
Abstract
Cell mobilization, a process that influences circulation, margination, and finally, homing play key roles in the regeneration processes mediated by stem cells. Recent studies as well as prior studies from our group indicate an important role of the spleen in hematopoietic reconstitution, but to date the role of the spleen in hematopoietic reconstitution has been unclear and it has not been precisely documented in ablated animals. Therefore, we undertook the present study to define more closely the role of the spleen in hematopoietic reconstitution in lethally irradiated mice. After transplantation of irradiated mice with lacZ+ -marked lin−/CD117+ bone marrow cells, we compared splenectomized mice (TS, splenectomy performed prior to irradiation) to nonsplenectomized, irradiated mice (TN) as well as to normal (unirradiated) mice. Impaired hematopoietic reconstitution was observed in TS mice. Splenectomy markedly altered the distribution of hematopoietic stem cells, as demonstrated by fluorescence-activated cell sorting analysis of endogenous CD117+ cells in the thymus and bone marrow of recipients. Cell engraftment was demonstrated by histochemical and polymerase chain reaction analyses of recipient tissues. These experiments demonstrated that in TS animals, transplanted hematopoietic stem cells mobilized to extravascular tissues, particularly the gastrointestinal tract. The number of donor cells in recipient tissues continued to increase for 30 days after transplantation with the highest numbers observed in the TS group. DNA marking analysis led to the conclusion that engrafted cells were not only integrated into recipient tissues but were also capable of performing complex cellular processes, including proliferation and repair. Our results are consistent with the novel possibility that cellular repair markedly affects stem cell regenerative functions and that repair is markedly influenced by the integrity and presence of organs not directly involved in specific tissue regeneration processes, particularly the spleen.
Introduction
H
During homeostasis, homing constitutes a major mechanism that maintains the integrity of all organs and tissues. Various cells show different homing profiles, which can be objectively measured [8,9]. Several components of the essential process of stem cell homing have now been characterized. Cell adhesion molecules and their ligands, extracellular matrix components, chemokines, and specialized bone marrow niches all participate in the precise regulation of this process [4,9,10].
Kawaguchi (2005) utilized B220+ precursors with a short-term reconstitution capacity from CD117+/B220− leukocyte subset, which contains hemopoietic stem cells (HSCs). Experimental transplantation of B220+CD117+ bone marrow cells in lethally irradiated mice led to B220+ B-lymphocyte renewal within 8 days after transplantation. The increase in B-cell number ceased by day 10, but cell numbers remained at the same level for 8 subsequent weeks. B-cell production in the recipient bone marrow occurred 8 weeks post-transplantation; reconstitution following transplantation of these cells was faster than after CD117high CD71− cell transplantation [11].
In our prior studies, we evaluated hematopoietic repair under several conditions. We observed a statistically significant difference in granulocyte-macrophage colony-forming units (CFU-GM) numbers in both spleen and bone marrow within day 8 after transplantation of CD117+B220− (lacZ +) cells. Hematopoietic repair assessed by peripheral cell numbers started on day 12 after transplantation [12], demonstrating a slower hematopoietic repair after transplantation of separated cells [13,14].
Several questions thus emerge. One unanswered question is whether a lower hematopoiesis repair affects engraftment of transplanted cells. Another seeks definition of the role of endogenous cells and their microenvironment in stem cell regeneration. Endogenous hematopoietic cells represent a natural reserve participating in hematopoiesis reconstitution [15,16]; organ-specific c-kit+ lymphoid precursors are present in various immune organs including liver, thymus, and bone marrow [17].
In our experimental model, whole-body irradiation of mice by lethal doses was used for elimination of endogenous hematopoiesis and demonstration of the effect of HSCs transplantation [18,19]. Our previous results and those of others opened the question of how the spleen influences cellular circulation and in vivo homing and, consequently, participates in the recovery of hematopoiesis [20]. Therefore, the goal of this study was to compare different effects of whole-body irradiation and splenectomy on the circulation and homing of transplanted bone marrow HSCs, selected by their expression of CD117+ and the absence of expression of lineage antigens. We examined how splenectomy influences circulation and homing of transplanted cells to the bone marrow as well as to sites of extramedullary hematopoiesis. The results show that intravenously infused HSCs do not necessarily engraft in the bone marrow. Splenectomy markedly alters cellular repair processes by its influence on the distribution and homing of engrafted stem cells. Similar alterations may result from ablation of other organs. Our results indicate that cell-mediated repair can be markedly influenced by the integrity and presence of organs not directly involved in the repair process, such as the spleen.
