
Abstract
An ongoing debate surrounds the existence of stem cells in the adult endowed with capacity to differentiate into multiple lineages. We examined the possibility that adult bone marrow cells participate in recovery from chemical diabetes through neogenesis of insulin-producing cells. Small-sized cells negative for lineage markers derived by counterflow centrifugal elutriation from the bone marrow were transplanted into mice made diabetic with streptozotocin and sublethal irradiation. These cells homed efficiently to the injured islets and contributed to tissue revascularization. Islet-homed CD45-negative donor cells identified by sex chromosomes downregulated GFP, expressed PDX-1 and proinsulin, and converted the hormone precursor to insulin. An estimated 7.6% contribution of newly formed insulin-producing cells to islet cellularity increased serum insulin and stabilized glycemic control starting at 5 weeks post-transplant and persisting for 20 weeks. Newly differentiated cells displayed normal diploid genotype and there was no evidence of fusion between the grafted stem cells or their myeloid progeny and injured β-cells. Considering the extensive functional incorporation of insulin-producing donor cells in the injured islets, we conclude that the adult bone marrow contains a subset of small cells endowed with plastic developmental capacity.
Introduction
T
In type 1 diabetes it is essential to abrogate the autoimmune reaction that threatens any cell type with phenotypic characteristics (antigenic targets) of β-cells before attempts to regenerate the tissue [9]. Interventions aiming to reconstitute the islet mass have yielded controversial data so far. It is generally recognized that hematopoietic and mesenchymal stromal cells derived from the bone marrow and umbilical cord blood (UCB) exert a positive effect on islet recovery from chemical injury [10 –26]. The therapeutic effects have been attributed primarily to facilitation of β-cell recovery from chemical injury through support of β-cell viability and function [11,13,17,24], local immunomodulation [10,14,20,25], neovascularization [11,16,20], and local secretion of growth factors [18,22]. Insulin production has been sporadically observed in donor cells [10,11,19,26]; however, significant conversion of adult cells from hematopoietic compartments to produce insulin that would be of therapeutic value has been observed only in transplants of bone marrow cells (BMC) after myeloablative irradiation [12] and of UCB cells in neonate immunodeficient mice [27]. Data reported by these in vivo studies differ from the effective induction of insulin production in vitro, which was convincingly demonstrated for both adherent [28] and nonadherent fractions of BMC [29] and UCB cells [30,31]. If the capacity of differentiation is present in various cell types, negative reports might originate from suboptimal timing, detection technique, or selection of the subset of primitive cells derived from the bone marrow [32]. Progenitors derived from this compartment undertake a default and dominant hematopoietic differentiation trait under most experimental conditions [33].
One way to circumvent the predisposition of bone marrow-derived progenitors to reconstitute the immunohematopoietic system during acute stages of islet injury is the use of subsets that lack short-term hematopoietic activity [34,35]. The smallest elutriated fraction of the bone marrow has poor radioprotective capacity, but elaborates delayed and durable long-term reconstituting potential [34 –36]. This subset is endowed with the capacity to differentiate into multiple non-hematopoietic lineages [37], including liver [38], and single cells contribute widely to reconstitution of epithelial tissues [39]. In this study we examined the contribution of adult bone marrow-derived primitive cells to recovery from chemical diabetes through neogenesis of insulin-producing cells. Using an experimental approach designed according to our interpretation of the literature [9] and careful evaluation of the target tissue [32], we found that adult bone marrow-derived cells home efficiently to the injured islets, differentiate to produce insulin, and restore near-euglycemic levels through contribution of ∼8% to insulin-producing cells.
