
Abstract
Our previous studies have suggested that transduction of Wnt11 directly increases bone marrow-derived mesenchymal stem cells (MSCs) differentiation into cardiac phenotypes. In this study, we investigated whether Wnt11 enhances MSC-mediated cardioprotection via paracrine fashion after acute ischemia. MSCs were harvested from male rat bone marrow and transduced with Wnt11 (MSCWnt11). An acute myocardial infarction model in rats was developed by ligation of the left anterior descending coronary artery. MSCWnt11 were transplanted into the peri-infarct region after acute myocardial infarction. To mimic ischemic injury, cultured cardiomyocytes (CMs) isolated from neonatal ventricles were exposed to hypoxia. ELISA studies indicated that the release of Wnt11 (3.45-fold) as well as transforming growth factor-β2 (TGFβ2) (1.5-fold) was significantly increased from MSCWnt11 compared with transduced control MSC (MSCNull). Hypoxia-induced apoptosis and cell death was significantly reduced when CM were co-cultured with MSCWnt11 in a dual chamber system. The cell protection mediated by MSCWnt11 was mimicked by treating CM with conditioned medium obtained from MSCWnt11 and abrogated by Wnt11- and TGFβ2 neutralizing antibodies. Further, animals receiving MSCWnt11 showed a significant improvement in cardiac contractile function as assessed by echocardiography. Masson trichrome and TUNEL staining showed a significant reduction in infarct size and apoptosis of CM in MSCWnt11-treated animals. Transplantation of MSCWnt11 improved cardiac function. The release of Wnt11 and other factors from transplanted MSCWnt11 is more likely responsible for protection of native CM at risk.
Introduction
S
Wnt proteins also play an important role in a variety of biological processes, including cell growth and proliferation, cell polarity and migration, apoptosis, and differentiation [15]. Wnt3a induces apoptosis via the canonical pathway [16] and chronic Wnt3 stimulation may contribute to stem cell depletion and aging in young Klotho mice [17]. In contrary, Wnt11 can reduce PC3 cells apoptosis [18] and increase cell survival in breast cancer cells [19]. Overexpression of Wnt11 enhances viability of Chinese hamster overy-K1 cells (CHO-K1 cells) and extends their grow periods [20]. Wnt11 has been reported to induce cardiomyogenic differentiation in stem cells, including unfractionated bone marrow mononuclear cells [21] and circulating progenitor cells [22], suggesting that Wnt11 may mediate myogenic differentiation. Recently, we have directly transduced Wnt11 into MSC and found that Wnt11 increases the potential of MSC transdifferentiation into cardiac phenotype [23].
Transplantation of MSC engineered Wnt11 into the border of ischemic myocardium may persistently release Wnt11 and other paracrine factors into ischemic myocardium. It is unclear whether Wnt11 can confer a protection on existing native CM in ischemic microenvironment when it is delivered by MSC. In this study, we investigated the cytoprotective ability of MSCWnt11 against ischemic injury via transplanting these cells into ischemic myocardium and co-culturing these cells with native CM. Our results indicate that Wnt11 as well as other factors released from MSCWnt11 may contribute to cardiac function recovery from acute ischemic injury and protect CM at risk.
Methods
All protocols were approved by the University of Cincinnati Animal Care and Use Committee, and conform to the Guidelines for the Care and Use of Laboratory Animals prepared by the National Academy of Sciences and published by the National Institutes of Health (NIH publication No. 85-23, revised 1996).
MSC transduction with Wnt11
MSCs were obtained from femurs and tibias of male SD rats, as described previously [24,25] with minor modifications. Cultured cells were transduced with recombinant Wnt11 using the murine stem cell virus (pMSCV) retroviral expression system, as described previously [23]. Briefly, Wnt11 was excised from pcDNA-Wnt11 with HindIII and XhoI restriction enzymes, and cloned into pMSCV-EGFP. GP2-293 cells (Clontech) were transduced with pMSCV-EGFP-Wnt11 using Fugene 6 transfection reagent. After 24 h, supernatant was filtered and incubated with MSC for 12 h in the presence of 10 μg/mL polybrene (Sigma). Stable transduced MSC clones were obtained by puromycin selection (Sigma; 3 μg/mL for 5 days). Negative control MSCs were either transduced with pMSCV-IRES-EGFP (MSCNull), or not transduced (MSCbas).
Preparation of conditioned medium
The conditioned medium (CdM) from cultured MSC was obtained as previously reported [26]. Briefly, MSCs were trypsinized and seeded in a 15-cm plate. After 24 h, regular culture medium was replaced with serum- and antibiotic-free Iscove’s Modified Dulbecco’s Medium (IMDM). Twenty-four hours later, medium was collected and centrifuged (2,500 rpm for 3 min) to remove cell debris. The medium was transferred to ultra-filtration conical tubes (Amicon Ultra-15 with membranes selective for <5 kDa), and centrifuged (4°C, 4,000g for 30 min) to produce CdM. The final concentration was adjusted with PBS to 100× of collected CdM. Wnt11 and TGFβ2 in concentrated CdM were measured using western blot and ELISA (R&D kit), respectively.