Materials and Methods
Mice
B6;129S-GT(ROSA)26Sor mice (n=6) were obtained from the Jackson Laboratories and their cells were used as donor cells for transplantation, because they are tagged with lacZ gene and express Escherichia coli β-galactosidase (β-gal) to allow unambiguous identification of transplanted cells. F2 hybrid mice B6129SF2/J (n=6; in each group) were used as recipients of transplanted bone marrow cells. Control group consisted of F2 hybrid mice B6129SF2/J (n=6) without irradiation and splenectomy.
Splenectomy
Splenectomy was performed under general anesthesia. After laparotomy, the spleen was removed and the wound in the abdominal wall was closed with stainless-steel clips, which were left in place. Mice were allowed to recover for at least 14 days before they were subjected to different protocols.
Isolation of lin−/CD117+cells from the bone marrow
Bone marrow cells were obtained from femur of 8–10-week-old donor mice and suspended in Dulbecco's phosphate-buffered saline (Sigma) supplemented with 2% fetal calf serum. Bone marrow cells (2×107) were subjected to negative immunomagnetic selection with a Lineage Cell Depletion Kit (Miltenyi Biotec) for removal of differentiated cells. These cells were then subjected to positive immunomagnetic sorting with CD117 MicroBeads (Miltenyi Biotec). Lin−/CD117+cells from bone marrow were labeled with CD117 (c-kit)-PE (Miltenyi Biotec) and anti-lineage-biotin/anti-biotin-APC for determination of purity of isolated cells.
Irradiation and reconstitution
Recipient animals (F2 hybrid mice) were exposed to 9 Gy irradiation from a 60Cobalt source (Chisotron, Chirana) at a dose rate of 1.3 Gy/min. Suspension of 5×104 lin−/CD117+ (lacZ+ ) bone marrow cells were transplanted by intravenous injection into recipient animals at 3 hours after irradiation.
Clonogenic assays for myeloid progenitors
Cells from the bone marrow and spleen were harvested on days 8, 12, and 30 following mice transplantation. CFU-GM were assayed after plating cells in MethoCult GF M3434 with cytokines (StemCell Technologies). Aggregates with more than 40 cells were considered to be colonies and were scored with light microscopy at 12–14 days after incubation at 37°C in a humidified atmosphere of 5% CO2 in air.
Flow cytometric detection of CD117+ cells
Cells from whole heparinized peripheral blood and single-cell suspensions prepared from the entire thymus and bone marrow from mice (n=6) exposed to 9 Gy irradiation and from nonirradiated controls (n=6) were analyzed using the flow cytometer CyAN-ADP (DakoCytomation) on days 8, 12, and 30 post-transplantation. Cells were double-stained with rat anti-mouse CD45R/B220-FITC antibody (clone RA3-6B2; SouthernBiotech) and CD117/c-Kit-PE (clone 2B8; SouthernBiotech).
Histology
Recipients were sacrificed under deep anesthesia on the 8th, 12th, and 30th day following transplantation and their tissues (bone marrow, thymus, liver, spleen, stomach, and small intestine) were carefully removed and fixed in 4% paraformaldehyde in phosphate buffer (pH 7.8) for 15 min. Identification of transplanted cells was performed with X-gal solution (pH 7.8) and then the samples were dehydrated and embedded in paraffin. For immunohistochemistry, deparaffinized sections containing X-Gal+ cells were treated with microwaves for antigen retrieval (3×10 min, citrate buffer, pH=6.0), then with 3% hydrogen peroxide to inhibit pseudoperoxidase activity, and thereafter with unconjugated anti-mouse Fab fragments to prevent a nonspecific binding of secondary antibody. A proliferative marker was detected with anti-PCNA antibody (clone 5A10; Immunotech), which was followed by incubation with donkey anti-mouse biotinylated antibody. After incubation with streptavidin coupled with horse radish peroxidase, the detection was visualized with 3,3′-diaminobenzidine. Tissue sections were counterstained with hematoxylin and examined in Olympus BX-51 microscope equipped with a digital DP-71 camera.
