
Abstract
Transplantation of retinal cells has recently provided a promising therapeutic approach for retinal degeneration. Here, we differentiated initially retinal progenitors (RPs) from adherent feeder-free human embryonic stem cells (hESCs) with the use of defined media supplemented with a specific combination of growth factors. The differentiated RPs highly (>80%) expressed related molecular features that included Six3 at an early stage in addition to Crx, Rx, Pax6, Otx2, and Chx10 at later stage. Next, we examined the induction of photoreceptors by Shh and/or the coculture of rabbit retinal pigmented epithelium with hESCs-derived RPs. The differentiation of retinal cells was demonstrated by protein and gene expression in all groups. However, S-Opsin, a cone photoreceptor marker, had higher expression in the presence of Shh, whereas expressions of Gli and Hes1 decreased in the same group. Finally, hESC-derived RPs were treated with Shh transplanted into the subretinal space of sodium iodate–injected albino-type adult rabbits and analyzed 4 weeks later. Transplanted retinal cells survived, migrated into retinal layers, and restored a small but significant B-wave. The grafted cells expressed photoreceptor markers, S-Opsin and Rhodopsin. Our results indicate that putative hESC-derived retinal cells express related genes and proteins. Further, our results show that retinal-like cells can be useful replacements for photoreceptors in retinal diseases.
Introduction
R
RPE is a densely pigmented epithelial monolayer present between the choroid and neural retina. This layer is critical for photoreceptor survival, phagocytosis of shed rods and cones in the outer segments, and secretion of several neurotrophic growth factors, which have important roles in the differentiation of photoreceptors and proper ocular development [21 –23]. On the other hand, during eye development, sonic hedgehog (Shh) is a regulatory factor involved in differentiation of RPs into photoreceptors [24 –29].
Therefore, in the present study, we initially differentiated RPs from adherent feeder-free hESCs. Next, Shh and/or a monolayer of in vivo derived RPE, prepared as an inducing cell layer that replicated the retinal microenvironment, were used to induce photoreceptor differentiation. Finally, the differentiated retinal cells from the Shh-treated group were transplanted into the subretinal space of sodium iodate–injected rabbits to evaluate both their ability to incorporate into retinas and improve functionality.
Materials and Methods
Culture of hESCs
The hESC line, Royan H5 [30], was used in these experiments. The cells were expanded and passaged on matrigel under feeder-free culture conditions in hESC medium that contained 100 ng/mL basic fibroblast growth factor (bFGF) as previously described [31]. The medium was changed every other day, until day 7.
Differentiation to RPs
