
Abstract
Molecular and cellular signaling pathways are involved in the process of neural differentiation from human embryonic stem cells (hESC) to terminally differentiated neurons. The Sonic hedgehog (SHH) morphogen is required to direct the differentiation of hESC to several neural subtypes, for example, dopaminergic (DA) or motor neurons. However, the roles of SHH signaling and the pathway target genes that regulate the diversity of cellular responses arising from SHH activation during neurogenesis of hESC have yet to be elucidated. In this study, we report that overexpression of SHH in hESC promotes the derivation of neuroprogenitors (NP), increases proliferation of NP, and subsequently increases the yield of DA neurons. Next, gene expression changes resulting from the overexpression of SHH in hESC-derived NP were examined by genome-wide transcriptional profiling. Categorizing the differentially expressed genes according to the Gene Ontology biological processes showed that they are involved in numerous cellular processes, including neural development, NP proliferation, and neural specification. In silico GLI-binding sites analysis of the differentially expressed genes also identified a set of putative novel direct target genes of SHH in hESC-derived NP, which are involved in nervous system development. Electrophoretic mobility shift assays and promoter-luciferase assays confirmed that GLI1 binds to the promoter region and activates transcription of HEY2, a NOTCH signaling target gene. Taken together, our data provide evidence for the first time that there is cross-talk between the NOTCH and SHH signaling pathways in hESC-derived NP and also provide significant new insights into transcriptional targets in SHH-mediated neural differentiation of hESC.
Introduction
T
hESC are pluripotent cells derived from the preimplantation blastocyst and are able to differentiate into all cell types of the body [4]. To date, hESC have been successfully differentiated into a great variety of cells that make up the central nervous system, including dopaminergic (DA) neurons, motor neurons, glial cells, astrocytes, oligodendrocytes, and neural crest stem cells. The directed differentiation of hESC toward the neuroectoderm lineage can be achieved by coculture of hESC with stromal cells like PA6 and MS5 [5 –8]. Alternatively, hESC can be treated with neural inductive factors like fibroblast growth factor 2 (FGF2) [9 –11] and Noggin [12,13] to produce neuroprogenitors (NP) that can then be further differentiated to specific neuronal subtypes with regional identity. SHH is one of the factors added at this stage that is required to efficiently obtain neurons of ventral identity, for example, motor neurons and DA neurons [7 –9,14 –16]. Apart from its role in ventralizing NP, there is evidence in mouse ESC (mESC) that suggests that SHH signaling is required for neuroectoderm development and survival of NP [17,18].
We recently showed that the SHH signaling pathway in hESC is poised for activation upon differentiation and influences the determination of early differentiated hESC toward the neuroectoderm lineage [19]. This prompted us to study the role of SHH in hESC neural differentiation. Here we identify the cellular responses and the direct SHH target genes arising from SHH activation in hESC-derived NP and provide a new paradigm for understanding the molecular pathway of hESC neural differentiation.
Materials and Methods
Cell culture and differentiation
hESC line HES-3 (46 XX) was from ES Cell International, whereas the human induced pluripotent stem cell (iPSC) line (IMR90) was generously provided by JA Thomson from University of Wisconsin–Madison [20]. 293FT cells were from Invitrogen. Cells were cultured as previously described [21,22]. To initiate differentiation, undifferentiated hESC were broken into clumps and grown as embryoid bodies (EB) for 4 days as previously described [21]. EB were then transferred onto laminin-coated (10 μg/mL; Sigma) 6-well plates. Cells were grown in N2B27 media containing DMEM/F12, 1×N2, 1×B27 without vitamin A (all from Invitrogen), 1% NEAA, 1 mM L-glutamine, 25 U/mL penicillin–streptomycin, and 0.1 mM β-mercaptoethanol. N2B27 medium was supplemented with 500 ng/mL Noggin (R&D Systems). After 10 days, compact clumps were formed and were cut up manually with a pipette and grown as neurospheres in suspension in N2B27 media supplemented with 20 ng/mL EGF (Peprotech) and 20 ng/mL FGF2. The neurospheres were passaged every 6–7 days using TrypLE Express (Invitrogen).