Quantitative real-time polymerase chain reaction detection of lacZ gene expression
The quantitative real-time polymerase chain reaction (qRT-PCR) detection of lacZ gene expression was analyzed by ABI 7500 Fast Real-Time PCR System (Applied Biosystems). Total RNA was isolated from samples using Qiagen RNeasy® Kit (Qiagen). These were converted into cDNA via Superscript™ III First-Strand Synthesis System (Invitrogen). cDNA (30 ng) was then analyzed in triplicates. The mRNA sequence for the target lacZ gene was obtained from NCBI database (number of sequence: NC 011750.1; Gene ID: 7151913) and the primers and probe (Generi Biotech) were subsequently designed as previously described with the following sequences (5′–3′): forward primer GCG TTA ACT CGG CGT TTC AT, reverse primer GCG CTC AGG AAT TCA GAC, and probe CGG TTA CGG CCA GGA CAG TCG TTT G. The amplification was completed using TaqMan® Fast Universal PCR Master Mix (Applied Biosystems). As a housekeeping gene, Eukaryot 18S rRNA endogenous control kit (P/N4319413E) provided by Applied Biosystems was used. The time–temperature profile used was 50°C for 2 min, 95°C for 10 min followed by 40 times of 95°C for 15 min, and 60°C for 1 min. The relative expression ratio (ΔΔCt) was then calculated using ΔCttarget and ΔCthousekeeping of evaluated and reference (tissue from standard F2 mice) samples.
Statistical analysis
Data were expressed as the mean±standard deviation of the mean to analyze hematopoietic recovery, flow cytometry, and qRT-PCR. Statistical significance was determined using the Student's t-test. The differences were assumed significant at P≤0.001.
Results
Hematopoietic recovery after transplantation
Analysis of hematopoietic recovery was performed in lethally irradiated nonsplenectomized (TN) and splenectomized (TS) mice by days 8, 12, and 30 following transplantation of lin−/CD117+ (lacZ +) separated from the bone marrow. Control group (n=6) consisted of normal mice without lethal irradiation or splenectomy. Evaluation of hematopoietic recovery after transplantation was based on peripheral blood count changes (lymphocyte, grynulocyte, monocyte, erythrocyte, and thrombocyte counts) and changes of CFU-GM numbers in the bone marrow.
When TN and TS groups were compared with the control group, a significant drop in the numbers of lymphocytes, granulocytes, and thrombocytes was observed in the peripheral blood on day 8 after cell transplantation. When TN and TS groups were compared, a significant difference was also noted in thrombocyte count as a significant drop occurred in the TS group. Similarly, a drop of monocyte count was also found in the TS group. Contrary to this, no significant changes in erythrocyte counts were observed in both the TN and TS groups (Table 1).
Statistically significant differences in the hematopoietic recovery shown as the peripheral blood cell count and a total number of CFU-GM colonies in a control group (n=6) and transplanted mice (by days 8, 12, and 30 post-transplantation) were indicated in ATN (n=6), BTS (n=6), and Cdifferences of TN versus group TS. The data are reported as arithmetic mean±SD; CFU-GM colonies/105 MNCs. Statistical significance was determined using Student's t-test. The differences were assumed significant at P≤0.001.
CFU-GM, granulocyte-macrophage colony-forming units; MNCs, mononuclear cells; TS, splenectomized mice; TN, nonsplenectomized mice; SD, standard deviation of the mean.
On day 12 after transplantation, a rise in lymphocyte, granulocyte, and thrombocyte counts in peripheral blood occurred. This rise, however, did not reach normal values measured in the control group. In the TS group the significant drop in monocyte count persisted. Changes in erythrocyte counts were not statistically significant (Table 1).
On day 30 after transplantation, the rise of lymphocyte count to normal values was observed only in the TS group. In the TN group, normal values were not reached when compared with controls. Granulocyte and monocyte counts were normalized in both TN and TS groups. Erythrocyte counts were not significantly changed. Contrary to this, a significant rise of thrombocyte count was observed in the TS group when compared with both the control group and the TN group (Table 1).
On day 8 after transplantation, a significant rise of CFU-GM was found in the bone marrow in the TS group when compared with both the control and TN groups. At day 12 and 30, normalization of CFU-CM count was observed only in the TN group. Contrary to this, by days 12 and 30, a further drop of CFU-GM count in the TS group was observed and this was significant when compared with controls and the TN group (Table 1).