Microarray hybridization
To investigate the secretome of MSCWnt11, large-scale microarray analysis was performed at the Genomics and Microarray Laboratory of the University of Cincinnati Medical Center as described in published reports [27,28]. Total RNA from MSC was isolated using Trizol reagent (Invitrogen), followed by DNase treatment and purification using RNeasy mini kit (Qiagen) when culture dishes of MSC were approximately 80% confluent. Fluorescence-labeled cDNAs were synthesized from total RNA using an indirect amino allyl labeling method via an oligo(dT)-primed, reverse transcriptase reaction. The cDNA was labeled with monofunctional reactive Cyanine-3 and Cyanine-5 dyes (Cy3 and Cy5; Amersham). The 70-mer rat oligonucleotide Operon Library Version 3.0 (26,962 oligos; Operon Biotechnologies) was suspended in 3× SSC at 30 μM and printed at 22°C and 65% relative humidity on aminosilane-coated slides (Cel Associates) using a high-speed robotic Omnigrid machine (GeneMachines) with Stealth SMP3 pins (Telechem). Spot volumes were 0.5 nL and spot diameters are 75–85 μm. The oligonucleotides were crosslinked to the slide substrate by exposure to 600 mJ of ultraviolet light. Imaging and data generation were performed using GenePix 4000A and 4000B and associated software (Axon Instruments). To assess the effects of Wnt11 overexpression on gene expression in MSC, pair-wise comparisons were performed using 2 filtering strategies: comparison of transcripts expressed in 4 samples, and at least 1.5-fold change (P<0.05).
Immunoblotting of Wnt11
Wnt11 released from MSC into concentrated CdM was measured using immunoblotting. Protein concentrations were quantified with Bio-Rad DC-Protein Assay Reagent (Bio-Rad). Denatured proteins (50 μg) were separated using 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to nitrocellulose membrane (Bio-Rad), and immunoblotted overnight at 4°C with one of the primary antibodies: anti-Wnt11, or anti-β-actin (Cell Signaling). The membranes were then incubated with HRP-conjugated secondary antibody, washed, and developed with the ECL plus kit (GE Healthcare). Blots were analyzed by densitometry with NIH image software (AlphaEase FC, Version 6.0.0).
Co-culture CMs with MSCs
Native CMs were isolated from ventricles of 1- to 3-day-old SD rats using a commercially available neonatal CM isolation kit (Worthington Biochemical) as previously described [24]. To selectively enrich myocytes, dissociated cells were preplated for 2 h to allow nonmyocytes to attach to the bottom of the culture dish. The resultant suspension of myocytes was transferred onto collagen-coated culture dishes. To assay the cardioprotection, CMs were either co-cultured with MSCs in a dual-chamber system at a ratio 10: 1 (CMs: MSCs) or treated with CdM 16 h before cells were exposed to hypoxia.
Cell hypoxic injury
To mimic ischemic injury, cultured CMs were placed for 24–72 h in a hypoxic incubator (Sanyo, CO2/O2 incubator, MCO-18M), adjusted to 37°C and 5% CO2 with 94% N2 and 1% O2. Apoptotic cells were labeled with annexin-V-PE (Roche), examined under Olympus BX 41 microscope equipped with a digital camera (Olympus), and counted by fluorescence-activated cell sorting. Cell injury was estimated by measuring lactate dehydrogenase (LDH) release with a commercially available kit (Biovision). Cell survival was evaluated by MTT uptake (Promega).
Acute myocardial infarction model
Myocardial infarction (MI) model was created in 2- to 3-month-old SD female rats by permanent ligation of left anterior descending coronary artery (LAD). Animals were anesthetized with ketamine (100 mg/kg, I.P.) and xylazine (5 mg/kg, I.P.), and then mechanically ventilated. A left thoracotomy was performed at fourth intercostal space. The heart was exposed and LAD was ligated by a 7-0 ethicon suture just below the atrioventricular border. The chest was then closed, and the animals were maintained on buprenorphine (0.5 mg/kg, S.C.) for 24 h to alleviate pain. Sham surgery animals underwent the same surgical procedure except LAD ligation.
Animals were randomly divided into 5 groups: 1 sham surgery control and 4 MI groups receiving different treatments. The MI hearts were intramyocardially injected with medium (50 μL), MSCbas, MSCNull, or MSCWnt11 (1.5×106 cells in 50 μL medium) in ischemic border immediately after LAD ligation.
MSC survival in ischemic myocardium
The animals were sacrificed on day 4 post-MSC transplantation. The presence of surviving GFP+ donor MSCs was observed by GFP staining. Further, the number of survived Sry gene expressing male MSC in female recipient hearts was evaluated by real-time polymerase chain reaction (PCR) as previously described [26].
Assessment of cardiac function
Echocardiography was performed 4 weeks post-ligation using HDI 5000 SonoCT (Phillips) with a 15-MHz probe. Left ventricle (LV) parameters were obtained from 2-dimensional images and M-mode interrogation in long-axis view. LV internal dimensions (LVID) were measured at both systole and diastole. LV fractional shortening (FS) and ejection fraction (EF) were calculated as: FS=(LVIDd−LVIDs)/LVIDd×100; EF=[(LVIDd)3- (LVIDs)3]/(LVIDd)3×100. The data were averaged over 3 consecutive cardiac cycles.
Evaluation of apoptotic CM and MI
The hearts were arrested in diastole stage and fixed in 4% paraformaldhyde. Each heart was cut into 6 slices of 1.5 mm from the apex to the base and embedded in paraffin. Sections (5 μm thick) from slice 2 to slice 5 in each heart were mounted on microscopic glass slides and stained with TUNEL and Masson Trichrome, respectively.
Apoptotic CMs were evaluated by TUNEL assay in serial paraffin sections with ApopTag kit (Chemicon) 7 days post-transplantation. The amount of apoptotic CM in the infarcted border area was expressed as the ratio of TUNEL-positive CM nuclei to the total number of CM nuclei.