Materials and Methods
Animal preparation, diabetes, and transplantation
Mice used in this study were C57Bl/6J (B6, H2Kb, CD45.2), B6.SJL-Ptprca Pepcb/BoyJ (H2Kb, CD45.1), and C57BL/6-TgN(ACTbEGFP)1Osb (GFP, H2kb, CD45.2) purchased from Jackson Laboratories and housed in a barrier facility. All procedures were approved by the Institutional Animal Care Committee. Diabetes was induced in female mice (aged 6–8 weeks) by 5 daily consecutive intraperitoneal injections of 60 μg/g Streptozotocin (STZ; Calbiochem) [32]. STZ was diluted in phosphate-buffered saline (PBS; Beit Haemek) and was used within 15 min of preparation. Blood glucose levels were monitored with a standard glucometer (Accu-Chek Sensor; Roche Diagnostics) in mice fed ad libitum at constant daytime hours (9–11 AM). Diabetes was considered at glucose levels exceeding 250 mg/dL in 2 consecutive measurements. Glucose tolerance test was performed by intraperitoneal injection of 2 g glucose and blood levels measurements after 60 and 120 min. Before cell transplantation (day 0), recipients were sublethally irradiated at 675 rad (total body irradiation) using an X-ray irradiator (RadSource 2000) at a rate of 106 rad/min (day 1). Intravenous infusion of cells was performed in 0.2 mL PBS.
Cell isolation
Whole BMCs from wild-type or GFP-positive male donors were harvested by flushing of medullar cavities of femur, tibia, and iliac bones. Single cell suspensions (5×108 cells) were loaded into the chamber of a counterflow centrifuge (Beckman Instruments) operating at a constant speed of 3,000 rpm [40]. Fractions were collected in 200 mL at elutriation flow rate of 25 mL/min to isolate the smallest subset of nucleated cells (Fr25), and the largest cells were collected in the rotor off position. Fr25 cells were lineage-depleted by incubation at 4°C with rat-anti mouse monoclonal antibodies (mAb) against CD5 (clone 53-7.3), GR-1 (clone RB6-8C5), Mac-1 (clone M1/70), B220 (clone RA3-3A1/6.1) extracted from hybridoma cell lines (ATCC), and purified TER119 (eBioscience) [32]. Secondary goat-anti-rat antibodies conjugated to magnetic beads (Dynal Biotech) were used to trap cells coated with primary antibodies in a magnetic field as previously described [40].
Flow cytometry
Measurements were performed with a Vantage SE flow cytometer (Becton Dickinson) on cells that underwent red cell lysis. Briefly, cells were suspended in lysis buffer (4.15 g NH4Cl, 0.5 g KHCO3, and 0.15 g disodium ethylenediaminetetraacetic acid) for 4 min at room temperature; lysis was arrested with excess ice-cold medium followed by 2 washes. Hematopoietic chimerism was determined by GFP fluorescence or mAb against minor antigens CD45.1 (clone A20; eBioscience) and CD45.2 (clone 104; eBioscience). In all measurements non-specific binding was prevented by addition of 1 μL mouse serum.
Tissue preparation
Mice were sacrificed by CO2 asphyxiation., and exsanguination was performed by infusion of 30 mL of ice-cold PBS containing 1.5% fresh paraformaldehyde and 0.1% glutaraldehyde through a catheter inserted in the left ventricle. Excised pancreata were placed in this medium for 2 h at 4°C for additional fixation, and immersed in 30% sucrose overnight [32]. Tissue embedded in OCT (Sakura Finetek) was frozen in isopentane suspended in liquid nitrogen and sectioned (3–6 μm) with a Cryotome (Termo Shandon). Cryosections were stored at −80°C before analysis.
Determination of blood insulin
Serum from NOD mice was collected by centrifugation and assessed in 96-well Microtiter Assay Plates (Millipore) using the Rat/Mouse Insulin ELISA Kit (R&D Systems). Absorbance at 450 and 590 nm was determined using an ELISA PowerWave-10 in a plate reader (BioTeK). Insulin standards were used to determine a calibration curve.