To induce DA neurons differentiation, neurospheres were dissociated into small clumps and plated onto laminin-coated 24-well plates and fed with 200 ng/mL FGF8, 200 ng/mL SHH (both from R&D Systems), and 200 μM ascorbic acid (AA; Sigma). After 7–10 days, reformed rosettes were selectively excised and replated onto laminin-coated coverslips at ∼50,000–150,000 cells per cm2 in N2B27 media. The following growth factors were added: 1 μM cyclic adenosine monophosphate (Sigma), 20 ng/mL brain-derived neurotrophic factor (Peprotech), and 20 ng/mL glial cell line-derived neurotrophic factor (Peprotech), 200 ng/mL FGF8, and 200 ng/mL SHH. The cells were fed every 3–4 days and allowed to differentiate for up to 14 days (Supplementary Fig. S1; Supplementary Data are available online at
Generation of stable SHH-overexpressing hESC lines
hESC lines (HES-3) overexpressing SHH (CHEF-SHH-IRES-DsRed2) and Vector control (CHEF-DsRed2) constructs were generated as previously described [23]. CHEF-SHH-IRES-DsRed2 construct encodes the mouse full-length Shh linked through an IRES (internal ribosome entry site) to red fluorescent protein (DsRed2) driven by CHEF1 promoter.
RNA extraction and real-time PCR analysis
RNA samples were extracted from cells using QIAGEN RNAeasy kit according to manufacturer's instruction. Reverse transcription into cDNA was preformed using Superscript First Strand Synthesis System (Invitrogen) according to manufacturer's protocol. Quantitative real-time PCR was performed as previously described [21]. Primers used for amplification are listed in Supplementary Table S1.
Transcriptional profiling
Genome-wide gene expression profiling was carried out for biological quadruplicates of NP derived from SHH-overexpressing hESC (SHH-NP), from control vector expressing hESC (Vector-NP) and nontransfected hESC (H3-NP), using the Affymetrix GeneChip Human Genome array (HG-U133 Plus 2.0). The details are described in Supplementary Material and Methods. Chips were scanned and normalized expression signals were computed using RMA [24]. Differential hybridized features were identified using Limma [25]. In Limma, P values were compiled from 4 sets of technical replicate data and obtained from moderated t statistics or F statistics using empirical Bayesian methods. P values were then adjusted for multiple testing with Benjamini and Hochberg's method to control the false discovery rate [26]. All differentially expressed genes were filtered using the statistic criteria of adjusted P values <0.05 and a 1.5-fold change in expression ratio compared with both Vector-NP and H3-NP. Statistically significant over- or under-representation of particular gene ontology (GO) term pathways were identified with either a classical hypergeometric test or a conditional hypergeometric that uses the relationships among GO terms to decorrelate the results. All computations were done using the statistical programming language R in combination with Bioconductor tools.
In silico analysis of GLI-binding sites
The regulatory sequences 5 kb upstream and downstream of the 1.5-fold differentially expressed genes were extracted from the Biomart database (
Flow cytometry, immunocytochemistry, and western blot analysis
Flow cytometry, immunocytochemical staining, and western blot analysis were performed as previously described [19,23]. For immunocytochemical staining, images were acquired using a Zeiss Axiovert 200M inverted fluorescent microscope and Zeiss LSM 510 Meta Confocal Microscope (Carl Zeiss;
Electrophysiology recording
Spontaneous postsynaptic currents and membrane potential were monitored by standard patch clamp protocol. In brief, a neuron intended for postsynaptic recordings was patched with pipettes that were pulled from borosilicate glass capillary tubes using a pipette puller. The whole-cell pipette solution contained (in mM) 135 CsCl, 10 HEPES, 1 EGTA, 1 Na-GTP, 4 Mg-ATP, and 10 QX-314 (pH 7.4, adjusted with CsOH). The resistance of pipettes filled with the intracellular solution ranged between 4 and 5 MΩ. After formation of the whole-cell configuration and equilibration of the intracellular pipette solution, the series resistance was adjusted to 10 MΩ. Cells whose initial Rs exceeded 15 MΩ were excluded from analysis. Synaptic currents were monitored with EPC 10 (HEKA). The bath solution contained (in mM) 140 NaCl, 5 KCl, 2 CaCl2 or 2 MgCl2, 10 HEPES, and 10 glucose (pH 7.4, adjusted with NaOH). Spontaneous postsynaptic currents and membrane potential were monitored. The temperature in the recording chamber was controlled by TC344B dual temperature controller (Warner Instruments).