The differentiation procedure is outlined in Fig. 1A. For neural differentiation induction, feeder-free hESCs were allowed to proliferate in hESC medium for 7 days. hESCs were subsequently induced to neural ectoderm in NE medium that contained noggin (1 ng/mL; R&D, 1976-NG), human recombinant Dkk-1 (1 ng/mL; R&D, 5439-DK/CF), and human recombinant insulin-like growth factor-1 (IGF-1, 5 ng/mL; R&D, 291-GI) in DMEM-F12 medium supplemented with 10% knockout serum replacement, 0.1 mM nonessential amino acids, 2 mM
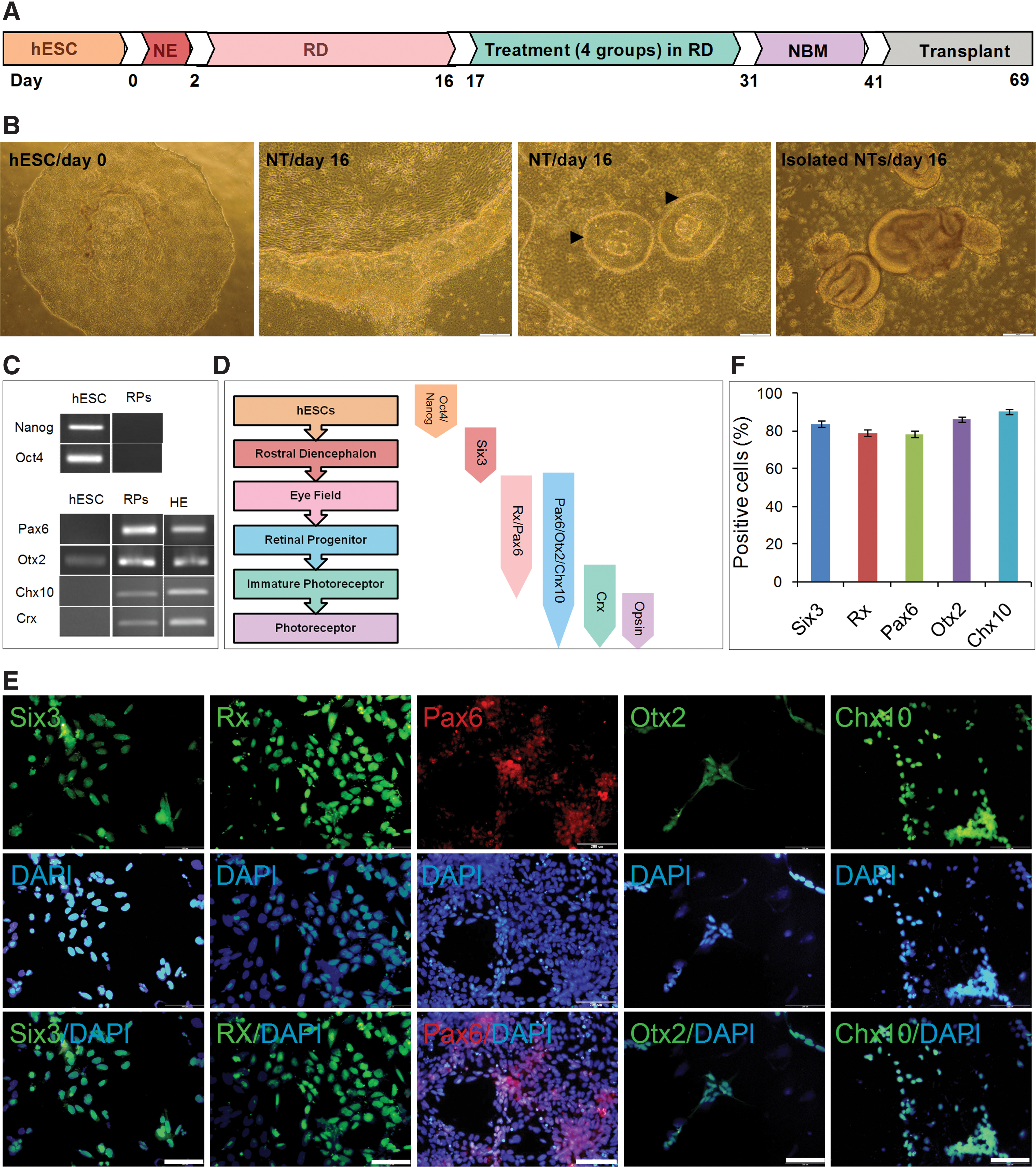
Generation of retinal progenitors (RPs) from human embryonic stem cells (hESCs).
Animals
Subjects of this study were albino and pigmented rabbits (Department of Physiology, Isfahan University, Iran) that weighed between 1.5 and 2.0 kg. All animal care, surgical processes, and postoperative euthanasia were undertaken in strict accordance with the approval of the Ethical Committee of Royan Institute. All surgical procedures were performed under anesthesia produced by a combination of ketamine (35 mg/kg) and xylazine (5 mg/kg).
Isolation and culturing of rabbit RPE
Pigmented rabbits were sacrificed by an overdose of ketamine and xylazine. Then, RPE was isolated according to established protocols [33]. In brief, the globes were enucleated and washed in Ca2+- and Mg2+-free phosphate-buffered saline supplemented with penicillin/streptomycin. The intact globe was incubated in 2% dispase (Gibco, 17105-041) for 20 min at 37°C. The incision of the globe was made 3 mm posterior of the limbus circumferentially. The anterior segment was discarded, and the posterior segment was extended by 4 incisions radially and incubated in DMEM/F12 supplemented with 10% FBS for 2 h at 37°C. The majority of the RPE sheet was lifted carefully from the choroid and neural retina as visualized under a stereo microscope (Olympus, SZX16). The isolated sheet was cultured on filter insert (0.4 mm; Millicell-CM-PICMO3050) in the presence of RD medium. Filter inserts were maintained in 6-well plates. The medium was changed every other day for 14 days.
MTS assay
To investigate whether RD medium affected survival of RPE cells, a cell proliferation assay was carried out by MTS assay. Briefly, 2×105 RPE cells were cultured in 96-well tissue culture plates under induction medium. Two weeks after plating, cell proliferation assay was carried out by MTS/PMS (Promega, G5421) according to the manufacturer's protocol. Absorbance was measured at 492 nm by an ELISA reader (Awareness).
Experimental design
One day after replating of dissociated NTs on laminin/poly-
Preparation of cells for transplantation
Immediately before transplantation, the hESC-derived retinal cells were harvested by trypsinization, washed 3 times in PBS, and resuspended the cells to the desired concentration. For identification after transplantation, we utilized human nuclear antigen (HNu), a selective nuclear marker of cells that originate from humans or PKH-26 (PKH26-GL; Sigma-Aldrich), according to manufacturer's protocol. PKH-26 staining provides the identification of grafted cells in vivo. To ensure labeling procedure, we examined the cells using fluorescence microscopy. A sample of the resulting single-cell suspensions was stained with trypan blue dye exclusion (Sigma) and counted with a Neubauer hemocytometer.