Masson trichome staining was carried out to delineate fibrous tissue after MSCs were transplanted for 4 weeks. The stained sections were photographed using a digital camera (model D30; Hitachi). Infarct size was defined as the sum of the epicardial and endocardial infarct circumference divided by the sum of the total LV epicardial and endocardial circumferences using computer-based planimetry. LV anterior wall thickening was expressed as percentage of septum thickness. Quantitative assessment of each parameter was performed using image analysis software (version 1.6065; NIH).
Statistical analysis
The in vivo data were obtained by the investigator who was blind to the animal groups. The in vitro experiments were carried out in triplicate. Quantitative data were presented as mean±SEM. One-way analysis of variance with the Holm-Sidak method and/or Bonferroni correction (SigmaStat 3.1; Systat Software) was used to determine the significance of differences between experimental groups and the difference was considered significant if the P value was less than 0.05.
Results
Cardioprotection of MSCWnt11
First, we observed the morphological changes and LDH release from CM after exposure to hypoxia for various time periods. As shown in Fig. 1, few CMs were shrunk when cells exposed to hypoxia for 12 h. However, significant morphological changes were observed when CMs were incubated under hypoxic condition for 24 h. Based on cellular changes, we selected hypoxia for 24 h as a time point for apoptotic study. Most CMs were inversely injured and detached when CMs were treated under hypoxia for 48 h or longer (72 h). No cell survived and induced 15-fold increase in LDH release compared with normal control after CMs were exposed to hypoxia for 72 h. Moreover, no significant protection was detected in MSCWnt11 co-culture group against 48 h hypoxia in our preliminary studies. Thus, we selected 40 h as the time point to observe whether MSCWnt11 reduced cell death/increased CM survival.
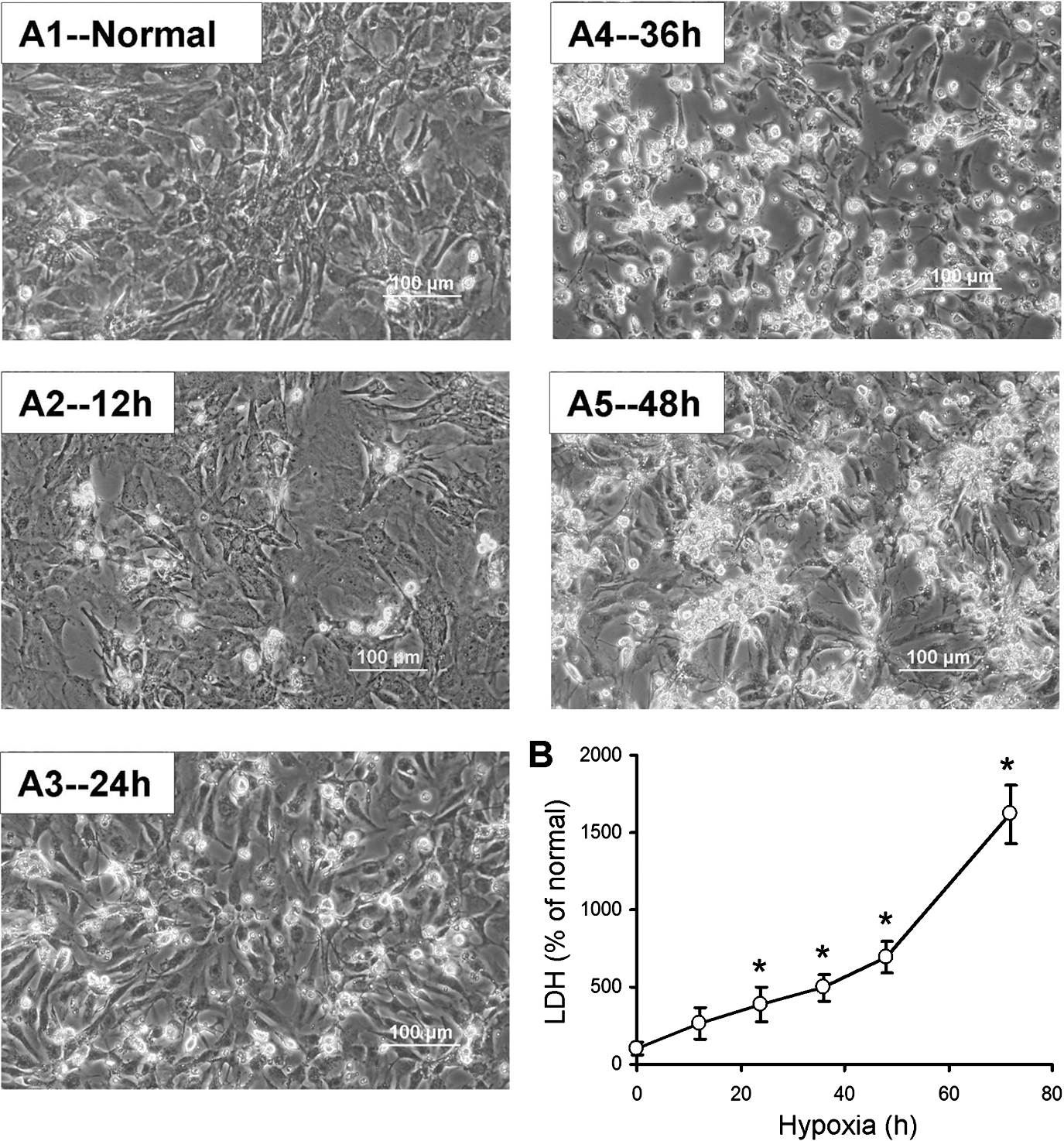
Morphology of cultured CMs
Second, primary cultured CMs were divided into 5 groups: normal control, hypoxic control, and 3 MSC co-culture groups to investigate whether MSCWnt11 confers a cardioprotective effect. After 24 h cells exposure to hypoxia, the number of annexin-V-positive cells in CM cultured alone was significantly increased, whereas their number significantly reduced when co-cultured with MSCWnt11 (Fig. 2). CM cultures exposed to hypoxic conditions for 40 h exhibited shrunken morphology associated large number of dead cells, increased LDH release, and reduced MTT uptake (Fig. 3). On the contrary, co-culture with MSCWnt11 significantly protected CM morphology, decreased LDH release, and increased MTT uptake.