Cell count assay
About 1×105 p75+/PSA-NCAM+ sorted cells of SHH-NP were seeded into 24-well ultra-low suspension plates and neurospheres formed after 3–5 days. p75+/PSA-NCAM+ positive neurospheres were harvested 7 and 14 days after isolation and treated with TrypLE Express to obtain a single cell suspension. The viable cell number for each sample was determined using Trypan Blue exclusion with a hemocytometer (Neubauer).
Statistical analysis
Statistical significance of differences between values was evaluated by an unpaired Student's t-test in Microsoft Excel. Significance was set at a P value of <0.05. Analysis of variance (ANOVA) and Newman–Keuls post hoc analysis (STATISTICA 10; StatSoft) were used for 3 group comparisons. Data are presented as mean±SEM unless indicated otherwise.
Cell proliferation assay
Cell proliferation assay was performed using the Click-iT EdU (thymidine analog 5-ethynyl-2′-deoxyuridine) flow cytometry assay kit (Invitrogen) and according to manufacturer's protocol. Briefly, EdU was added into the culture media at a final concentration of 10 μM for 2 h to allow the incorporation of EdU into DNA. Cells were then harvested, fixed, and permeabilized followed by the incubation with reaction cocktails for 30 min in the dark. EdU incorporation was detected using a FACS Calibur (Becton Dickinson) and results were analyzed with the CellQuest Software (Becton Dickinson).
Electrophoretic mobility shift assays
Electrophoretic mobility shift assays (EMSA) was performed as previously described [21]. Nuclear extracts of 293FT cells overexpressing GLI1 were used for gel shift assays. For supershift assay, hESC-derived NP nuclear extracts were incubated with rabbit anti-GLI1 antibody (Abcam). The details are described in Supplementary Material and Methods. Probe sequences are listed in Supplementary Table S3.
Luciferase reporter assay
The human HEY2 promoter-luciferase reporter was obtained from Switchgear (Menlo Park). Mutations were made on the putative GLI1 binding sites in the HEY2 promoter of the reporter plasmid using the QuikChange Multi Site-Directed Mutagenesis kit (Stratagene) and sequence-verified. The wild-type or mutant HEY2 promoter luciferase plasmids were cotransfected with pRL-TK (Promega) internal control plasmid and the SHH expression vector (Open Biosystems) as indicated. A dual luciferase system (Promega) was used as previously described [19].
Results and Discussion
Generation of SHH-overexpressing hESC and neural differentiation characterization
To establish the role of SHH in hESC neural differentiation, a stable SHH-overexpressing hESC line was first generated. The overexpression of SHH was done in undifferentiated hESC so that exogenous SHH could be present throughout the differentiation process. It would also overcome the limitation of diffusion of SHH into the inner layers of cells grown as clumps or spheres in suspension [27]. Undifferentiated hESC were transfected with the pCHEF-SHH-IRES-DsRed2 plasmid that encodes Shh with high-expression CHEF1 promoter system [23], or the control empty vector pCHEF-DsRed2. Two SHH-overexpressing stable cell lines were obtained after geneticin antibiotic selection and only one of these lines was characterized more extensively in further experiments. Immunofluorescence staining of a typical colony of the SHH-overexpressing hESC showed that the cells were SHH positive and coexpressed DsRed (Supplementary Fig. S2). The SHH-overexpressing hESC could be maintained in the undifferentiated state for over 10 passages and was karyotypically stable (data not shown). The maintenance of pluripotency in the SHH-overexpressing hESC was congruent with the observation in our previous study that exogenous SHH did not speed up differentiation of hESC [19].
Next, a neural differentiation protocol modified from reported methods in Zhang et al. [11] and Pera et al. [28] was developed so that the SHH-overexpressing hESC could be differentiated in a controlled and reproducible fashion (Supplementary Fig. S1). hESC differentiated EB were plated in serum-free media supplemented with the BMP inhibitor Noggin for 10 days, after which the clumps were PAX6 positive and displayed a compact and tight morphology that contained small rosettes at the edges (Supplementary Fig. S3A, B). The clumps with rosettes were dissected from surrounding fibroblastic cells and grown in suspension as neurospheres that were Nestin positive (Supplementary Fig. S3C, D). Neurospheres contained NP and were passaged for <5 passages to avoid any changes in differentiation potential [13]. The expression of neuroectoderm markers in the NP was examined by quantitative real-time PCR, and transcripts of SOX1, MSI, NESTIN, and PAX6 were shown to be upregulated by >2.5-fold when compared with undifferentiated hESC (Supplementary Fig. S3E). Expression of the pluripotent marker OCT4 was detected at very low levels in the NP (Supplementary Fig. S3E). The NP also had low expression levels of mesodermal and endodermal markers when compared with spontaneously differentiated EB (Supplementary Fig. S3F). This confirmed that the NP did not contain cells from other lineages. Flow cytometry analysis showed that NP expressed a comprehensive set of neural stem cell (NSC) surface markers [29]: A2B5 (34%±18%), FORSE-1 (51%±20%), p75 (83%±12%), PSA-NCAM (94%±3%), and CD133 (34%±11%) (Supplementary Fig. S3G). To further characterize the NP within the neurospheres, neurospheres were dissociated into smaller clumps and replated on laminin-coated plates. After 4–5 days in culture, rosettes were abundantly formed in the culture (Fig. 1A–C), demonstrating that neural induction was achieved using the protocol.