Immunofluorescence and fluorescence in situ hybridization
Immunofluorescence was designed according to several considerations to allow detection of the donor cells with variable GFP fluorescence intensities [40]. Donor cell analysis in host pancreata was performed in 2 steps to include immunofluorescence in stained frozen sections and fluorescence in situ hybridization (FISH) [32]. Sections were fixed in acetone and permeabilized by incubation with 0.2% Saponin, 1% bovine serum albumin, and 0.1% Triton-100; were stained with primary antibodies for 1 h; washed; and counterstained with respective secondary antibodies for 30 min at room temperature. Nuclei were labeled with Hoechst-33342 (1:1,000; Molecular Probes), and sections were mounted in anti-fade medium (Dako) and air-dried. Serial cryosections from fixed pancreas were immunostained with primary antibodies: polyclonal mouse-anti-proinsulin (1:20; R&D Systems), goat-anti-PDX-1 (1:5,000, kindly provided by Shimon Efrat), rat anti-Pecam-1 (CD31, 1:100, CBL 1337, Chemicon-Millipore), biotinylated anti-mouse CD45 (1:100; Biolegend), biotinylated donkey-anti-mouse (1:1,000), and rabbit-anti-GFP (1:100; Santa Cruz Biotechnology). These primary antibodies were counterstained with fluorescein isothyocyanate-labeled donkey-anti-rabbit (1:200, Jackson Immunoresearch) and goat-anti-rat mAb (1:200; Santa Cruz), Alexa Flour 568-conjugated donkey-anti-goat (1:500), and Cy3-conjugated rat-anti-goat (1:200; Molecular Probes). Biotinylated primary antibodies were counterstained with Cy3-conjugated Streptavidin (1:400) and Cy5-conjugated Streptavidin (1:500) both from Jackson Immunoresearch.
X and Y chromosomes were painted in fresh sections or immunostained slides after treatment with 0.025% pepsin. Slides were rinsed with distilled water and 2 × saline sodium citrate (buffer) (SSC), dehydrated by sequential immersion in 70%, 85%, and 100% ethanol at room temperature and air-dried. A second fixation in warm 70% formamide was followed by repeated dehydration in gradually increasing ethanol concentrations. Nuclear probes (Applied Spectral Imaging) denatured at 74°C (7 min) and 37°C (30 min) were hybridized at 37°C overnight, followed by sequential washing in 0.4 × SSC stringency solution at 74°C and 2 × SSC solution supplemented with 0.1% NP4O detergent at room temperature. Washed slides were mounted with anti-fade containing 4′,6-diamidino-2-phenylindole. In our hands, Y chromosome was detected using a painting probe in 77%±14% of islet cells in male controls (approximately 400 islet cells).
Images were acquired with an Axioplan 2 (C. Zeiss) fluorescence microscope equipped with an Apotome and with a C. Zeiss confocal laser scanning microscope (LSM) using AxioVision 4.5 and LSM 510 software, respectively. Images were pseudocolored and RGB reconstructed using Adobe Photoshop software.
Statistical analysis
Data are presented as means±standard deviations for each experimental protocol. Results in each experimental group were evaluated for reproducibility by linear regression of duplicate measurements. Differences between the experimental protocols were estimated with a post hoc Scheffe t-test and significance was considered at P<0.05.
Results
The experimental model
A series of optimization experiments was performed to determine the approach (Fig. 1A): (a) Five doses of 60 μg/g STZ administered on consecutive days results in sustained hyperglycemia (400–500 mg/dL), and survival of C57BL/6 mice without exogenous insulin supplementation. (b) Since it is unknown whether the grafted cells home first to the bone marrow or the injured organ [41], the diabetic mice were sublethally irradiated at a dose of 675 rad. This combined injury causes severe damage to islets (Fig. 1B) and significant 43% (16 of 37) mortality; however, the hyperglycemic mice survived without exogenous insulin administration. (c) Analysis of the optimal temporal experimental sequence showed that infusion of the cells early after injury yields the best glycemic control. (d) The smallest subset of BMCs was collected by counterflow centrifugal elutriation (Fr25) and depleted of cells expressing lineage markers (Fr25lin−), resulting in a population largely positive for CD45 and negative for GR-1 [40]. Mice with chemical diabetes infused with Fr25lin− subset, which are not radioprotective [40], survived primarily through early recovery of endogenous hematopoiesis after sublethal irradiation.