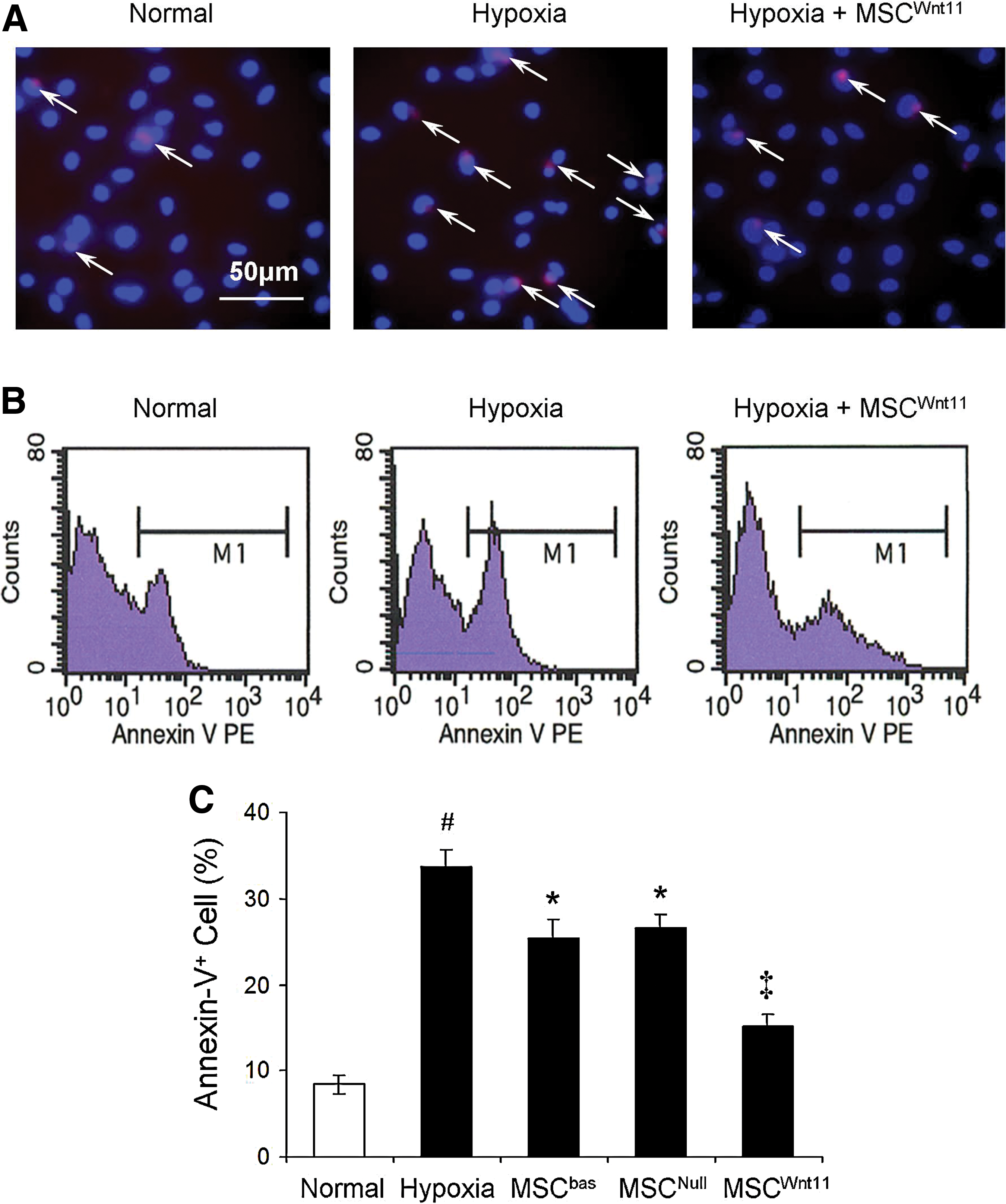
MSCWnt11 prevented CM apoptosis after exposure to hypoxia for 24 h in dual-chamber system.

Cytoprotective effect of MSCWnt11 on hypoxia-treated CM for 40 h in dual-chamber system.
Third, CMs were pretreated with concentrated CdM obtained from MSCWnt11 (CdMWnt11), MSCbas (CdMbas), or MSCNull (CdMNull) for 16 h before they were exposed to hypoxia to investigate whether the observed cardioprotection was due to secretion of Wnt11 and other factors from MSCWnt11. The ratio of CdM to fresh medium was 1:20. MTT uptake was significantly increased in CMs treated with CdMWnt11, compared with that treated with CdMbas and CdMNull (Fig. 4A).

Cardioprotection of CdM obtained from various MSCs.
