
Abstract
Induced pluripotent stem (iPS) cells are a potential cell source for regenerative medicine. However, the tumorigenicity of iPS cells is a big concern for clinical application. In addition to the genetic manipulation of the reprogramming process and the greater risk of tumor formation, it is unclear whether iPS cells with normal development potential are still tumorigenic. Here, we investigated 3 mouse iPS cell lines, including one line that is able to generate full-term mice via tetraploid blastocyst complementation. We found that a small number of undifferentiated iPS cells could be steadily isolated and expanded after long-term differentiation of cells in vitro or in vivo. The residual undifferentiated iPS cells could be expanded and redifferentiated, and undifferentiated pluripotent stem cells could again be isolated after further rounds of differentiation, suggesting that residual undifferentiated iPS cells could not be eliminated by extended cell differentiation. The residual undifferentiated cells could form teratomas in vivo, indicating that they are a potential tumorigenic risk during transplantation. These findings prompt us to reconsider the strategies for solving the tumorigenic problem of iPS cells, not only focusing on improving the reprogramming process.
Introduction
S
To date, most iPS cells have been generated by transduction of somatic cells with retroviruses or lentiviruses [2,3,6,14]. The random integration of transgenes and the integrated genes themselves that include potent oncogenes such as c-Myc and Klf4 could be great risks for tumor formation during clinical application. Such risks could be potentially reduced by using different reprogramming approaches, such as using nonintegrating adenoviruses, plasmids, recombinant proteins, or chemical compounds [15 –22]. However, it is unclear whether iPS cells with normal developmental potential are still tumorigenic. Our previous work revealed that a small number of nonmutant mouse ES cells could maintain their pluripotency during in vitro and in vivo differentiation, and that these contaminated undifferentiated ES cells are still tumorigenic during transplantation (unpublished data). Since iPS cells possess many characteristics of ES cells, understanding whether iPS cells with normal developmental potential possess the same ability to maintain a small number of cells in a pluripotent state during differentiation is important.
In this study, 3 iPS cell lines, including one that is able to generate full-term mice via tetraploid blastocyst complementation and others with the ability to produce chimaerism with or without germline transmission [23], have been tested. Two cell differentiation systems, the embryoid body (EB)-differentiation system and teratoma-formation system, were utilized to test whether residual pluripotent stem (rPS) cells exist after long-term differentiation of mouse iPS cells. We demonstrated that rPS cells did exist in long-term EBs and teratomas in all types of cells tested, and they could not be eliminated by extended cell differentiation. In addition, the rPS cells could be repeatedly isolated after multiple rounds of cell differentiation in vitro and in vivo.
Materials and Methods
iPS cell culture
Three mouse iPS cell lines (IP14D-1, with the ability to produce tetraploid complementation pups; IP14D-2, with the ability to produce chimaerism without germline transmission; and IP20D-3, with the ability to produce chimaerism with germline transmission) were obtained from Beijing Stem Cell Bank (Beijing, China) [23]. Cells were maintained on mitomycin-treated mouse embryonic fibroblast (MEF) feeder cell layers in Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 15% (v/v) fetal bovine serum (FBS; Invitrogen), 1,000 units/mL leukemia inhibitory factor (R&D, Minneapolis, MN), 0.1 mM nonessential amino acids, 1 mM sodium pyruvate, 2 mM L-glutamine, and 100 μM 2-mercaptoethanol (all from Sigma, St Louis, MO). Same culture conditions were used for the establishment and expansion of rPS cells.
Differentiation of iPS cells in vitro
EBs were generated as previously described [24]. Briefly, iPS cells grown on MEFs were split onto a gelatin-coated dish in Iscove's modified Dulbecco's medium (IMDM; Invitrogen) containing the same supplements as in the iPS cell growth medium and were cultured for 2 days. Single-cell suspensions were then prepared by treatment with 0.25% trypsin/ethylenediaminetetraacetic acid (Invitrogen). Cells were then seeded at a density of 104 cells/mL in differentiation medium (IMDM supplemented with 1% ascorbic acid, 2 mM L-glutamine, and 97.5 μg/mL monothioglycerol) on a bacteriological petri dish (Fisher Scientific, Pittsburgh, PA).
Differentiation of iPS cells in vivo
iPS cells were suspended at 2×107 cells/mL in DMEM containing 10% (v/v) FBS. A cell suspension of 100 μL was subcutaneously injected into the dorsal flank of a nude mouse. Six weeks after injection, tumors were surgically dissected, fixed in 4% (v/v) formaldehyde, and then embedded in paraffin. Sections were stained with hematoxylin and eosin. For frozen sections, tumors were fixed in 4% (v/v) formaldehyde, then cryoprotected in 30% sucrose for 3 days, and finally sectioned on a microtome.
Establishment of rPS cells from EBs and teratomas
To establish rPS cells from EBs, day 20 EBs were collected, dissociated by incubation with 0.3% collagenase (Sigma) for 20 min, and then plated on MEF-coated dishes with iPS cell culture medium. Cells were passaged as regular iPS cells after 4–5 days.