Surgical procedure, experimental grouping, and transplantation
Sodium iodate–induced model of RPE degeneration
A single injection of sterile 50 mg/kg solution of sodium iodate (NaIO3; Sigma) in saline was administered into the ear marginal vein of each rabbit [34,35]. After 7 days, cell transplantation was performed.
Experimental grouping and transplantation
Eighteen adult albino rabbits with normal eye function were selected and randomly assigned into normal and NaIO3-injured groups. The normal group (n=6) had normal, intact eye function and received no treatment. Seven days after NaIO3 injury, the animals were assigned into 3 groups. The sham group (n=6) received DMEM/F12 in the subretinal space. hESC-derived retinal cells were transplanted (transplanted group) into the left eyes of 6 other NaIO3-injured animals and the right eyes of the same animals were used as controls that underwent no additional manipulations.
For sham or cell transplantation, NaIO3-treated albino rabbits were anesthetized with ketamine and xylazine. To access the subretinal space, small supra nasal conjuctival peritomy was performed; the conjuctiva and tenon were separated from beneath the sclera. A 90% partial thickness sclera incision was made at a distance of 7–8 mm from the limbus. Before transplantation, confirmation of cellular viability was performed with trypan blue exclusion dye. A total of 50,000–80,000 cells in 1.5 μL of DMEM/F12 were injected into the subretinal space using a pulled fine glass micropipette. The fundus and elevated retina were visualized with a 35° lens. The conjunctiva was sutured with Vicryl 7.0. Starting 2 days before transplantation, all groups of rabbits in transplanted/control and sham groups received daily injections of cyclosporine-A (20 mg/kg, s.c., Sandimmune; Novartis Pharmaceuticals) [36] for a total of 4 weeks to prevent rejection.
Electroretinography
Rabbits were dark-adapted for at least 4–5 h, anesthetized with ketamine and xylazine, and prepared for recording electroretinography (ERG). Pupils were dilated with Midrax (Tropicamid) 5 min before ERG recording. A gold ring electrode embedded in a contact lens was placed on a drop of 2% methyl cellulose on the cornea. Ground and reference copper electrodes were placed in the forehead and lateral conthus, respectively. Three different test flash intensities were sent and responses collected from each rabbit. ERGs were carried out under scotopic, phototopic, and Max conditions. ERG signals were amplified 5,000, filtered between low-pass filter 30 Hz–10 kHz and high-pass filter 0.02–10 kHz.
Sacrifice and cryosection
After ERG analysis, all rabbits of the sham and control/transplanted groups were sacrificed. Rabbit eye globes were dissected and postfixed at 4°C and then cryoprotected with 30% phosphate-buffered sucrose at 4°C for 96 h. Samples were longitudinally cryosectioned at 10 μm and stored at −20°C until staining.
Immunocytofluorescence and immunohistofluorescence staining
Immunocytofluorescence staining was performed as a standard protocol using the antibodies listed in Table 1. Labeled cells were visualized with either FITC- or TRITC-conjugated secondary antibodies, and nuclei were stained with DAPI (Sigma, D8417). A fluorescence microscope (Olympus BX51, Japan) equipped with an Olympus DP70 camera was used to evaluate the number of positive cells labeled with retinal cell markers out of approximately 500 DAPI nuclei-labeled cells.
For immunohistofluorescence staining, sections were rehydrated in 0.1 M PBS and permeabilized with 0.4% Triton X-100 in PBS. After blocking nonspecific sites with 10 mg/mL bovine serum albumin, sections were incubated overnight with primary antibodies. Antibodies implemented for immunohistofluorescence are presented in Table 1.
For survival studies, we utilized anti-HNu and PHK-26. Normal IgG from the species of origin of the primary antibodies served as negative controls. After washing with PBS, the cells were incubated with the appropriate FITC- and TRITC-conjugated secondary antibodies for 1 h at 37°C. Sections were washed 3 times with PBS and counterstained with DAPI. The cover slips were mounted onto glass slides with Entellan. The same fluorescence microscope and photography was used to photograph positive cells that were labeled with different cell markers.