Fourth, to identify the factors underlying MSCWnt11-mediated cardioprotection, gene expression in MSCWnt11 was assayed using large-scale microarray and the secreted factors were analyzed in CdM. Approximately 400 transcripts were novel and upregulated more than 1.5-fold in MSCWnt11 compared with MSCNull under hypoxic (24 h) condition. Thirty transcripts including TGFβ2 were upregulated more than 2.5-fold (Table 1). TGFβ2 expression was 2.81-fold higher in MSCWnt11 than in MSCNull under hypoxic conditions. Indeed, MSCWnt11 not only released Wnt11 (Fig. 4B), but also increased the release of TGF-β2, especially when they were exposed to hypoxia for 24 h (94.0±6.1 pg/mg protein in CdMWnt11 versus 66.0±9.5 pg/mg protein in CdMNull, P<0.05) (Fig. 4C).
NA, not available.
Finally, to confirm the cardioprotective effect of Wnt11 and TGFβ2, CMs were pretreated with Wnt11- or TGFβ2-neutralizing antibodies for 16 h before exposure to hypoxia. Wnt11-neutralizing antibody (6.0 and 12.0 μg/mL) and TGFβ2-neutralizing antibody (3.0 and 6.0 μg/mL) partially blocked the effect of CdMWnt11 on reducing LDH release and increasing MTT uptake (Fig. 4D, E). However, cytoprotection after treatment with both antibodies did not show a significant additive effect at the concentrations we used.
In vivo studies
MSCs and control medium were transplanted into female rat hearts immediately after development of MI. A total of 90 rats were used in this study, excluding bleeding during surgical procedures or animal death. Thirty rats were sacrificed on day 4 post-MI for assessing MSC survival in ischemic myocardium, 30 rats were utilized at 1 week post-LAD ligation for TUNEL staining, and another 30 rats were utilized at 4 weeks after different treatments to evaluate contractile function and MI.
MSCWnt11 survival in ischemic myocardium