To establish rPS cells from teratomas, teratomas were harvested 6 weeks after injection. Single-cell suspensions were achieved by treatment with 0.3% collagenase for 60 min, and then cells were plated on MEF-coated dishes with iPS cell culture medium. Cells were passaged as regular iPS cells after 5–7 days.
Immunofluorescence staining
Immunofluorescence staining was carried out using standard procedures as previously described [25]. Goat anti-Mouse IgM-RPE was purchased from Southern Biotechnology Associates, Inc. (Birmingham, AL). Mouse anti-human SSEA-1 antibody was purchased from Santa Cruz Biotechnology Inc. (Santa Cruz, CA). Slides were observed with an Olympus IX50 fluorescence microscope system (Olympus, Tokyo, Japan) after staining.
Alkaline phosphatase and nuclei staining
Staining for alkaline phosphatase (AKP) was performed using a kit from shanghai rainbow medical reagent research company (Shanghai, China) following protocols provided by the manufacturer. Nuclei counterstaining was performed with 4′,6-diamidino-2-phenylindole (DAPI) (Biomol, 1:500 dilution).
Flow cytometry analysis
Flow cytometry analysis was carried out as previously described [25]. Briefly, EBs or teratomas were harvested and dissociated by enzymatic digestion to achieve a single-cell suspension. Cells were then labeled with anti-SSEA-1 antibody (Santa Cruz). All samples were analyzed on an Epics ALTRA flow cytometer (Beckman Coulter, Fullerton, CA), and the data were analyzed using CXP software (Beckman Coulter).
Reverse transcription–polymerase chain reaction analysis
Total RNA was extracted with TRIzol (Invitrogen) and reverse transcribed into cDNA with a reverse transcription–polymerase chain reaction (RT–PCR kit; TaKaRa, Shiga, Japan). The primer sequences (all from Sangon Technology Co., Shanghai, China), reaction conditions, and the sizes of each product are listed in Supplementary Table S1 (Supplementary Data are available online at
Results
Residual undifferentiated pluripotent stem cells in long-term differentiated EBs
iPS cells and EB differentiation
To test whether a small number of undifferentiated pluripotent stem cells reside in long-term differentiated EBs, 3 mouse iPS cell lines (IP14D-1, IP14D-2, and IP20D-3) were utilized. The iPS cells express an Oct-4-enhanced green fluorescent protein (GFP) reporter, in which the GFP expression is driven by the Oct-4 promoter. Since similar results were achieved from all 3 lines tested, data from IP20D-3 are presented in the whole article; whereas data from IP14D-2 are presented as supplementary files, and data from IP14D-1 are not shown. Parental iPS cells exhibited morphology similar to ES cells that packed tightly with smooth and refractile ring borders (Fig. 1A, Supplementary Fig. S1A). The majority of iPS cells (89.0%) expressed Oct-4-GFP (Fig. 1A, B and Supplementary Fig. S1A). RT–PCR analysis showed that Oct-4, Nanog, and Rex1 were all expressed in iPS cells (Fig. 3C). After 6 days of differentiation, Oct-4-GFP negative cells were observed in EBs (data not shown), indicating that some cells had undergone differentiation. RT–PCR analysis confirmed that FGF5 (ectodermal marker), Nestin (ectodermal marker), Brachyury (mesodermal marker), Flk-1 (mesodermal marker), and GATA-6 (endodermal marker) were all expressed in day 6 EBs (Fig. 3C). Interestingly, Oct-4-GFP positive cells could be still observed even after 20 days of EB differentiation (Fig. 1C, Supplementary Fig. S1B). Flow cytometry analysis showed that approximately 12.4% (Fig. 1D) of cells were still positive for GFP expression, indicating that undifferentiated stem cells might exist at this stage.