The presented results show that splenectomy leads to thrombocytosis in the peripheral blood because of elimination of spleen. Further, splenectomy probably has a significant stimulating influence on production of progenitor cells (CFU-GM) in the bone marrow early by day 8 after cell transplantation. On the contrary, a suppressive effect of splenectomy significantly reducing CFU-GM count was found in the bone marrow of the TS group on day 30 (Table 1).
Fluorescence-activated cell sorting analysis of bone marrow and thymus
Flow cytometric examination of central hematopoietic organs, thymus (Fig. 1A) and bone marrow (Fig. 1B), was performed in the TN and TS recipients to assess the presence of CD117+ (lacZ −) cells. Analysis was carried out in the TN and TS groups on the 8th, 12th, and 30th day after transplantation of lin−/CD117+ (lacZ +) bone marrow cells. Analysis in the control group (n=6) was performed in normal mice without lethal irradiation and splenecomy.
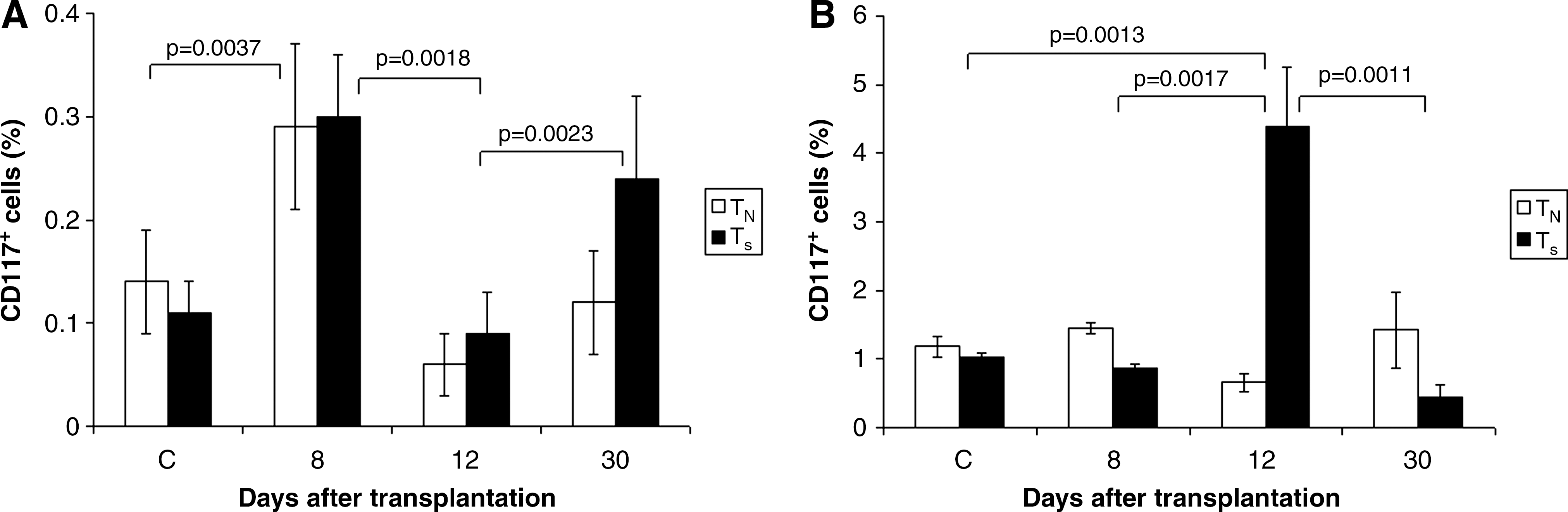
Fluorescence-activated cell sorting analysis of thymus and bone marrow of transplanted animals. Analysis of CD117+ (lacZ
−) cells in thymus
On day 8 post–transplantation, the significant rise in CD117+ cells in thymus was noted in both the TN and TS groups when compared with controls. However, another drop was recorded by day 12 but by day 30 the rise occurred again. This was significant but did not reach the values reached by day 8 post-transplantation. Moreover, this rise was observed only in the TS group (Fig. 1A). In the bone marrow, the significant rise in CD117+ cells was seen 12 days post-transplantation in the TS group when compared with the control and TN groups. (Fig. 1B). Although splenectomy induced a significant rise in CD117+ cells in the thymus as early as 8 days after transplantation, there was no significant difference when the control group was compared with the nonsplenectomized group (TN). It was also observed that splenectomy influenced a significant rise in CD117+ cells in thymus up to day 30 post-transplantation. Splenectomy significantly influenced the count of CD117+ cells in the bone marrow as early as by 12 days after transplantation. During interval between days 8 and 30 after cell transplantation, no significant changes were seen when compared with controls and the TN group. It can be concluded that splenectomy influences the number of CD117+ cells in thymus in the first place, and in the bone marrow this influence gradually ceases and lasts for only a short time (Fig. 1A, B).