The fate of male GFP-positive MSC in female rat myocardium was observed by GFP staining and quantified by real-time PCR for sry gene expression on day 4 after respective treatments. No GFP-positive cells were seen in medium-transplanted rat heart samples. However, we found many GFP-positive cells in MSC-transplanted rat heart (Fig. 5A). Male MSC were further quantified by real-time PCR for sry gene expression (n=4). No sry gene was detected in normal myocardium as well as medium-treated animals. The expression of Sry gene was significantly higher in MSCWnt11-transplanted animal hearts (Fig. 5B, C). The number of MSC was significantly increased in MSCWnt11-transplanted hearts compared with MSCNull-transplanted hearts (Fig. 5D). All of these results suggest that overexpression of Wnt11 significantly increased MSC survival in the ischemic environment.
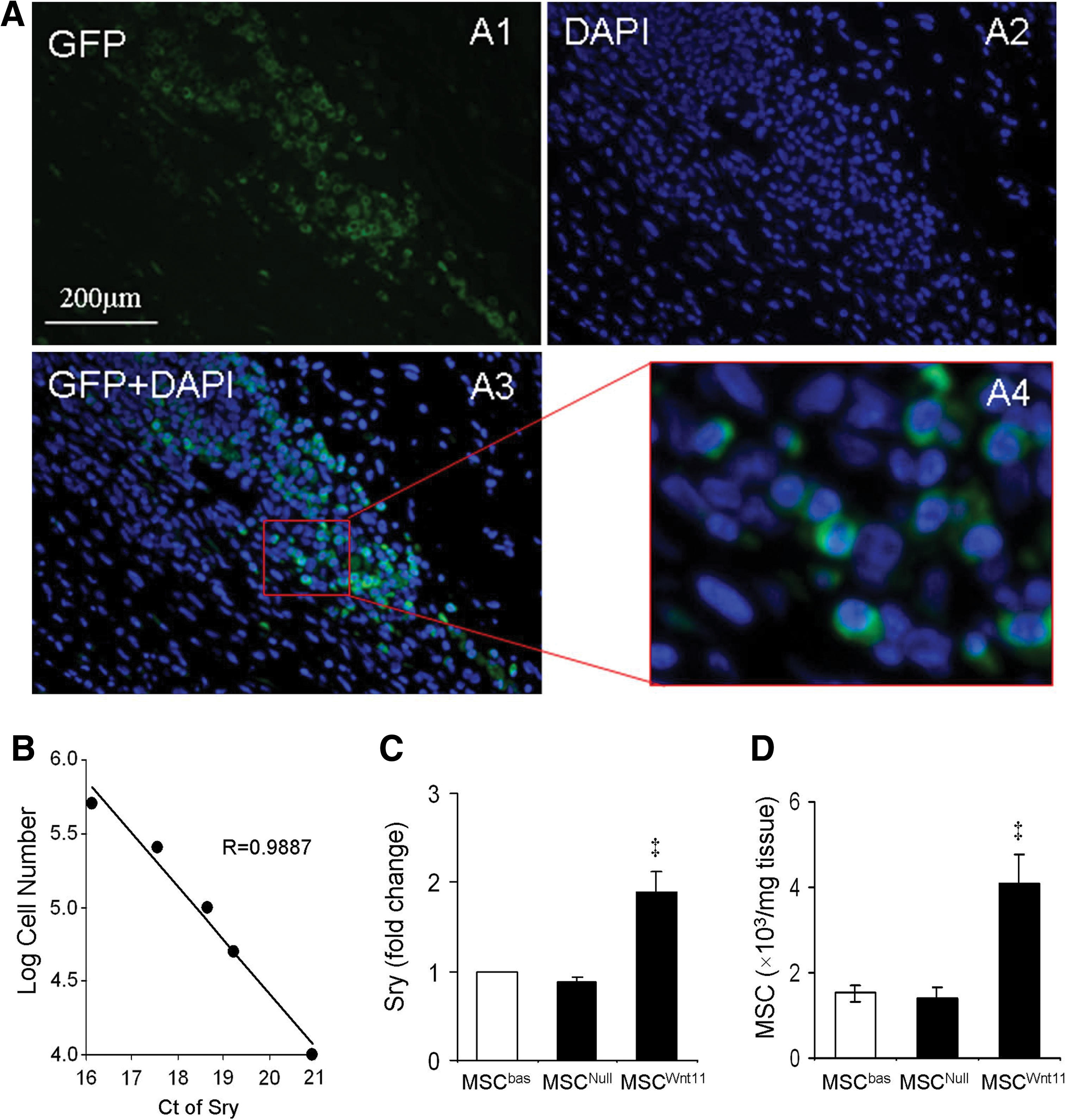
MSC survival in ischemic myocardium at day 4 post-transplantation. Panel
Cardiac function recovery and myocardial protection
The temporal changes in global heart function were measured in 6 animals per group using echocardiography after MSCs were transplanted into ischemic myocardium for 4 weeks. EF and FS were significantly reduced in LAD-ligated animals. The animals transplanted with MSCWnt11 showed a remarkable improvement in EF and FS as compared with other groups (Fig. 6A). Masson trichrome staining showed that MSCWnt11-transplanted animals had significantly reduced infarct size (30.1%±3.0% vs. 44.6%±4.1% in control, P<0.05) and increased LV anterior wall thickness. No infarction was observed in sham operation animals (Fig. 6B, C). TUNEL-positive cells in the ischemic border area were reduced significantly in MSCWnt11-treated animals (5.3%±0.35%), compared with the animals treated with control MSCs (MSCNull and MSCbas) (6.8%±0.46% and 6.4%±0.54%) and medium (9.9%±0.71%) (Fig. 7). This suggests that the cardiac function recovery is positively associated with MSCWnt11-mediated cardioprotection.
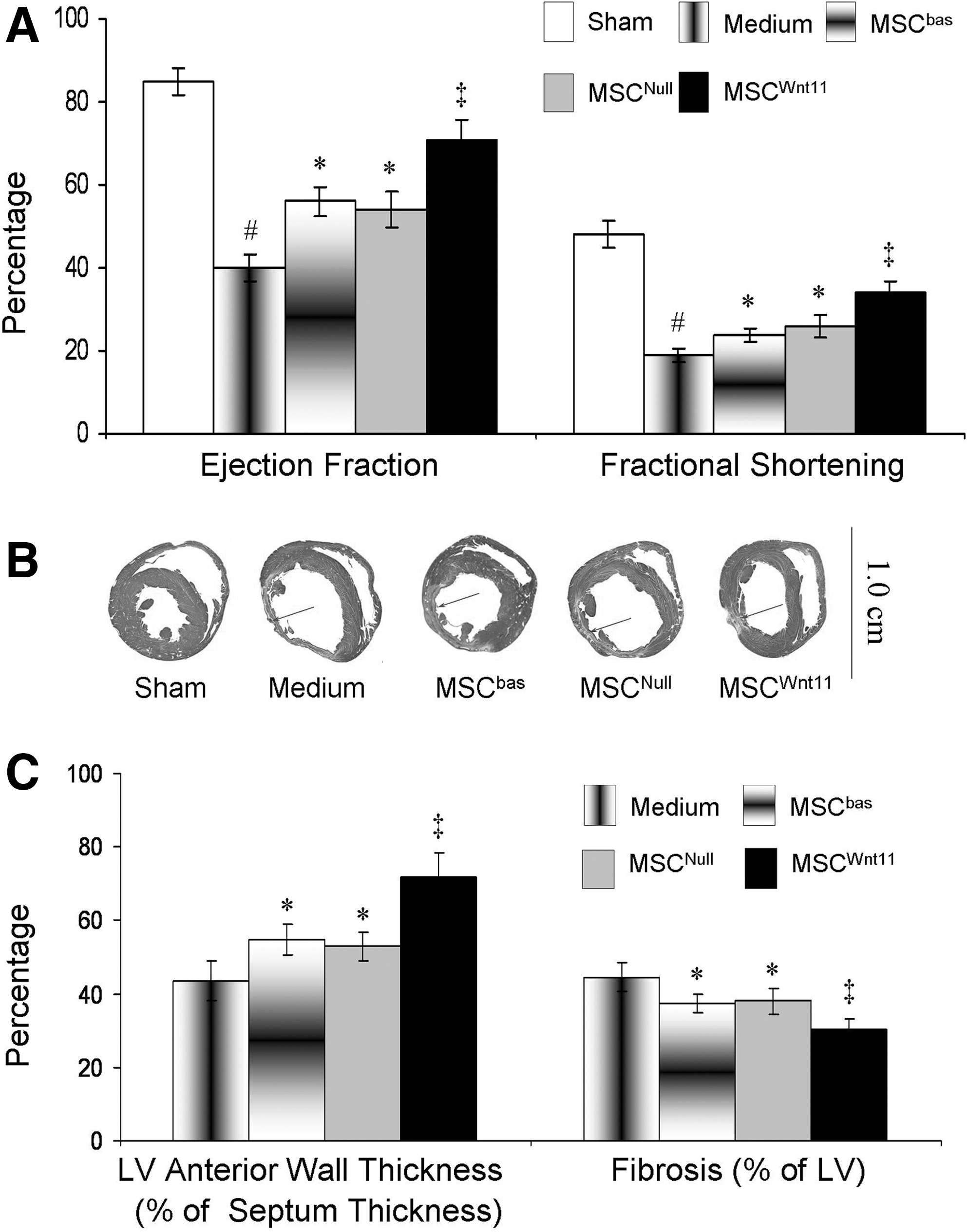
MSCWnt11 improved cardiac function and reduced infarcted myocardium size 4 weeks after they were transplanted into ischemic border following left anterior descending coronary artery ligation.