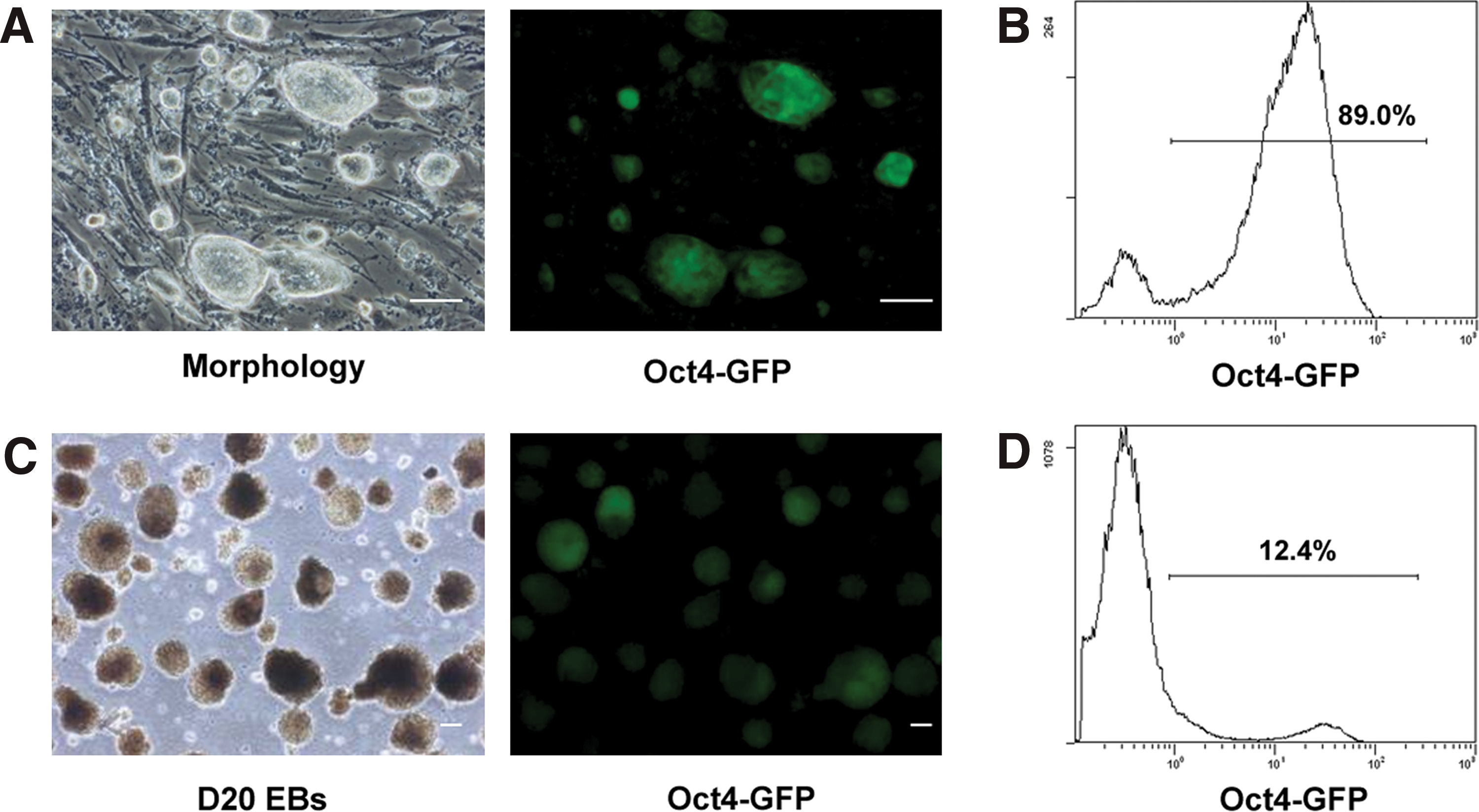
Differentiation of induced pluripotent stem cells in vitro.
Residual undifferentiated pluripotent stem cells derived from EBs
To detect whether long-term differentiated EBs contain undifferentiated stem cells, day 20 EBs were collected, dissociated, and then replated on MEF-coated dishes with iPS cell culture medium. Interestingly, iPS cell-like colonies were observed after 4–5 days of culture (Fig. 2A, Supplementary Fig. S1C). These cells could be continuously expanded under the same conditions without morphological changes for over 20 passages (data not shown). In addition, cells were positive for AKP staining (Fig. 2B, Supplementary Fig. S1C) and expressed Oct-4 and SSEA-1 (Fig. 2C, D and Supplementary Fig. S1C). RT–PCR analysis confirmed that Oct-4, Nanog, and Rex1 were all expressed in these cells (Fig. 3C). These were termed “rPS cells.” Further, rPS cells could also be easily isolated from day 40 EBs (data not shown). All these results demonstrated that residual undifferentiated cells do exist in long-term EBs, and this phenomenon was observed in all 3 types of mouse iPS cells tested.