Histochemical analysis in tissues after transplantation
To verify presence of transplanted cells and to examine their distribution in recipient tissues, tissue samples obtained from TN (Fig. 2A) and TS (Fig. 2B) mice were incubated with X-Gal substrate to identify cells with enzymatic β-gal activity that is characteristic of donor lacZ + cells. Microscopic examination of histological sections confirmed a gradual increase in tissue colonization by transplanted cells in time. By day 8 after transplantation, recipient tissues contained only sporadic positive cells. By day 12, the findings were more pronounced. A relatively large number of diffusely scattered X-Gal+ cells colonized the thymic cortex. Positive hematopoietic cells were visible in the splenic and hepatic parenchyma. In organs of gastrointestinal tract, grafted cells colonized the lamina propria mucosae and rarely the tela submucosa; these cells occasionally aggregated in lymphatic nodules. We observed positive nodules not only in the small intestine but rarely also in the colon, stomach, and esophagus. Thirty days after transplantation, a number of positive cells still increased—more X-Gal+ cells occupied tissues in TS animals.
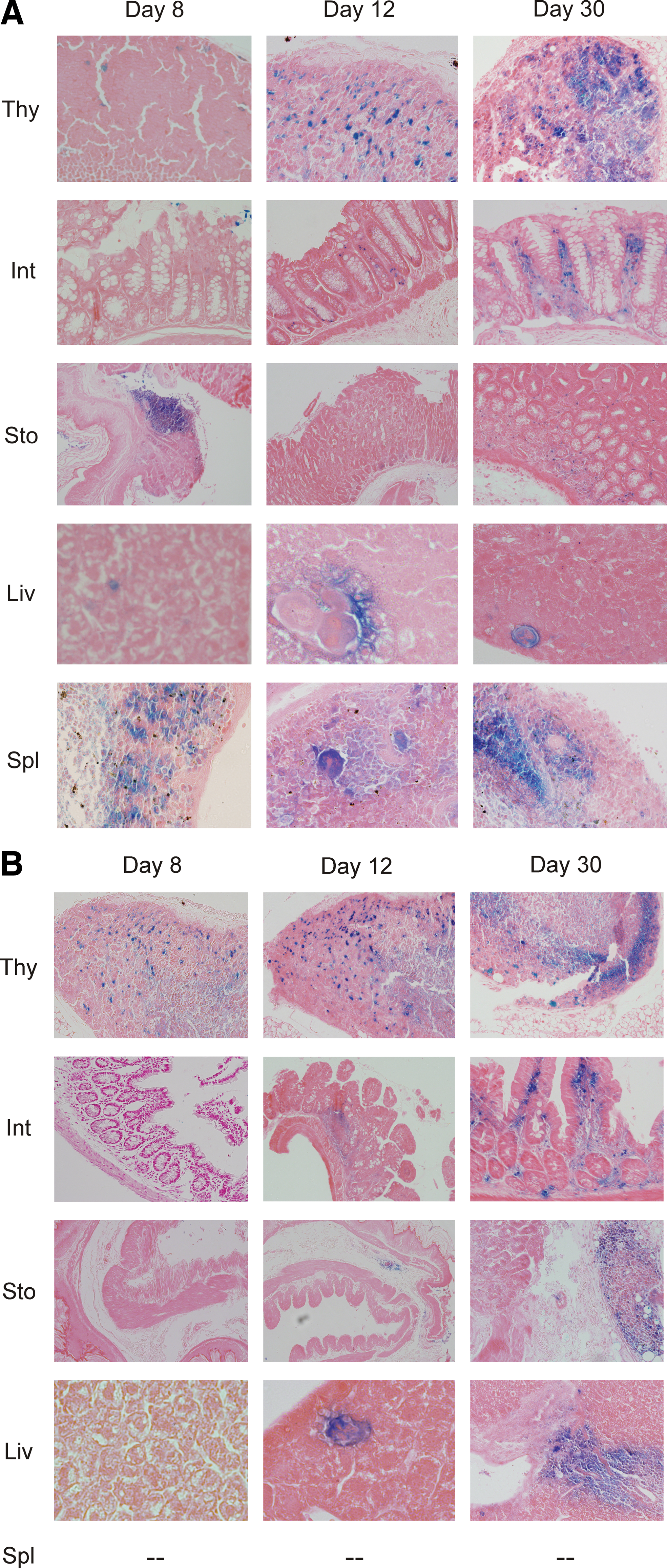
Histochemical analysis of cell engraftment. For unambiguous identification of transplanted ROSA26 cells, the thymus (Thy), small intestines (Int), stomach (Sto), liver (Liv), and spleen (Spl) isolated from TN mice on days 8, 12, and 30 were processed for X-Gal histochemistry
We analyzed proliferating cell nuclear antigen (PCNA) immunoreactivity of transplanted cells to find out whether they undergo mitotic divisions following their engraftment. The thymus and spleen sections contained too large amounts of overlapping PCNA+ and lacZ+ cells that rendered microscopic analysis difficult. On the other hand, the liver contained only few transplanted cells. Moreover, the small intestine was the most suitable organ for analysis, because its sections contained sufficient amount of lacZ+ cells that did not fuse in large areas and, as a result, both positive and negative cells could be observed side by side. PCNA immunoreactivity showed a characteristic distribution in the intestinal covering epithelium: epithelium of Lieberkühn crypts was immunopositive, whereas epithelium in intestinal villi remained negative. However, intestinal covering epithelium did not show any β-gal activity; X-Gal+ cells were located in the core of intestinal villi (Fig. 3A, B). Some cells also coexpressed PCNA or PCNA was expressed by some recipient endogenous cells. There was no significant difference in numbers of PCNA+ cells in TS versus TN mice.