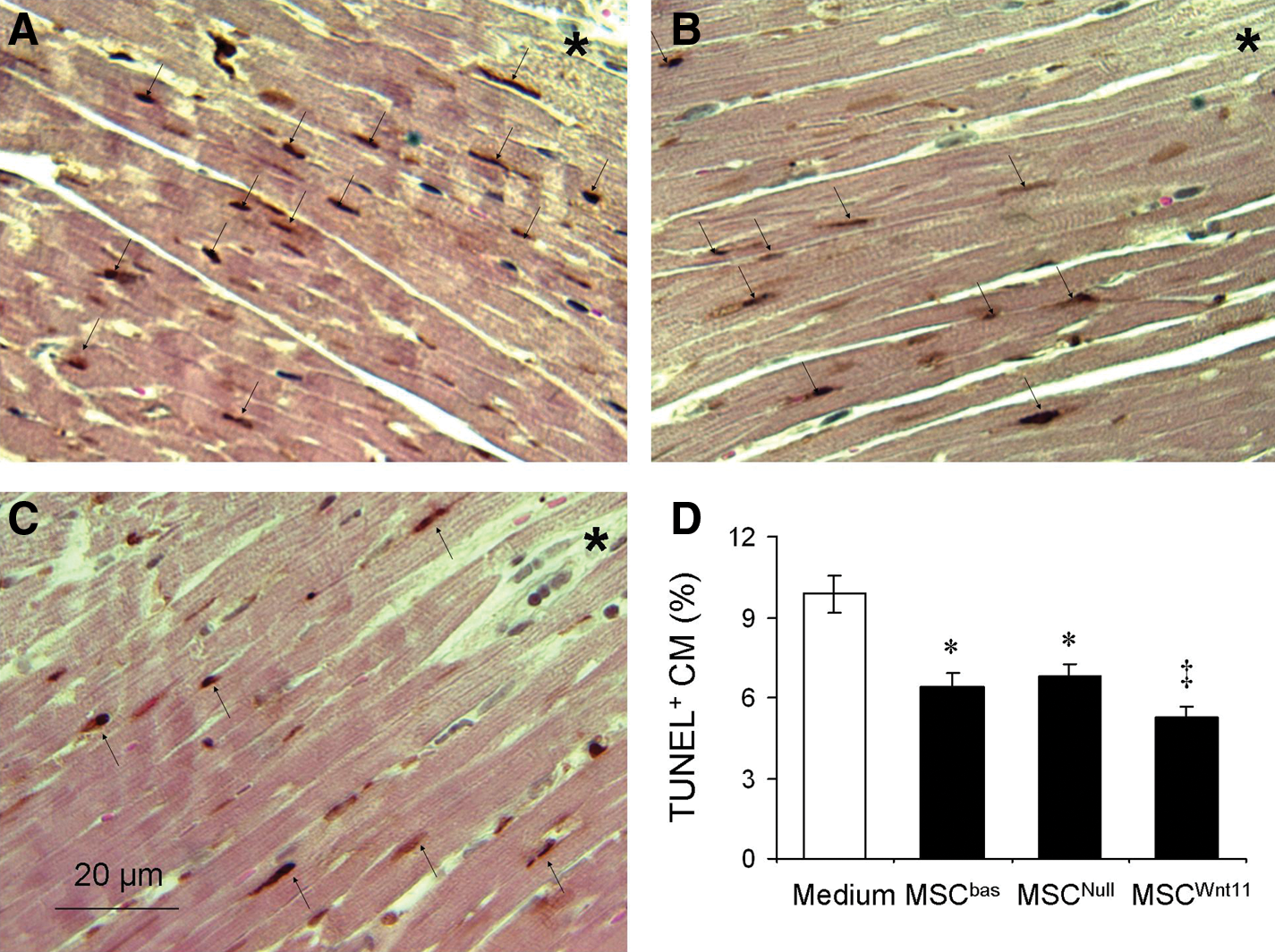
Apoptotic CMs in the peri-infarct area 7 days after myocardial infarction.
Discussion
This study makes 2 contributions to stem cell-mediated therapy of infarcted myocardium. First, gene-engineered MSCs secreted Wnt11 that conferred a cardioprotection against oxidative stress. Second, Wnt11 delivery with MSC protected native CM in ischemic environment and improved cardiac function.
More than a decade of research in experimental models indicates that delivery of stem cells into ischemic myocardium regenerates injured tissue, improves cardiac function, and protects tissue from further insult. We report a marked improvement in global performance together with a reduced apoptosis, fibrosis, and increased LV wall thickness after transplantation of MSCWnt11 into the border area of the ischemic heart compared with control MSCs. Successful cell therapy depends on survival of the transplanted cells. We transplanted GFP-positive male MSCs into female recipients to facilitate the detection of GFP-positive cells and quantified Y chromosome in the explanted female hearts. To our knowledge, this is the first study to report that Wnt11 significantly increases the survival rate of MSC in ischemic myocardium.