Reverse transcription–polymerase chain reaction and real-time reverse transcription–polymerase chain reaction for gene expression analysis
Total RNA was extracted from cultured cells using the QIAprep Spin Miniprep kit (QIAGEN, 27106) according to the manufacturer's protocol. Prior to reverse transcription (RT), RNA samples were digested with DNase I (Fermentas, EN0521) to remove contaminating genomic DNA. Standard RT was performed using 2 μg total RNA, oligo (dT) 18, and the Revert AidTM H Minus First Strand cDNA Synthesis kit (Fermentas, K1622) according to the manufacturer's instructions. Primer sequences, annealing temperature(s), cycles, and the lengths of amplified products are shown in Supplementary Table S1 (Supplementary Data are available online at
Real-time RT-PCR was performed using a Thermal Cycler Rotor gene 6000 (Corbett) with 10 μl SYBR Green PCR Master Mix (Takara), 5 pM of each primer, and 50 ng cDNA for each reaction in a final volume of 20 μl. Relative gene expression was analyzed using the comparative Ct method (2−ΔΔCt) [37]. All samples were normalized to levels of GAPDH, which was used as the loading control. All measurements were performed in triplicate. Real-time specific primer pairs (Supplementary Table S2) were designed by Beacon software (Version 7.2).
Western blot analysis
TRI reagent (Sigma-Aldrich) was used to extract protein from induced cells. Protein concentration was determined with a Bradford assay (Bio-Rad) kit. The prepared samples were electrophoresed on 12% SDS-PAGE gels (Bio-Rad, Munich, Germany) and then transferred to PVDF membranes (Bio-Rad, Munich, Germany). Membranes were blocked overnight in 10% skim milk (Merck) and incubated with primary different antibodies including S-Opsin (1:2000) and β-Tubulin (1:2500; Sigma-Aldrich, T5293; housekeeping gene or internal control) for 90 min. The blots were washed and then incubated with horseradish peroxidase–conjugated goat anti-rabbit IgG (1:8000) for 1 h. Bands were visualized after further washes using an ECL advanced western blot detection kit (GE Healthcare).
Statistical analysis
All values are stated as means±standard error of mean (SEM). Data were analyzed by 1-way ANOVA followed by LSD post hoc, with a significance threshold of P<0.05.
Results
Generation of RP cells from hESCs
The undifferentiated feeder-free hESCs were maintained in the presence of a high concentration of bFGF (100 ng/mL) on Matrigel (Fig. 1B). Then, the cells were differentiated into RPs, using a published protocol [32], without embryoid body (EB) formation in adherent conditions. The hESCs were treated with noggin, Dkk-1, and IGF-1 for 2 days. Next, the cells were treated in RD medium for an additional 2 weeks. Consequently, the neuroepithelium and NTs emerged from hESCs (Fig. 1B). At day 16, NTs were mechanically isolated under a phase-contrast microscope, replated on laminin/ornithine-coated plates, and cultured in RD medium for 1 day. RT-PCR analysis of differentiated cells at day 17 (= 2+14+1 days) showed that the cells lost expression of pluripotency genes, Oct4 and Nanog (Fig. 1C), and acquired expression of eye field genes (Fig. 1C, D). The efficiency of retinal determination was tested by analyzing the expression of several key eye-field transcription factors at the protein level (Six3 at day 9 and Rx, Pax6, Otx2, and Chx10 at day 17).
During early eye development, Six3 is first expressed in the undifferentiated neuroepithelium and then its expression is downregulated in the differentiating neural retina and postmitotic photoreceptors. We first examined the most rostral marker, Six3 (Fig. 1E, F). Quantification of immunostaining revealed that 83.8%±1.7% of differentiated cells were positive for Six3 (Fig. 1F). The immunostaining for Six3 was negative at day 17 (data not shown). For specification of Six3 rostral CNS, the transcription factor Rx (an early bona fide marker for the eye field) is needed and immunostaining results revealed that around 79%±1.6% of cells express this marker (Fig. 1E, F). Additionally, coexpression of Rx with Pax6 characterized the neural retinal lineage and 78.3%±1.8% of the cells expressed Pax6 (Fig. 1E, F). In the neural retina, Otx2 is expressed in postmitotic anterior neuroblast and acquires the ability to differentiate to various cell types. A total of 86.2%±1.6% of the differentiated cells expressed Otx2 (Fig. 1E, F). In early eye development, during optic vesicle and cup formation, the expression of Chx10 in the inner layer of the optic cup becomes gradually apparent. Thus, Chx10, considered as the earliest specific marker for RPs, was observed following the expression of Otx2 (Fig. 1E). Quantitative analysis of the cultures showed that 90.53%±1.3% of the cells were positive for Chx10 using immunofluorescence staining (Fig. 1F). The expression of Crx, a photoreceptor marker, was confirmed by RT-PCR (Fig. 1C).
Therefore, a large fraction of the hESCs in cultures differentiated to RPs when maintained under RD conditions.
Coculture of RPs with RPE and/or Shh factor
To improve the generation of neural photoreceptors from hESC-derived RPs, the RPs were cultured in the presence and absence of Shh and/or RPE in RD medium (see later).