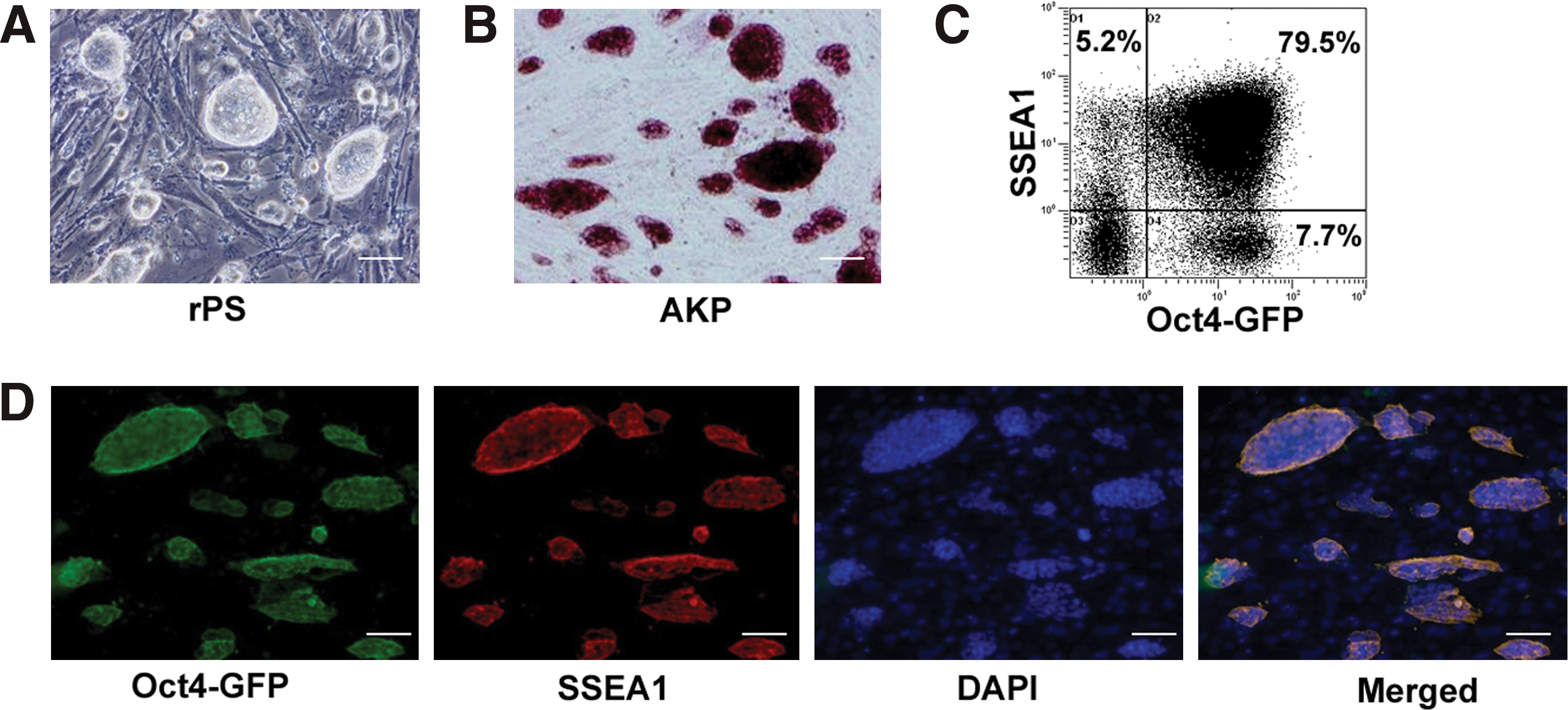
Residual pluripotent stem cells derived from long-term differentiation of mouse iPS cells in vitro.
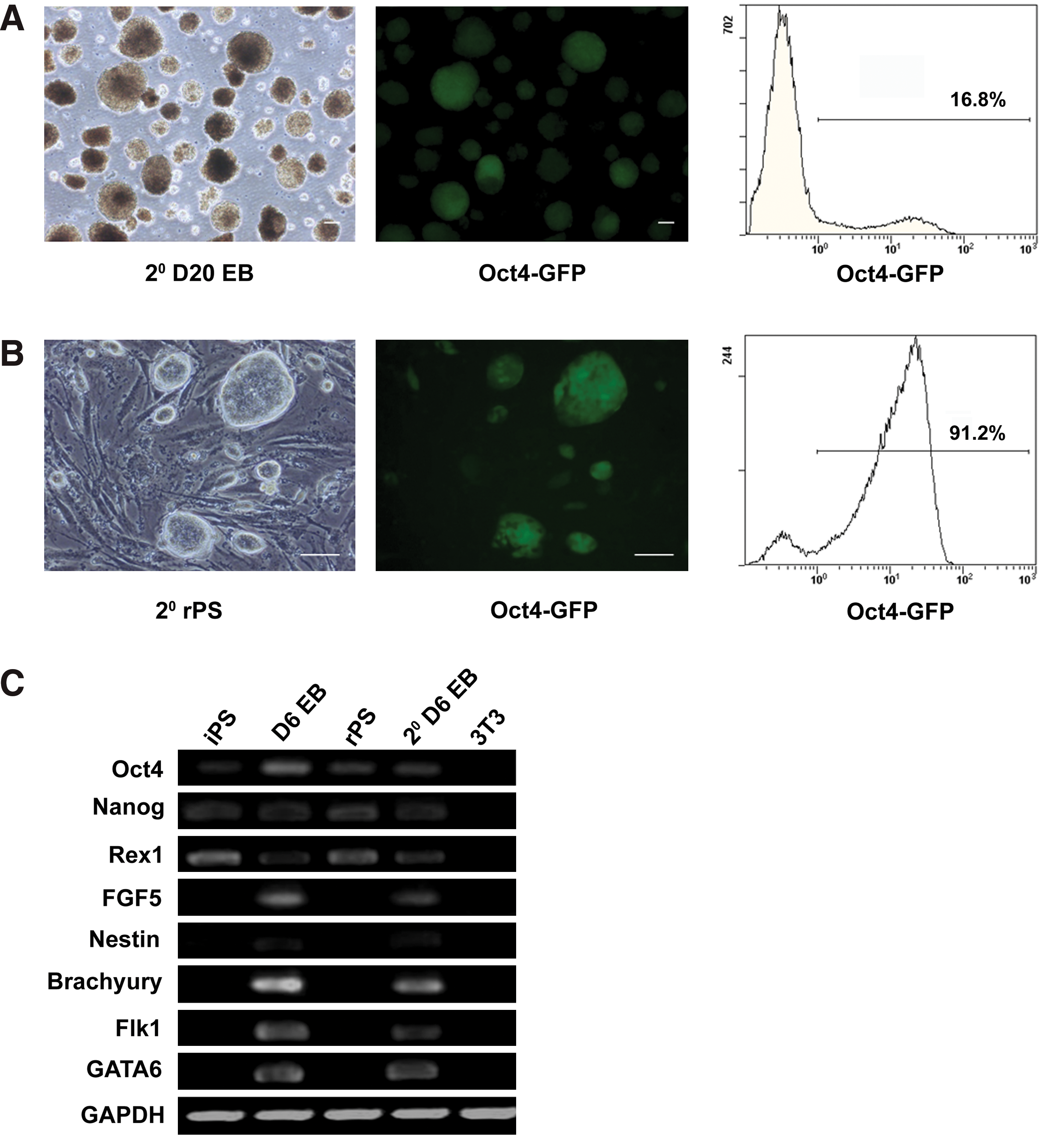
Redifferentiation of residual pluripotent stem cells and generation of secondary rPS cells.
Redifferentiation of EB-derived rPS cells in vitro and in vivo
To further test whether the EB-derived rPS cells could be redifferentiated and form secondary rPS cells, rPS cells at passage 5 were redifferentiated to form EBs. RT–PCR analyses showed that FGF5, Nestin, Brachyury, Flk-1, and GATA-6 were all expressed in secondary day 6 EBs (Fig. 3C). The expression of Oct-4/GFP again dropped to 16.8% in the secondary EBs at day 20 (Fig. 3A, Supplementary Fig. S2A). By replating the secondary EBs in iPS cell culture conditions, iPS cell-like colonies were observed. Again, these cells expressed Oct-4/GFP (Fig. 3B, Supplementary Fig. S2B), and they were positive for AKP staining (data not shown). Moreover, the secondary rPS cells could be redifferentiated and again formed rPS cells (data not shown). The differentiation potential of rPS cells was also evaluated by injecting them subcutaneously into nude mice. After 6–8 weeks, all mice (n=4) developed teratomas that contained tissues representative of all 3 germ layers including epithelial pearl, cartilage, and gut-like epithelium (Fig. 4B–D, Supplementary Fig. S2C). These results proved that residual undifferentiated cells were pluripotent and could redifferentiate in vitro and in vivo.