The improvement of cardiac function mediated by MSCWnt11 could be due to regeneration of the damaged myocardium. It has already been reported that Wnt11 promotes unfractionated bone marrow mononuclear cells or circulating progenitor cells transdifferentiation into myocardial phenotype [21,22]. More recently, it is reported that non-canonical Wnt signals promote myocyte maturation through caspase-mediated inhibition of β-catenin activity [12]. We have recently demonstrated that Wnt11 upregulated the expression of multiple cardiac genes in MSC, including GATA-4, brain natriuretic peptide, islet-1, and α-actinin [23]. Moreover, Wnt11 significantly increased the potential of MSC transdifferentiation into cardiac phenotype after co-culture with native CM for 7 days. The differentiation rate was significantly increased from 17.1%±3.1% in MSCNull to 26.1%±4.0% in MSCWnt11 [23].
Moreover, MSCWnt11 could contribute to the cardioprotection during acute ischemic phase. Several laboratories reported that multiple trophic factors released by MSCs attenuated tissue injury, inhibited fibrotic remodeling, as well as stimulated the recruitment and proliferation of tissue stem cells, and reduced inflammatory oxidative stress [25,29]. Our study indicates that MSCWnt11 significantly reduce CM injury, including prevention of apoptosis and reduction of cell death, both in vivo and in vitro when they exposed to ischemic microenvironment.
The severity of CM injury caused by hypoxia is time dependent. Apoptosis represents a crucial mechanism of CM loss in a great number of cardiac pathologies. Apoptotic cell death occurs in CMs after MI [30], and prevention of myocardial apoptosis may improve cardiac contractile function [30,31]. Cell death (or loss) in cultured CMs was mostly due to apoptosis, as evidenced by morphological changes, increased annexin V-positive cells, and LDH release.
The cardioprotection might be associated with secretion of Wnt11 and other paracrine factors from MSCWnt11. Two-chamber co-culture system study demonstrated that the paracrine factors released from MSCWnt11 can protect CM under ischemic environment. To verify the effect of factors released from MSCWnt11, CdM obtained from MSCWnt11 was used in CM culture system and showed significant cytoprotection capability. Present study provided the evidence that protective effect of paracrine factors released from the engrafted stem cells act on the host CMs and made them less vulnerable to apoptosis. These findings are consistent with the previous reports from other researchers that paracrine factors may be responsible for MSC-mediated cardioprotection [32]. To identify the factors responsible for MSCWnt11-mediated cardioprotection, we performed the microarray study. Approximately 800 genes were upregulated in MSCWnt11. Wnt11 and TGFβ2 were also significantly increased in CdMWnt11, and function-blocking antibodies to these 2 factors compromised the cardioprotective effect of CdMWnt11.
In this study, the isolation of CM directly from neonatal rat ventricles might yield a mixed population of many different cell types, including CMs, endothelial cells, and fibroblastic cells, although we isolated pure CMs using our previously described study to exclude nonmyocytes [24]. The observed effects in vitro (and probably in vivo) experiments may be related to other cells that need further experiments. Further, our preliminary studies also indicated that MSC could not protect CMs against hypoxia injury at a ratio of 1:40. All of these results suggested that the cardioprotection of MSCWnt11 against oxidative stress is limited.
Wnt11 has been shown previously an important mediator for the survival and/or viability of MCF-7 breast cancer cells [19] and CHO-K1 cells [20]. However, the role of Wnt11 in protection of CM remains unknown. It is considered that Wnt11 signaling promotes cell viability through coordination of Akt, JNK, and NF-κB key signaling pathways in cell survival and stress responses [20]. It has also been documented that Wnt11 signaling is sufficient to inhibit not only the canonical β-catenin-mediated Wnt signaling but also JNK/AP-1 and NF-κB signaling in the CHO cells [20]. Inhibition of the JNK/AP-1 pathway is mediated in part by the MAPK kinase MKK4 and Akt. We have previously reported that Wnt11 upregulated the expression of GATA-4 in MSC [23], which has been documented to upregulate the expression of growth factors [26], prosurvival factors (eg, Bcl-2), and several heat shock proteins, and cytokines (our unpublished data)—any of these could mediate the cytoprotective effects of GATA-4.
TGFβ superfamily plays essential roles in early embryonic development, cell mobility, growth, differentiation, and apoptosis [33,34]. TGFβ2, a key effector of organ morphogenesis, is reportedly regulated by Wnt11-mediated noncanonical signaling in developing heart and somites [35], and enhances efficacy of embryonic stem cell transfer for restoration of myocardial function [36]. However, the specific mechanism of TGFβ2 on cardioprotection is not clear, which renders future investigations.
In this study, we cannot exclude the possibility that other growth factors or proteins that were associated with Wnt11 upregulation are involved in MSCWnt11-mediated cardioprotection. Further study is needed to investigate the effect of individual paracrine factor.
Conclusions
Transplantation of MSCWnt11 improves LV function after acute cardiac ischemia. Overexpression of Wnt11 augments MSC-mediated cardioprotection. The underlying mechanism might involve direct cytoprotection of Wnt11 and paracrine factors secreted from MSCWnt11.
Footnotes
Acknowledgments
This work was supported by National Institutes of Health grants: HL083236, HL105176 (M.X.), and HL087246 (M.A.). Y-G.W. was supported by HL081859 and G-C.F. was supported by HL087861.
Author Disclosure Statement
There are no conflicts of interest.