Before the coculture of RPs with rabbit RPE, we evaluated the preserved identity and viability of cultured RPE after 14 days. The RPE cells maintained pigmented hexagonal morphology and expressed cytokeratin 18, an intermediate protein, ZO-1, the marker of tight junction (Fig. 2A), and natriuretic peptid receptor (NPR-A), a rabbit RPE marker [38] (Fig. 2B). In addition, MTS assessment showed the maintenance of RPE viability in the in vitro condition (Fig. 2C). Therefore, RPE maintained their viability and identity during culture.

Isolated retinal pigmented epithelium (RPE) in vitro and experimental design.
At day 17, RPs were cocultured in 4 groups (C: control/without treatment; R: RPE only; S: Shh only; R+S: RPE+Shh; Fig. 2D). After 14 days (day 31), some of the differentiated cells showed typical neuronal morphology with extended neurites (Fig. 3A) and further differentiation was achieved by an additional 10 days of culturing in NBM in all groups (day 41; Fig. 3B). RT-PCR analysis of the differentiated cells at day 41 revealed expression of the following photoreceptor markers in all groups: Recoverin, a later photoreceptor marker, Nrl and Nr2e3, rod photoreceptor transcription factors, Rhodopsin, a rod photosensitive pigment, and S-Opsin, a cone photoreceptor marker (Fig. 3). The differentiated cells also expressed Pax6, which is characteristic of ganglion and amacrine cells, and PKCα and Chx10, bipolar cell markers (Fig. 3C). Quantitative RT-PCR and western blot demonstrated that S-Opsin significantly increased (P<0.05) in the S group (Fig. 3D, E). At day 41, we further quantitatively analyzed expressions of Gli and Hes1, 2 mediators downregulated by Shh in the differentiation of RPs to both rod and cone cells. The results revealed significant reduction in expression of both factors upon removal of Shh and further culture in NBM (Fig. 3F). No significant difference was seen in expressions of Nr2e3 and Rhodopsin in all groups as indicated by qRT-PCR (Fig. 3H, I). However, Nrl was expressed more in the RPE coculture (Fig. 3G). Assessment of differentiated cells by immunofluorescence staining revealed that they were positive for Rhodopsin and S-Opsin (Fig. 4 and Supplementary Fig. S1) in addition to Lim1/MAP2, GFAP, and Chx10/MAP2 for horizontal, muller, and bipolar cell markers, respectively (Fig. 4). These markers have been previously described in retinal neurons of various species, including the human retina.