Residual pluripotent stem cells formed teratomas in vivo.
Residual undifferentiated pluripotent stem cells in teratomas
Deriving residual undifferentiated pluripotent stem cells from teratomas
To test whether the rPS phenomenon depends on the differentiation method, another cell differentiation system, the teratoma-formation system, was utilized. Parental iPS cells were first injected subcutaneously into nude mice. After 6–8 weeks, teratomas were formed and harvested. Flow cytometry analyses showed that about 0.6% of teratoma cells expressed Oct-4-GFP, with 0.3% of them co-expressing SSEA-1 (Fig. 5B, Supplementary Fig. S3A). The existence of Oct-4-GFP expressing cells was confirmed by frozen tissue sections and were found clustered together (Fig. 5C) or scattered in tissues (Fig. 5D). Pooled teratoma cells were then replated in iPS cell culture conditions. As expected, iPS-like colonies were observed after 5–7 days of culture and could be passaged similar to their parental cells. They showed similar characteristics to their parental cells, including morphology (Fig. 6A), AKP staining (Fig. 6B), and expression of SSEA-1 and Oct-4-GFP (Fig. 6D, Supplementary Fig. S3B). Flow cytometry analysis confirmed that 90.0% of cells expressed Oct-4, with 68.3% of them co-expressing GFP (Fig. 6C). Taken together, these results demonstrate that rPS cells also exist in teratomas, indicating that the residual phenomenon is not related to the differentiation methods.