Immunohistochemical detection of proliferating cell nuclear antigen (PCNA) expression in transplanted tissues. PCNA expression in the mucosa of small intestine of TN mice at 30 days after transplantation
qRT-PCR detection of lacZ expression in tissues after transplantation
To determine the participation of lin−/CD117+ in hematopoietic repair and their distribution in recipient tissues, we carried out qRT-PCR analysis of selected hematopoietic and nonhematopoietic tissues from both experimental groups with the aim to identify the lacZ gene expression that was used as an endogenous marker of transplanted cells. As PCR data are limited in terms of inability to distinguish cell migration and proliferation, all tissue samples were processed in parallel for histochemistry (see earlier).
By day 8 after transplantation, lacZ expression was detected in all tissue samples investigated (Fig. 4A, B). The highest value was reached in the spleen of TN mice, which highlights a crucial role of this organ in in vivo homing of transplanted hematopoietic cells. Relatively high levels of lacZ expression were also identified in the bone marrow and thymus of both transplanted groups. By day 12 post-transplantation, lacZ expression in tissues increased further, which indicated colonization of recipient organs with transplanted cells. Significantly increased numbers were observed, namely in the bone marrow (7×) and liver (15×) of TS animals. By day 30 following transplantation, lacZ mRNA reached significantly different values, especially in the spleen (7×), thymus (6×), and small intestine (nearly 5×) in TN mice. By that time, only minimal changes were observed in the liver and stomach. In general, lacZ expression in the bone marrow of TN mice reached higher levels than in TS mice, whereas in the thymus, small intestine, and liver, a higher degree of engraftment was encountered in TS animals (Fig. 4A, B). These results are consistent with engraftment of the cells in sites of extramedullary hematopoiesis. The spleen thus modifies circulation and homing of stem cells destined for engraftment.

qRT-PCR detection of lacZ expression in recipient tissues. Expression of lacZ on days 8, 12, and 30 after transplantation in TN
Discussion
Transplantation of hematopoietic stem cells and progenitor cells has a variable role in rescue of hematopoiesis. Migration of hematopoietic stem cells through the blood, across the endothelial lining of the vasculature to different organs, and to their bone marrow niches requires active navigation. Homing is a rapid, multistep, poorly understood process [21] that is essential for successful transplantation [22].