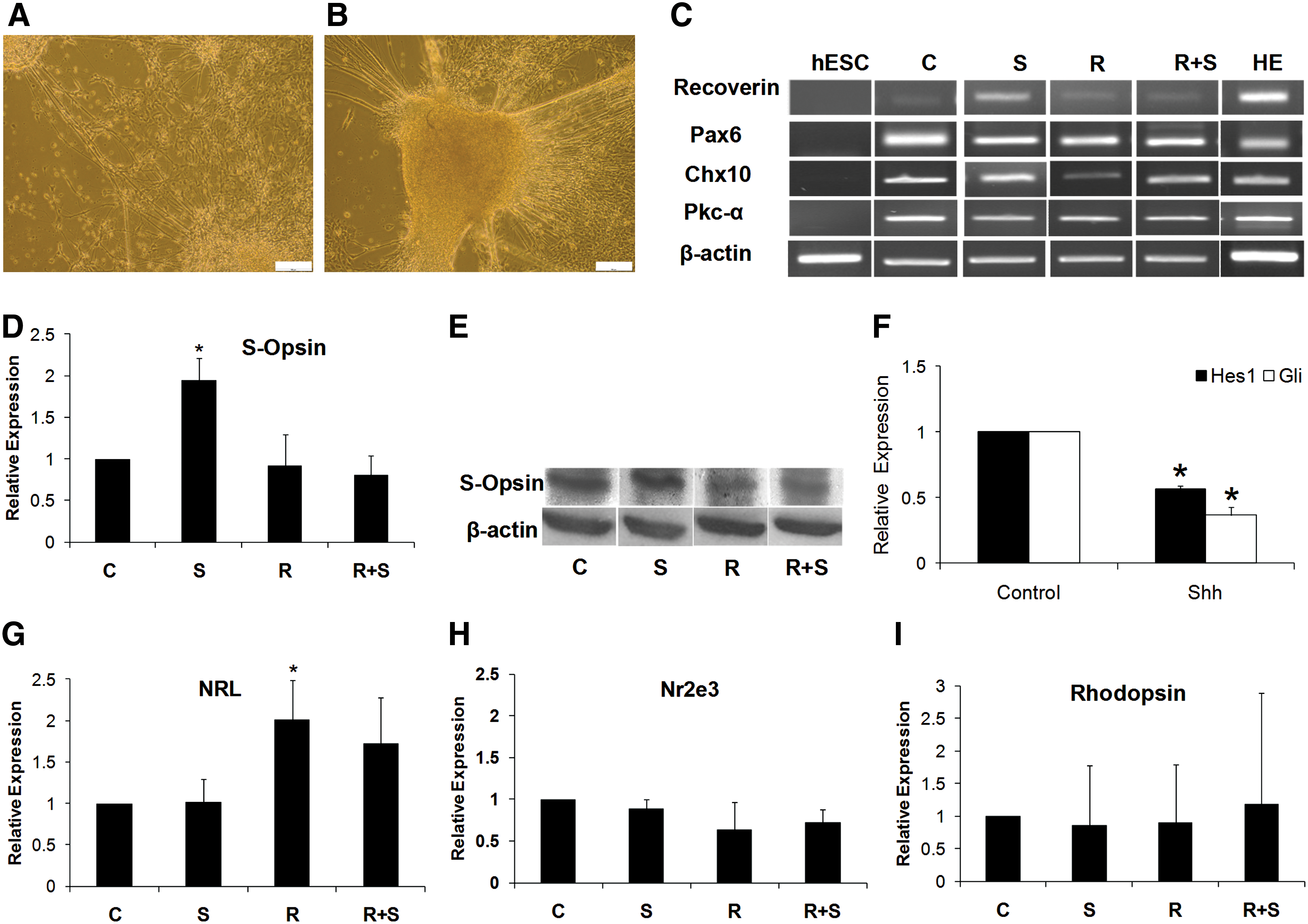
Transcript analyses of RP-derived retinal cells under different treatments at day 41. Representative phase-contrast micrographs of differentiating retinal cells at day 31

Representative immunofluorescence staining of RP-derived retinal cells in Shh group at day 41. The retinal cells expressed Rhodopsin and S-Opsin, photoreceptor markers. Cells colabeled with horizontal cell marker Lim1 (green) and the neuronal marker Map2 (red) are shown. Immunostaining showed presence of glial cell with GFAP marker and bipolar Chx10 cells coexpressed Map2. Nuclei were counterstained with DAPI. Color images available online at
In vivo transplantation
Next, to evaluate whether retinal cells derived from hESCs improve the function of damaged retina, we transplanted retinal cells derived from the Shh group, which expressed more S-Opsin into the NaIO3-injected rabbit model of RPE degeneration. Cyclosporine A was injected daily to preclude rejection of the human cells by the host. The cells were dissociated and immediately transplanted into the subretinal space of adult animals. Examination of histological sections of the retina was conducted in 4 groups as described earlier. Seven days after injection, the outer nuclear layer was disorganized and the numbers of RPE and photoreceptor cell nuclei were decreased (Fig. 5A). Transplantation success was microscopically indicated by the presence of a retinal bleb in the fundus of the eye immediately after injection.
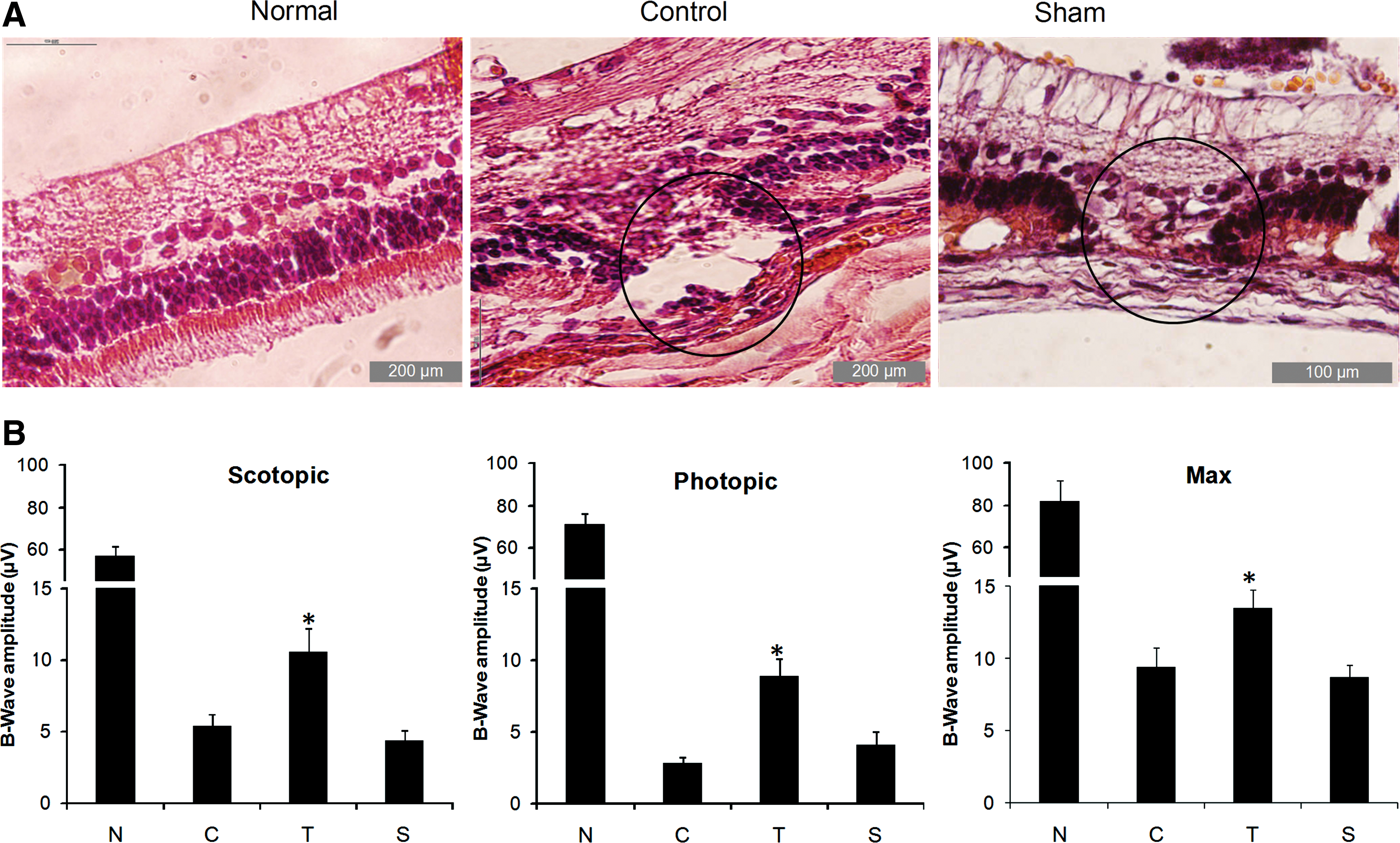
Histology and electroretinography of NaIO3-injured rabbit model.
After a survival period of 4 weeks, ERG analysis of the animals showed that scotopic (dark-adapted), photopic (light-adapted), and Max B-wave peak amplitudes were reduced in control groups, which received NaIO3 in comparison with normal animals (Fig. 5B and Supplementary Fig. S2). Four weeks after cell transplantation, a small light response was observed in comparison with the control group, which was unresponsive. Additionally, the scotopic, photopic, and Max B-wave peak amplitudes increased in the cell-transplanted animals when compared with the control and sham groups under similar excitation conditions (B-wave amplitude of scotopic: 10.6±1.7, photopic: 8.9±1.2, and