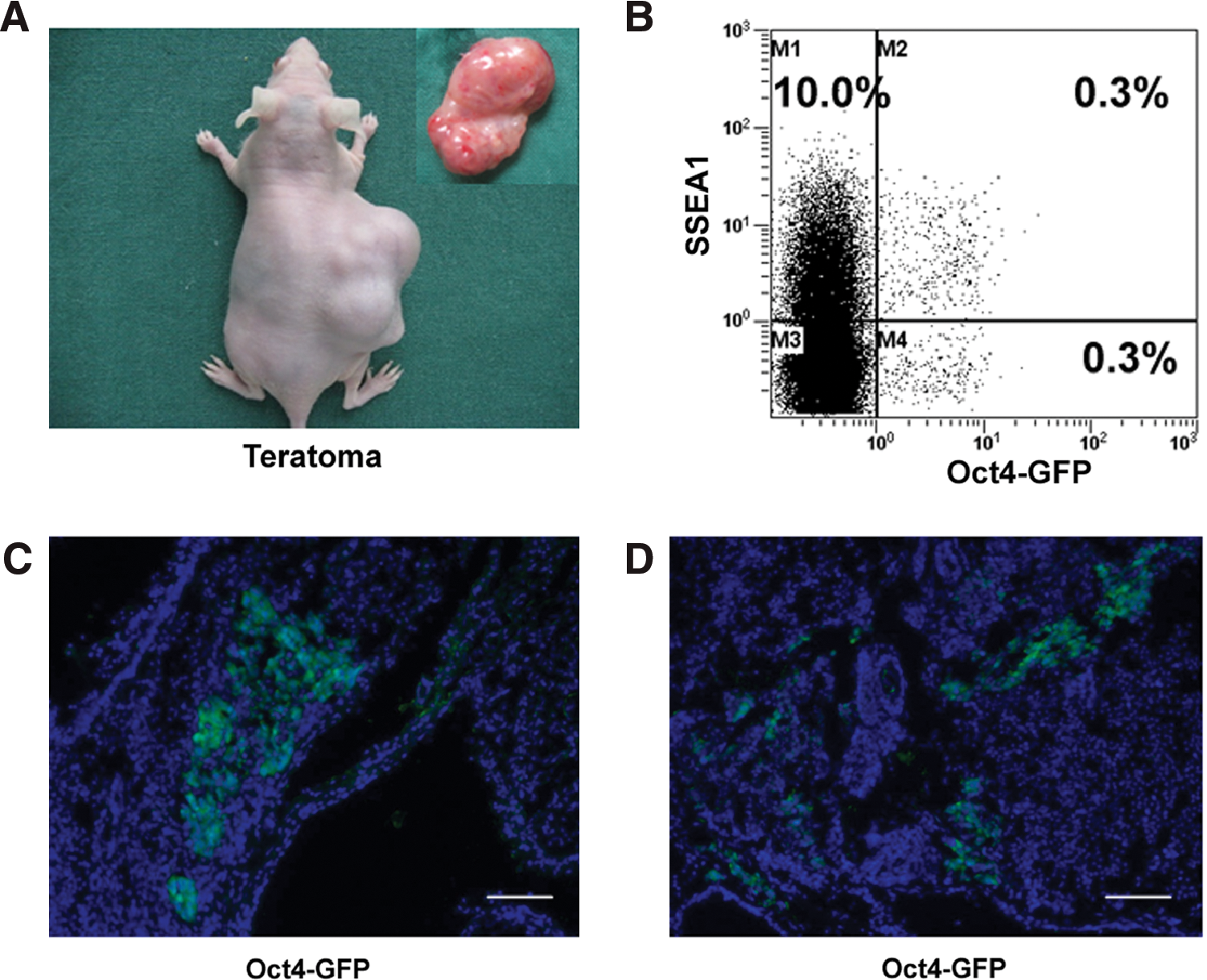
Differentiation of iPS cells in vivo.

Residual pluripotent stem cells derived from teratomas.
Redifferentiation of teratoma-derived rPS cells in vitro and in vivo
To further test whether the teratoma-derived rPS cells could be redifferentiated and form secondary rPS cells, cells were redifferentiated to form EBs. After 6 days of differentiation, Oct-4/GFP negative cells were observed under fluorescence microscopy (Fig. 7A). Flow cytometry analysis showed that only 28.8% of cells from day 6 EBs were Oct-4/GFP positive (Fig. 7A), indicating that the majority of rPS cells underwent differentiation in vitro. When rPS cells were injected into nude mice, teratomas again formed after 6–8 weeks (n=4) (Fig. 7B, Supplementary Fig. S3C). Not surprisingly, secondary rPS cells could be rederived from secondary teratomas in iPS cell culture conditions and showed similar characteristics to their parental cells (Fig. 7C, Supplementary Fig. S3D). These data suggest that teratoma-derived rPS cells were also pluripotent and could redifferentiate in vitro and in vivo.