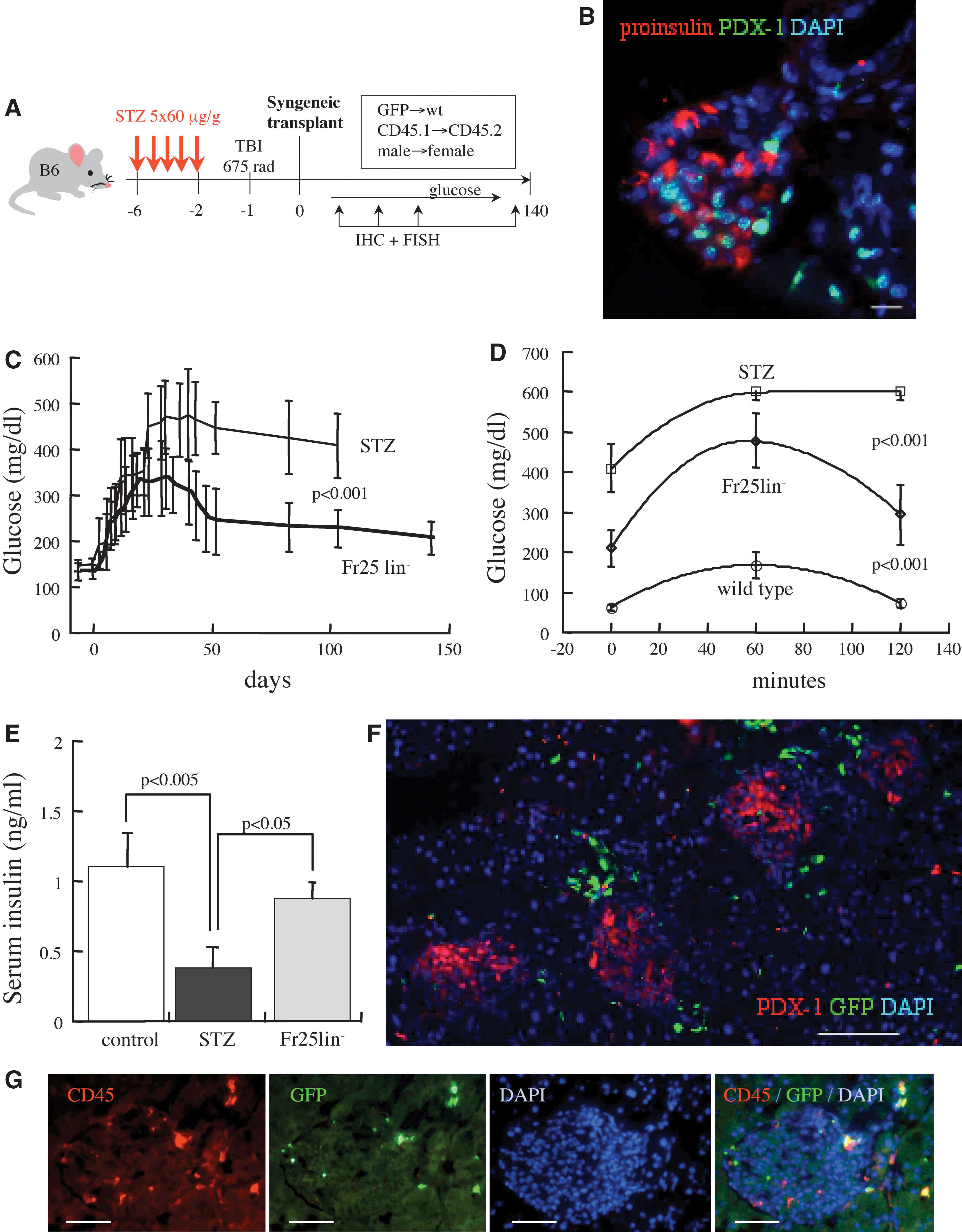
The experimental model and cell isolation.