Max: 13.4±1.3 μV in the cell-transplanted group compared with scotopic: 5.4±0.8, photopic: 2.8±0.4, and Max: 9.4±1.4 μV in the control group; P<0.05; Fig. 5B). ERG results showed that the mean B-wave amplitude following light flash in sham surgery was not significantly different from the control group after 4 weeks (scotopic: 4.4±0.7, photopic: 4.1±0.9, and Max: 8.7±0.8 μV in the sham group; Fig. 5B).
To evaluate transplanted cell survival, migration, and fate, rabbits were euthanized and their eyes removed, fixed, and sectioned after 4 weeks. The transplanted retinal cells were identified by lipophilic red dye (PKH26) labeling or immunofluorescence for a selective marker of human cell (HNu) and other photoreceptor markers. We found that HNu colabeled with PKH hESC-derived retinal cells survived in the subretinal space and migrated into the outer and inner nuclear layers and ganglionic cell layer of the retina (Fig. 6A–C) over the 4-week period. Therefore, histological analysis following ERG testing confirmed that the animals that demonstrated a response to light had a successful transplant. Of note, in 2 of the 6 rabbits we could not observe any PKH- or HNu-positive cells in their injected eyes perhaps because of apoptosis; these 2 rabbits did not present detectable ERG and were therefore excluded from analysis.

Tracing of transplanted cells in retinal sections at 4 weeks after cell transplantation.
To determine the phenotype of PKH-positive cells, immunoflourescence was performed for Rhodopsin and S-Opsin. Examination of sections revealed that the majority of the transplanted cells were observed in the outer nuclear layer and expressed Rhodopsin and S-Opsin markers for rod and cone cells, respectively (Fig. 6D, E).
Therefore, a proportion of the transplanted hESC-derived retinal cells are light responsive and may transmit this response to the bipolar cells.
As undifferentiated hESCs can give rise to teratomas, we also assessed all eyes for evidence of teratoma formation. However, no teratomas were observed in any of the rabbits that received transplanted cells during our study. It is probable that if teratomas were to arise, they would be derived from undifferentiated hESCs. It is, however, improbable that any of the cells remained undifferentiated after 41 days of retina-directed cell culture prior to transplantation.