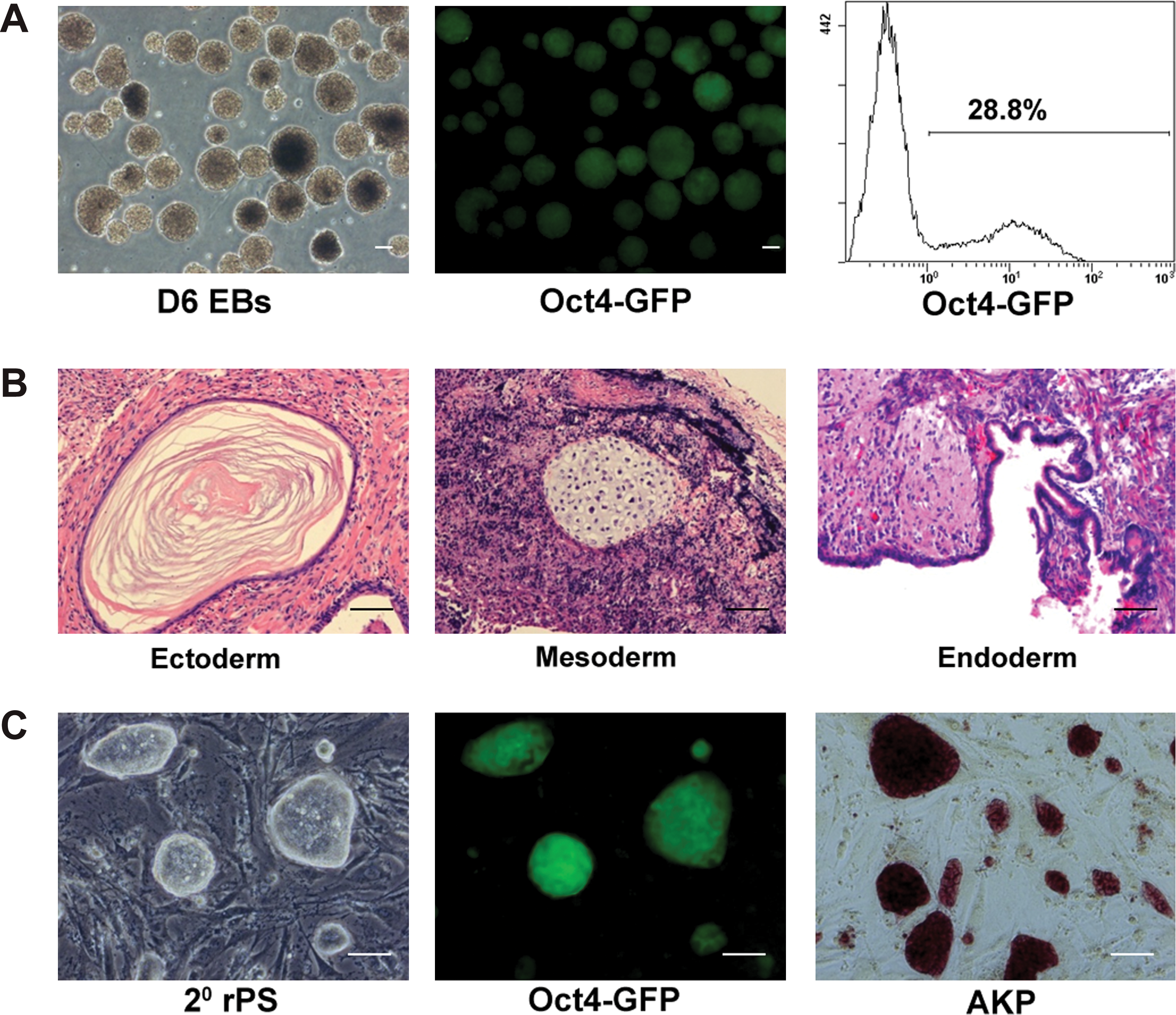
Pluripotency of rPS cells derived from teratomas.
Discussion
Pluripotent stem cells, including ES cells and iPS cells, are the potential sources for cell-based therapy due to their unlimited proliferation potential and pluripotent differentiation capacity. However, the presence of undifferentiated cells after induced or spontaneous differentiation could be tumorigenic during transplantation [26 –28]. Although the presence of pluripotent cells in spontaneous differentiation of ES and/or iPS cells might be due to the presence of cells of germ cell lineage [29,30], the current work demonstrates that the residual pluripotent cells are likely undifferentiated iPS cells rather than germ cells, as these cells were expanded under iPS cell culture conditions and were phenotypically similar to their parent cells.
Compared with ES cells, iPS cells obviously have many advantages for therapeutic use. However, as reviewed by several recent publications, iPS cells are more tumorigenic than ES cells due to their genetic and epigenetic causes [13,31]. On the one hand, the reprogramming process, which inevitably involves genetic manipulation, could introduce a greater risk for tumor formation. Studies have demonstrated that the tumorigenic potential of iPS cells depends on the origin of the cell types from which iPS cells are derived [28]. It is also affected by the methods used for reprogramming. Reprogramming methods that do not involve genomic integration have been reported to generate human iPS cells less susceptible to insertional mutagenesis. Thus, the tumorigenic risks related to the derivation of iPS cells could be reduced when the reprogramming techniques have been improved. On the other hand, iPS cells are derived from mature somatic cells that have undergone multiple cell divisions and may have already acquired genetic mutations. Such genetic changes could not be eliminated no matter what kind of reprogramming methods are utilized [32].