Neural differentiation of hESC.
To confirm that the NP are multipotent, NP were further differentiated by replating them as monolayer cultures without the mitogens epidermal growth factor (EGF) and FGF2. After 2 weeks, neurons (MAP2+/TH+) and astrocytes (GFAP+) were observed (Fig. 1D, E). However, oligodendrocytes were rarely obtained. A similar observation has been reported [30], suggesting that oligodendrocytic differentiation requires a more concerted effort which requires other growth factors and cell substrates [31]. Electrophysiological techniques (n=10 cells) were carried out to assess functional maturation of neurons differentiated from the NP. The whole-cell patch clamp experiment detected spontaneous postsynaptic currents and membrane potential form the neurons (Fig. 1F, G), confirming that the neural differentiation protocol used efficiently generates NP from hESC, which can be further differentiated into functional neurons and astrocytes. This optimized differentiation scheme using Noggin as neural inducer is an improvement on the currently published protocols. First, it eliminates the use of mouse feeder layer or stromal cells [7,28]; second, it employs a serum-free, chemically defined media, both of which reduce biological variations.
Overexpression of SHH enhances neural differentiation
To determine if overexpression of SHH would improve the efficiency of neural differentiation, the SHH-overexpressing hESC were differentiated into NP (SHH-NP) with sustained expression of the DsRed fluorescent protein and active N-terminal SHH proteins (Fig. 2A, B) and were characterized with SHH target genes and NSC markers. Quantification of SHH levels by ELISA demonstrated that the supernatant of SHH-NP cultures contained ∼20–50 ng/mL of SHH proteins (data not shown). Highly upregulated SHH (20-fold), PTCH1 (7-fold), and GLI1 (4-fold) mRNA expression was detected by real-time PCR, suggesting the activation of the SHH pathway (Fig. 2C). SHH-NP were also analyzed by real-time PCR for neuroectoderm markers. SHH-NP had a 2-fold increase in NESTIN and 5-fold increase in SOX1 expression in SHH-NP as compared with control NP differentiated from nontransfected hESC (H3-NP) and from control vector transfected hESC (Vector-NP) (Fig. 3A). Western blot analysis corroborated with the gene expression studies as demonstrated by the increase in NESTIN and SOX1 protein expression in SHH-NP (Fig. 3B). There was also an increase in expression of NSC surface markers in SHH-NP compared with average of control H3-NP and Vector-NP, respectively, A2B5 (70% vs. 40% positive), p75 (80% vs. 60% positive), CD133 (40% vs. 30% positive), and PSA-NCAM (90% vs. 80% positive) (Fig. 3C). All these surface markers have been demonstrated to be indicative of highly proliferative and multipotent NP derived from hESC [29,30,32]. Hence, the enrichment of cells expressing both extracellular and intracellular NSC markers in SHH-NP demonstrate that overexpression of SHH promotes the derivation of NP from hESC. This is consistent with our earlier study that long-term exposure of SHH promotes neuroectoderm differentiation of hESC-derived EB [19]. Moreover, it has also been showed that neural rosette cells could be maintained long-term in high-density cultures by activating the SHH and Notch pathways [10].