In our experiments, we used splenectomy and whole body lethal radiation to study tissue repair and contribution of homing of transplanted cells in these processes. The resulting reconstitution of hematopoiesis is associated with general repair processes that occur within experimental organisms. The issues linked to the regeneration processes and homing of transplanted cells have been intensively discussed [10,23]. Our study was aimed at comparison of different effects of whole body irradiation and splenectomy on circulation of transplanted lin−/CD117+ bone marrow cells. To trace distribution of transplanted cells in the recipient organism, we utilized donor cells tagged to endogenous vectors. We also focused on examination of the splenectomy role in cellular circulation and homing in the bone marrow and sites of extramedullary hematopoiesis and on cell compensation achieved by transplanted stem cells [20].
In these experiments, we confirmed the already known effect of splenectomy when thrombocytosis occurs in the peripheral blood [24]. This phenomenon is linked with circulation of other blood elements such as activated leukocytes and different developmental forms of megakaryocytes [25]. Circulation and homing of these cells may induce changes in the bone marrow, which lead to timely isolation and release of progenitor cell pool. This phenomenon was observed even in our experiments wherein, on day 8 after transplantation, a significant rise of CFU-GM count in the bone marrow could be seen in the TS group. This stimulating effect, however, did not last long, and on day 30 post-transplantation, again the significant drop in CFU-GM count in this group occurred. This finding documents once more the suppressive effect of splenectomy on the bone marrow [26].
Our results open the question how the splenectomy affects cellular circulation and homing and, consequently, participates in the recovery or regeneration process. To address this question, we performed fluorescence-activated cell sorting analysis of endogenous CD117+ (lacZ −) cells in the thymus and bone marrow. This experiment showed that splenectomy induces a significant increase in CD117+ counts as early as by day 8 post-transplantation. Splenectomy also significantly influenced the number of CD117+ cells in the bone marrow only 12 days after transplantation. Therefore, it can be concluded that splenectomy influences the counts of CD117+ cells mostly in the thymus, whereas in the bone marrow its influence is substantially decreased and lasts only a short time. These data indicate the links existing between the thymus and bone marrow that allow communication of organs participating in hematopoiesis repair [27,28].
A study, using population enriched of multipotent primitive hematopoietic progenitor cells of mice, confirms that cell homing occurs few hours after injection of GFP+ cells. Transplanted cells were detected not only in hematopoietic tissues but also in lungs, liver, muscles, colon, kidney, and brain [29]. Using histochemical staining, we detected transplanted cells in tissues of the gastrointestinal tract where donor cells settled in the lamina propria and rarely in submucosa. Occasionally, X-Gal+ cells aggregated in lymphatic nodules; positive nodules were observed not only in the small intestine but also in colon and stomach (Fig. 3A). Thirty days post-transplantation, the number of positive cells in the recipient tissues increased further and the TS group of TS animals contained more X-Gal+ cells (Fig. 3B) than TN mice. Recipient tissues were screened for PCNA-immunoreactive transplanted cells. An increase in the number of donor cells detected in the recipient tissues with the use of RT-PCR or histochemical analysis could be explained from proliferation of transplanted cells. Although the experiment described in this study did not involve labeling of newly synthesized DNA with thymidine analogs, we attempted to address this issue with immunohistochemical identification of endogenous proliferative markers in transplanted cells. The finding of coexpression of proliferative cell nuclear marker PCNA in nuclei of X-Gal+ cells confirmed that cells underwent mitotic division in host tissues even by day 30 after transplantation. Further, these findings gave evidence that transplanted cells were not only passively incorporated in recipient tissues but also were capable of performing complex activities such as mitotic division. Positive cells were not found among epithelial cells but in the stroma, for example, in the core of intestinal villi (Fig. 3A, B).
Our results demonstrate that splenectomy markedly alters cellular repair processes by its influence on the distribution and homing of engrafted stem cells and indicate that the cell-mediated repair can be markedly influenced by the integrity and presence of organs not involved directly in the repair process, such as the spleen. Thus, a tissue repair effected by stem cells is a complex, multifaceted process that is influenced by both soluble factors, cellular mobilization and organ integrity.
Footnotes
Acknowledgments
This work was supported by grant projects GACR 304/08/0329, MZO 00179906, and MSMT 0021620820 of the Czech Republic.
Author Disclosure Statement
No competing financial interests exist.