Fr25lin− cells home to the islets and ameliorate the course of chemical diabetes
Transplantation of 106 elutriated Fr25lin− cells into syngeneic irradiated diabetic mice had a marked impact on the course of chemical diabetes. The cell transplant reduced peak blood glucose levels (P<0.005) and caused significant control of hyperglycemia after approximately 5 weeks (Fig. 1C). The markedly improved glycemic control over several months (P<0.001 vs. STZ at 15 weeks), coinciding with the capacity to respond to glucose tolerance test (Fig. 1D) and elevated insulin levels (Fig. 1E) at the experimental end point. The time to partial reversal of hyperglycemia was longer in our experiments than reported for models that differed in the experimental sequence, STZ doses, and types of grafted cells [10,11]; however, glycemic control was markedly improved than previously reported. At 7 months post-transplant significant numbers of donor cells were observed in the pancreas; however, disruption of islet architecture and scattered expression of PDX-1 and proinsulin emphasize persistent injury over extended periods (Fig. 1F).
To assess the patterns of cell migration to and incorporation in the injured pancreas, male Fr25lin−CD45.2+GFP+ donor cells were transplanted into female CD45.1 diabetic recipients. The grafted Fr25lin−GFP+ cells migrated efficiently to the injured islets, and GFPbright cells seeded preferentially in the vascular pedicle, ducts, and islet perimeter (Fig. 1G). Approximately 50% of the islet-homed cells expressed CD45, possible mediators of immunomodulation [10,14,20,25], or other supportive mechanisms of islet recovery from injury [11,13,17,24]. However, a significant fraction of Fr25lin− cells that migrated to the inner regions of the islets were CD45-negative, suggesting participation in non-immunogenic mechanisms. One mechanism mediated by CD45− cells is the contribution of Fr25lin−GFP+ cells to islet revascularization, affirmed by incorporation of GFP+CD31+ cells in the vasculature [32] as reported for other cell subsets [11,15,20]. Notably, endothelial differentiation occurred at 1–4 weeks post-transplantation, when Fr25lin− had a minor hematopoietic contribution, preceding in time the substantial improvement in glucose homeostasis.
Fr25lin− cells produce insulin
Delayed and gradual recovery of glucose homeostasis implies that the partial resolution of hyperglycemia was caused by differentiation of the grafted cells within the process of islet remodeling. To test this hypothesis we performed detailed analysis of phenotypic markers of β-cells: pancreatic and duodenal homeobox gene-1 (PDX-1) as an essential transcription factor involved in development of the endocrine pancreas [42], and proinsulin to prevent eventual uptake of insulin by the grafted cells from the environment [32]. This analysis revealed variable levels of GFP, PDX-1, and proinsulin in islet-incorporated Fr25lin− cells from GFP donors (Fig. 2A), which might originate from inefficient detection of the reporter protein [32]. Juxtaposition of cytosolic GFPdim with nuclear PDX-1 in islet-resident cells was confirmed by confocal microscopy (Fig. 2B), whereas the transcription factor was not detected in GFPbright cells surrounding the islets. Abundant proinsulin expression in these cells questions whether they are functionally deficient and unresponsive to systemic hyperglycemia, resulting in accumulation of the hormone precursor. Intense staining for insulin in donor cells (Fig. 2C) demonstrates their capacity to process the hormone precursor and make a true contribution to glycemic control.