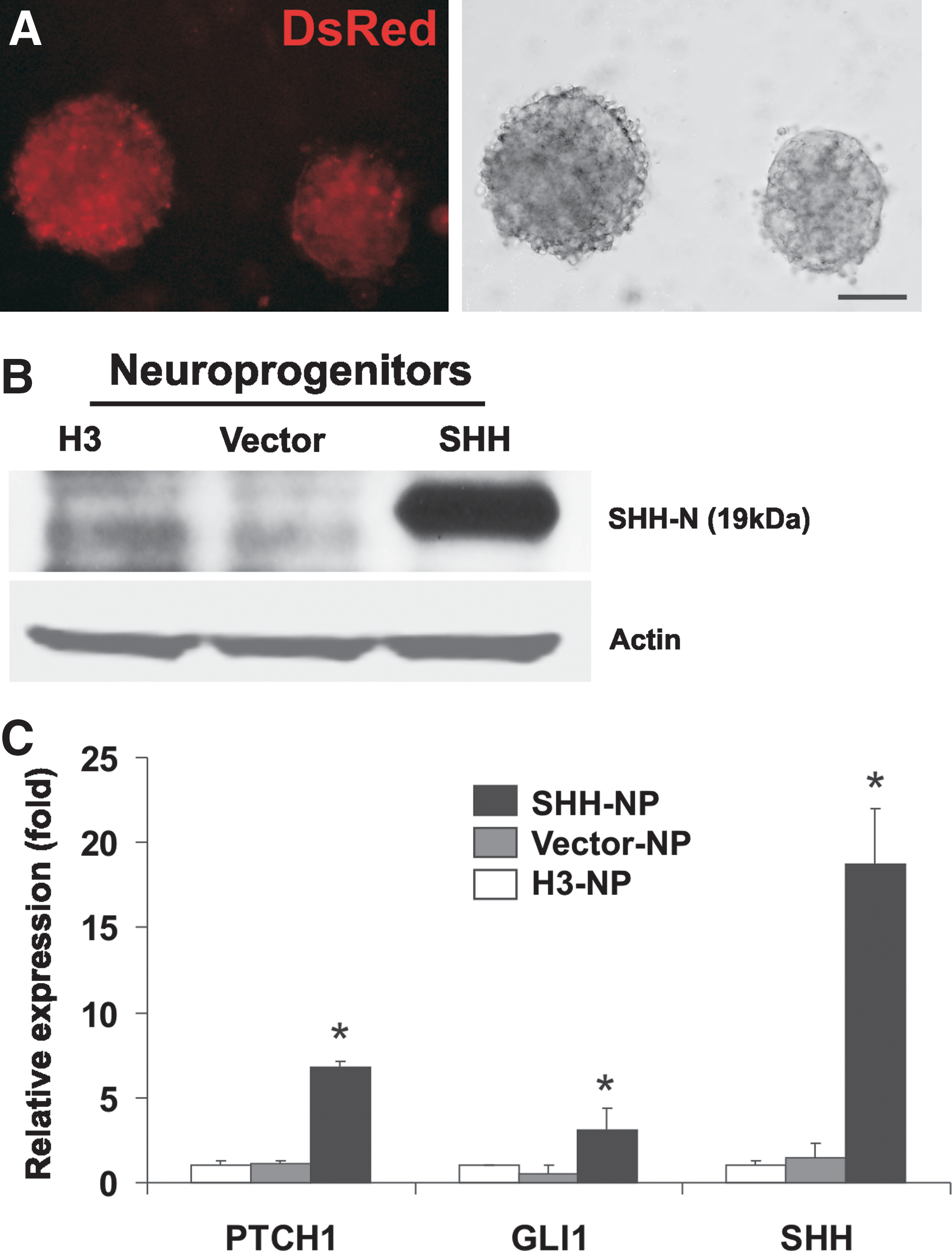
Generation of SHH-overexpressing hESC-derived NP.