Discussion
In concordance with results of Lamba et al. [32], we have shown that hESCs can differentiate efficiently into RPs using defined media supplemented with a specific combination of growth factors, which included noggin, a BMP antagonist, Dkk, a Wnt antagonist, and IGF-1. The differentiation was induced as an adherent monolayer culture that highly (over 80%) expressed molecular features of RPs, such as Six3 at the early stage and Crx, Rx, Pax6, Otx2, and Chx10 at later stage. In contrast to the protocol of Lamba et al. [32], which used EBs at the initial step, we maintained hESCs in the adherent system on a matrigel matrix without a feeder layer. Because the differentiation process through the EBs is spontaneous and somewhat stochastic, the formation of EBs leads to the formation of numerous cell types originating from all 3 germ layers [39]. Our findings indicate efficient production of RPs of relatively high purity. Moreover, the adherent culture protocol facilitates visualization of the process of neural conversion and NT formation. Additionally, many cells located inside the EBs are in an apoptotic status, indicating that most ESCs die instead of differentiating and producing healthy ESC-derived cells.
Considering the fact that both RPE/Shh are required for retinal development [21 –29], and recently, as Yue et al. [10] have shown that coculturing of monkey ESCs with ESC-derived RPEs is a useful and efficient method for inducing photoreceptors, we evaluated whether RPE and/or Shh affect the differentiation of hESC-derived RPs in vitro, using a coculture system. Our results have shown that isolated RPE maintained their features, intercellular connections, intact tight junctions, intracellular microfilaments, and morphologic characteristics, which is important for their function. After RP maturation in all groups, the marker expressions of photoreceptors, ganglion cells, amacrine cells, bipolar cells, and horizontal cells were observed. Although we observed a significant increased expression of Nrl in the RPE group, the expressions of Nr2e3 and Rhodopsin were not significantly different between the groups. Additionally, RPE did not improve the expression of a cone photoreceptor marker, S-Opsin, but it enhanced in the presence of Shh only. This may be related to limited coculture time or the lack of a suitable time window for the coculture of RPE with RPs to induce mature photoreceptor differentiation, or further factors such as retinoic acid might be required to promote rod formation [10]. On the other hand, in retina, RPE sheet is closely attached to photoreceptor cells or their precursors and, therefore, more significant improvement could be gained when hESC-derived RPs are directly cultured onto RPE sheet.
In the presence of Shh, expressions of Gli and Hes1 decreased. It has been shown that removal or inhibition of Hes1 mainly leads to formation of cone cells [40] and further factors are required to deviate the default pathway and promote rod formation [41].
Previously, mouse photoreceptors have been shown to incorporate into adult retinas following transplantation into the subretinal space [5]. Therefore, to test whether hESC-derived retinal cells would similarly incorporate into the retinas and improve functionality of NaIO3-injured retinal model of adult rabbits, we injected the Shh-treated retinal cells into the subretinal space in albino-type adult rabbits and analyzed them 4 weeks later. We found that transplanted hESC-derived retinal cells can be integrated into adult host retina and can restore a small but significant B-wave, whereas sham injections or control did not. It was demonstrated that human retinal cells derived from either hESC or hiPSCs integrate following transplantation and restore visual function in animal models [17,18]. The recognition of donor cells labeled with PKH was confirmed by expression of HNu. Immunohistochemical analysis revealed integration of these cells to the outer nuclear layer (ONL), inner nuclear layer (INL), and ganglionic layer. Further, grafted cells expressed Rhodopsin and S-Opsin. The restoration of an ERG response correlates with the efficiency of transplantation. The transplanted retinal cells rescued visual function by the increased significant B-wave when compared with the control group. Interestingly, functional restoration of rabbits was detected in both dark and light responses. The mechanism of this improvement remains to be determined; however, it may be due to the attendance of a subpopulation of precursor or mature cells that survived, migrated, and differentiated to photoreceptors, thus improving visual function. A previous report has shown that the outer limiting membrane is a barrier within the adult retina to prevent migration of donor cells. Communication between host and graft tissues has been shown to occur through interruption of outer limiting membrane and laminar organization [42]. Correspondingly, local disruption probably facilitates migration of donor cells [43].
Taken together, as seen with other studies, our results indicated generation of retinal fate from adherent feeder-free hESCs. Following transplantation of Shh-committed retinal cells, a number of cells incorporated into the retina, differentiated, and effected a significant improvement in visual restoration to the rabbits. This provides a promising approach for replacement of cells in retinal diseases such as pigmentosa and age-related macular degeneration.
Footnotes
Acknowledgments
This study was funded by a grant from Royan Institute and Iranian Council of Stem Cell Technology.
Author Disclosure Statement
None of the authors has any conflict of interest to disclose and all authors support submission to this journal.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.