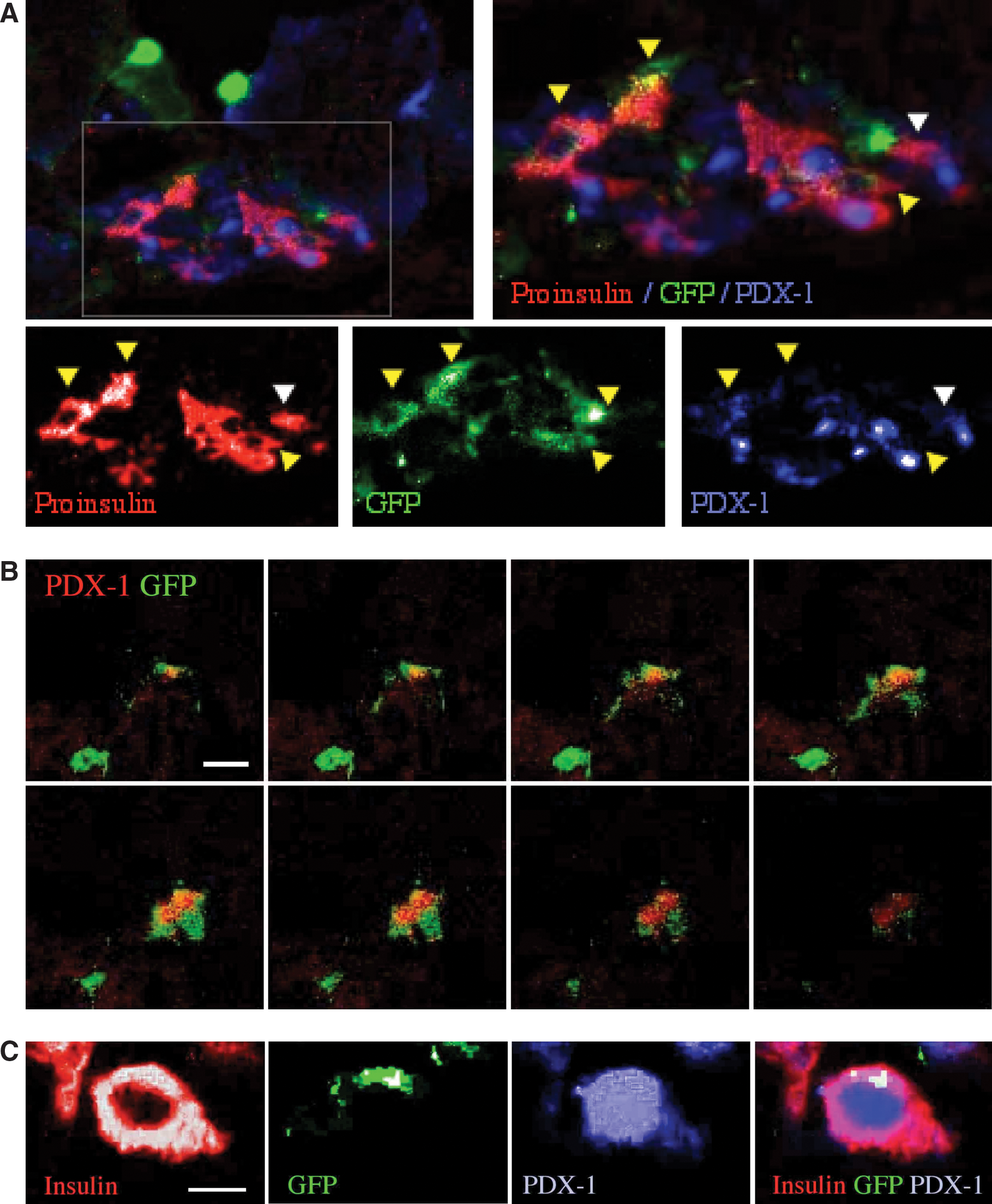
Islet-resident GFPdim donor cells produce insulin.
Quantitative evaluation of donor cell incorporation
Since downregulation of GFP might be characteristic of the process of differentiation of the Fr25lin− cells toward adopting a phenotype compatible with insulin production [32], further identification of donor cells was performed using a genomic marker in sex-mismatched transplants. Female recipients of male Fr25lin− cells displayed significant incorporation of Y chromosome among islet cells staining positive for PDX-1 (Fig. 3A). High-resolution analysis of 48 islets revealed the presence of Y chromosome and/or GFP donor cells in 38 islets, with significant variability in small islets as compared with larger islets (Fig. 3B). An overall 209 donor cells positive for PDX-1 and/or proinsulin among 2,750 evaluated β-cells indicate a 7.6% contribution to islet cellularity (Fig. 3C). In view of the challenges imposed by such analysis [32,43], this figure is a likely underestimate of differentiation of Fr25lin− cells.

Identification of donor cells by Y chromosome and quantitative analysis.