Putting aside the tumorigenic reasons just mentioned, the current work attempted to deduce whether iPS cells with normal developmental potential are still tumorigenic. Three iPS cell lines, including the one that is able to generate full-term mice via tetraploid blastocyst complementation, were tested. We found that undifferentiated iPS cells could be steadily isolated and expanded from long-term (day 20 and 40) EBs as well as from teratomas (Fig. 2, Supplementary Fig. S1) in all cell types tested. In addition, these undifferentiated iPS cells could be expanded and redifferentiated in vitro and in vivo, and undifferentiated pluripotent stem cells could be isolated after rounds of differentiation. The undifferentiated iPS cells could not be simply eliminated by extended cell differentiation, suggesting that the residual cells could be a potential tumorigenic risk during transplantation. However, both in EB and teratoma-formation processes, cells are spontaneously differentiated. Interestingly, Miura et al. also observed that a small number of Nanog positive cells existed during induced differentiation of iPS cells [28]. Based on those data, it is likely that keeping residual cells at undifferentiated state does not depend on the differentiation method utilized. Therefore, induced differentiation might decrease the percentage of residual cells, but probably cannot completely eliminate the residual cells.
Our previous work proved that residual pluripotent cells also existed during mouse ES cells differentiation [25]. The residual phenomenon in mouse ES was also observed by Enseñat-Waser et al. [33]. However, they found karyotypic abnormality in the residual cells, which consequently inhibit the cell differentiation, resulting in the maintenance of theses cells at undifferentiated state. On the contrary, we found that the residual ES cells were nonmutant cells that were able to redifferentiate and form chimeric mice by blastocyst injection (unpublished data). In the present study, we found that residual iPS cells were able to redifferentiate in vitro and in vivo (Figs. 3, 4), indicating they are likely normal cells. Further analyses of karyotype and their developmental potential should be performed. The mechanism of how these cells remain undifferentiated during differentiation remains unclear. Since the “residual” phenomenon also exists in mouse ES cell differentiation, we speculate that this might be a common characteristic of pluripotent cells no matter whether they are derived from embryos or by reprogramming. Another explanation could be that introduced genes for reprogramming (Nanog, Oct-4, Klf4, and c-Myc) might reactivate during differentiation, although this is not likely the case. Checking the expression of exogenous Nanog, Oct-4, Klf4, and c-Myc in the residual cells could directly answer this question.
Since residual iPS cells could not be eliminated by extended cell differentiation, strategies of removing these residual cells after differentiation should be considered. These strategies include targeted elimination of pluripotent cells by cytotoxic antibodies [34,35], the separation of undifferentiated cells on the basis of pluripotent-specific cell surface molecules [36], or introduction of “suicide” genes into cells that could be activated by specific drugs if teratomas were formed after transplantation [37]. Alternatively, it would be more attractive to reprogram somatic cells to a multipotent or a unipotent state without residual potential to avoid tumorigenic problems [13,38]. Szabo et al. converted human fibroblasts into multipotent hematopoietic progenitors of the myeloid, erythroid, and megakaryocytic lineages without traversing the pluripotent state or activation of mesodermal pathways [39]. Vierbuchen et al. also showed that mouse fibroblasts could be efficiently converted into functional neurons [40], and Ieda et al. demonstrated the induction of cardiomyocyte-like cells from postnatal cardiac and dermal fibroblasts [41]. Meanwhile, Feng et al. showed that NIH 3T3 cells as well as primary mouse embryo- and skin-derived fibroblasts can be converted into macrophage-like cells by retroviral expression of PU.1 and C/EBPα [42]. Those multipotent, unipotent and mature cells have great therapeutic implications, and those elegant studies open up the possibility of alternate methods of deriving safe patient-specific lineages for clinical application, hopefully without teratoma formation.
In conclusion, this is the first study to demonstrate that in iPS cells with normal development potential, a part of cells could be maintained in a pluripotent state during differentiation, which could be a tumorigenic risk in further transplantation. These findings help us better understand the nature of iPS cells. In addition, it prompts us to reconsider the strategies for solving the tumorigenic problem of iPS cells, not only focusing on improving the reprogramming process. Reprogramming somatic cells to a multipotent or unipotent state rather than a pluripotent state might be a more sensible strategy for clinical application.
Footnotes
Acknowledgment
This work was supported by the Major State Basic Research Development Program of China (2005CB522700, 2007CB948004, 2011CB964704), Shanghai Pu Jiang foundation (to Zhang WJ), Shanghai Shu Guang foundation (to Zhang WJ), Shanghai-Northeast England Science and Technology Cooperation Program (075407072), Shanghai Jiao Tong University School of Medicine Training Programme for Outstanding Doctoral Theses (to Fu Wei), and Shanghai Jiao Tong University School of Medicine Doctoral Innovation foundation (BXJ0920). The authors also appreciate the technical support from Deli Liu, Demin Ying, Lijuan Zong, and Juanjuan Wu.
Author Disclosure Statement
None of the authors have any disclosures to declare. No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.