hematopoietic recovery following BM transplant.
Several mechanisms could explain such an efficiency of ADSCs in supporting hematopoiesis. It has been shown that human ADSCs secrete various adipokines and cytokines, among them the hematopoietic factors IL-6, IL-7, M-CSF, G-CSF, and GM-CSF. However, the cytokine profile of ADSCs is similar to that reported for human BM-derived MSCs [5]. Therefore, it would be worthwhile to look for differences between adhesive cell surface molecules [34] or factors involved in HSC self-renewal and maintaining stemness such as components of the Wnt/β-catenin pathway [35].
As a whole, our results indicate that human ADSCs isolated and expanded in GMP media and reagents fully and efficiently support hematopoiesis in vitro and in vivo. ADSCs can therefore be used as an alternative to BM MSCs for the study of human hematopoiesis. Moreover, our data provide a rationale for the utilization of ADSCs in phase I clinical trials.
Footnotes
Acknowledgments
The authors thank Corinne Barreau (UMR STROMALab, Toulouse, France) for excellent assistance in FACS and Mireille André and Pascale Guillou (UMR 5241, Toulouse, France) for excellent technical participation for in vivo and in vitro experiments. The authors are grateful to Pr. P. Chiron and Dr. J.L. Tricoire (CHU Rangueil, Toulouse, France) for providing human bone marrow and Pr. J.P. Chavoin (CHU Rangueil, Toulouse, France) for providing human fat from liposuctions. This work was supported by the Etablissement Français du Sang (EFS).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.