Islet-resident Fr25lin− cells do not display hematopoietic markers
Rigorous assessment of cell differentiation often imposes the demonstration that stem cells or their myeloid progeny are not fusion partners of the injured β cells, causing lateral transfer of specific markers rather than stem cells adopting an endocrine pancreatic differentiation trait [44 –47]. The majority of grafted cells expressed CD45 (84%±11%); however, approximately 50% of islet-incorporated cells were negative for this pan-hematopoietic marker (Fig. 1F). Analysis of sequential sections of the injured islets demonstrates CD45 and GR-1 expression in GFP+ cells located at the islet perimeter, but not in cells located within the inner regions of the islets (Fig. 4A), consistent with non-hematopoietic phenotypes of endothelial and insulin-producing cells. Fusion is expected to result in a donor Y chromosome co-residing with several donor and host X chromosomes, whereas donor cells differentiating to express PDX-1 in the inner islet regions retained normal diploid XY genotype (Fig. 4B). A positive control for these experiments is formation of syncitia composed of several GFP+CD45+ (and GR-1+) cells in proximity to the islets (Fig. 4C). The scattered appearance of proinsulin in these syncitial clusters most likely reflects clearance of tissue debris by immune cells at the islet perimeter. Although rare events of residual tissue fusion with donor stem cells and their myeloid progeny may have occurred [48], we could not demonstrate such phenomena that might result in lateral transfer of β-cell specific markers.

Insulin-producing donor cells lack hematopoietic markers and are not products of fusion.
Discussion
This study presents evidence that the smallest nucleated BMCs contribute to insulin production and ameliorate glycemic control over an extended period of time in a model of murine chemical diabetes. Grafted cells identified by a genomic marker adopted phenotypic characteristic of β-cells, produced insulin, downregulated GFP expression and were found negative for determinants of hematopoietic lineages.
Previous reports have underlined participation of bone marrow-derived cells in remodulation of the islets after injury. As descendents of a common progenitor, hematopoietic and endothelial precursors residing in the bone marrow support tissue revascularization and contribute to neovascularization [10,11,14,15,20]. In addition, local immunomodulation achieved by mesenchymal stromal cells [19,25], one [14] or multiple BMC infusions [17], or concurrent bone marrow mobilization and induction of hematopoietic differentiation [10,20], support tissue remodeling after injury through β-cell expansion [11,49] and/or activation of tissue progenitors [2 –8]. Mesenchymal stromal and immuno-hematopoietic cells support islet recovery also by remodulation of the stroma and provision of growth factors [18,1,22]. Direct contribution to neovascularization by incorporation of GFP+CD31+ cells in islet vasculature is evidence of developmental versatility of the Fr25lin− cells used in this study. However, the various functions attributed to different cell types derived from hematopoietic compartments were largely inefficient in reinstitution of glucose homeostasis over extended periods [9 –25].
Modest contributions of various hematopoietic cells to islet cellularity have been reported, ranging from incidental incorporation [10,19,22,23,26,50,51] to 3% after myeloablative bone marrow transplantation [12] and 2.5% GFP+insulin+ cells negative for PDX-1 after chemical injury [11]. In contrast, Fr25lin− cells migrated efficiently to the injured pancreas, with particular affinity to the islets, ducts, and vascular walls, and improved significantly glycemic control: CD45-negative cells that infiltrated the islets upregulated PDX-1 and produced proinsulin. Detection of proinsulin was selected to prevent an experimental bias caused by uptake of insulin from the injured β-cells [32]. Accumulation of the hormone precursor in the differentiated donor cells might indicate inefficient conversion to insulin, or underdeveloped glucose sensing machinery [52,53]. The proven capacity of donor cells to convert proinsulin into insulin, and elevated serum insulin levels in mice that recovered normoglycemia demonstrate neogenesis of competent cells that produce and secrete the hormone. Reconstitution of 7.6% functional insulin-producing cells originating from the male donor Fr25lin− stem cell-enriched fraction is a substantial mechanism of stabilization of glucose homeostasis. Participation of donor cells in islet regeneration is most likely underestimated, considering the technical difficulties in combined IHC and FISH analysis, the physiological downregulation of GFP expression in differentiating progenitors, and the anticipated silencing in endocrine cells within the islets [32,54]. Nevertheless, neogenesis of endothelium and insulin-producing cells affirms the developmental plasticity of this subset of adult bone marrow-derived stem cells in addition to conversion into hepatocytes and various epithelia [37 –39].
The time frame for differentiation into insulin-producing cells reported here is longer than previously observed in the liver [38] and other epithelial tissues [39], and is shorter than the contribution of these cells to long-term hematopoietic reconstitution [34 –36,40]. It is conceivable that restriction of immuno-hematopoietic differentiation is strongest in multipotent stem cells residing in the marrow space. Accordingly, effective hematopoiesis from Fr25lin− cells occurs last among the various non-hematopoietic differentiation traits adopted by these adult bone marrow-derived stem cells [38,39]. The bone marrow is a likely reservoir of repair units because they can be readily distributed to sites of injury via the systemic circulation. Fr25lin− cells are not only physiological residents of the bone marrow, but are endowed with particular affinity to this compartment, and can be enriched using an in vivo homing assay [41]. Further, these cells display several significant characteristics of primitive (stem) cells: quiescent mitotic state early after transplantation [36,40], self-renewal capacity emphasized by indefinite multilineage hematopoietic reconstitution [34 –36,40], and functional contribution to multiple tissue, including endothelium, endocrine pancreas, liver [37], and epithelia [39]. In addition, these cells display glial and neuronal markers when implanted in models of retinal injury in the absence of radiation, questioning whether these cells require initial homing to and priming in the bone marrow (N. Goldenberg-Cohen, unpublished data). We do not exclude the possibility that the Fr25lin− and Fr25lin−PKHbright subsets include the CD45−SCA-1+CXCR4+lin− very small embryonic-like (VSEL) cells [51], which is consistent with detection of CD45− donor cells within the injured islets in this and prior studies [19,26,50]. However, unlike VSEL, most of the grafted cells are CD45-positive, and depletion of SCA-1+ subsets is unrelated to functional islet reconstitution by Fr25lin− cells (S. Iskovich, unpublished data). Furthermore, molecular analysis of the elutriated Fr25lin− subset shows low level expression of mRNA and protein of determinants of the endocrine pancreatic differentiation trait (N. Goldenberg-Cohen, unpublished data), negating the possibility of the presence of committed tissue progenitors within the fraction of BMCs used in this study [55,56].
Despite the rightful criticism raised by negative results [9,32,57,58], we report failure to detect fusion events. Both stem cells and their myeloid progeny have been shown to adopt parenchymal phenotypes through lateral transfer of markers caused by cell fusion [44 –47]. We could demonstrate neither pluriploid X chromosomes in the PDX-1+proinsulin+ donor cells consistent with stem-parenchyma chimerism, nor expression of CD45 or GR-1 consistent with uptake of β-cell markers through fusion or phagocytosis. The possibility that fusion can reconstitute tissue function remains to be determined in view of the very low frequency of these events [48]. According to the incidence reported in a fusogenic organ such as the liver [44 –47], it is estimated that fusion would account for 10−2–10−3 of the 7.6% donor cells producing insulin, and therefore is a quite insignificant mechanism.
The possibility to intervene in the process of islet remodeling and design regenerative approaches is a tangible perspective. Somatic pancreatic cells have been reverted to and endocrine phenotype by expression of few transcription factors [59] and interconversion of cell types within the islets has been achieved by regression to a more primitive state and redirection [60]. Our current data demonstrate robust neogenesis of insulin-producing cells from adult bone marrow-derived primitive cells in vivo, which can stabilize glycemic control. In addition to the capacity of nonmyeloablative bone marrow transplantation to arrest autoimmune insulitis [61], enriched autologous populations of primitive cells have the potential to regenerate the tissue.
Footnotes
Acknowledgments
We thank Dr. Saul Sharkis and Dr. Michael Collector for the outstanding support, discussion, and conceptual contribution to this study. Funding was provided by a generous grant from the Leah and Edward M. Frankel Trust for bone marrow transplantation.
Author Disclosure Statement
No competing financial interests exist.