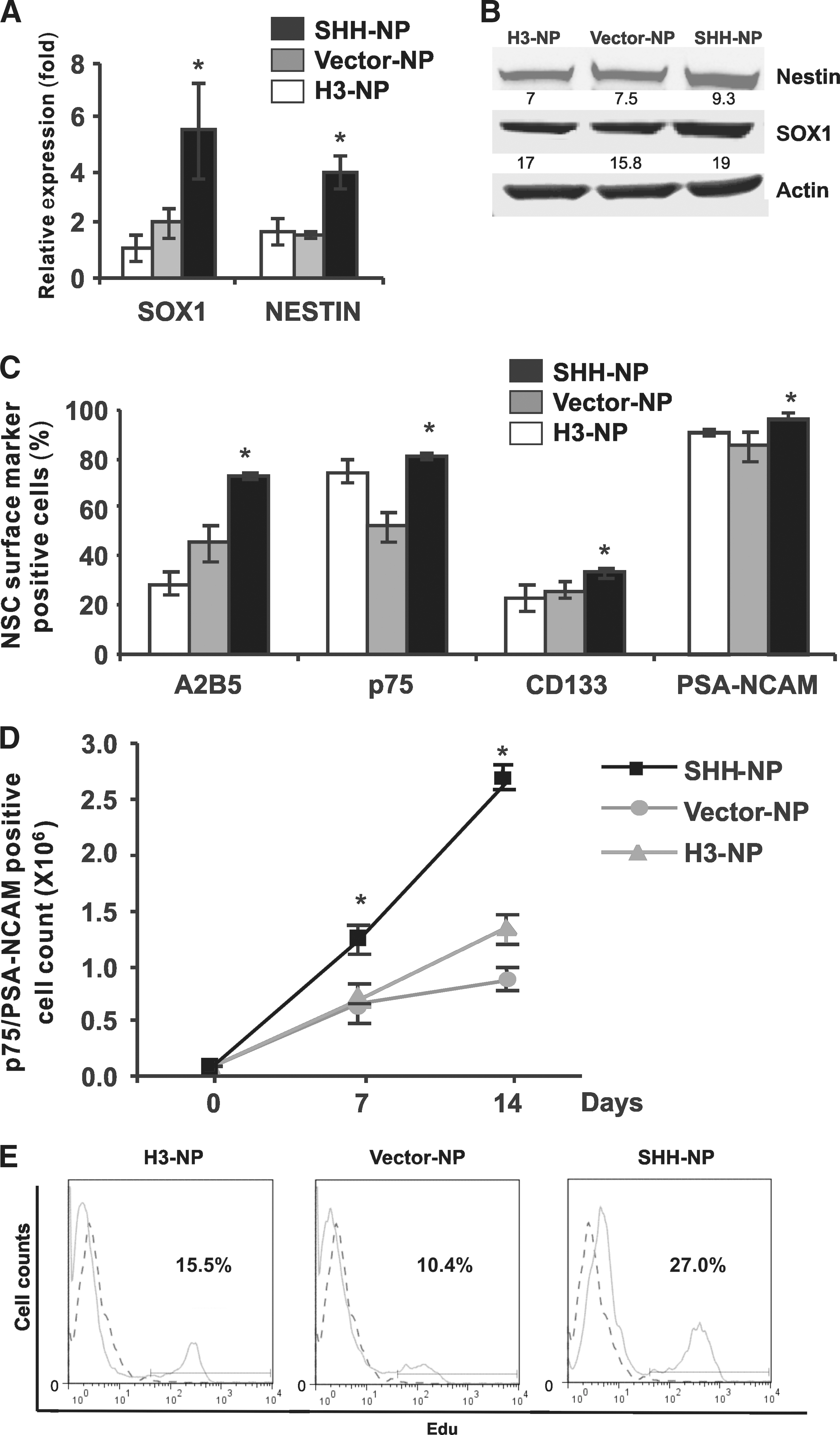
Overexpression of SHH in hESC-derived NP leads to an increase in expression of NSC markers and increase in proliferation.
Overexpression of SHH increases the proliferation of NSC markers sorted NP
As overexpression of SHH enhances neural derivation, there was interest to study if there was an increase in proliferation of NP with overexpression of SHH. Flow cytometry cell sorting based on the dual positive (+) expression of p75 and PSA-NCAM was carried out. Seven days after cell sorting, there was around a 2-fold increase of in the number of p75+/PSA-NCAM+ cells from the SHH-NP compared with control H3-NP and Vector-NP (Fig. 3D). A higher increase in cell number (2.5-fold) and in EdU stained-positive cells (about 12% more) of sorted p75+/PSA-NCAM+ cells from the SHH-NP was also observed even after 14 days culture (Fig. 3D, E). Hence, the results suggested that overexpression of SHH promoted the cell proliferation of p75+/PSANCAM+ NP. Further studies will be required to understand if the increase in cell number was through a change in survival, apoptosis, or both. During mouse development, Shh has been shown to regulate cell proliferation and survival in early neuroepithelial precursors, before 9.0 days post coitum (dpc) [33]. SHH is also known to have proliferative effects on cerebellar granule cell precursors, neuroepithelial cells, and precursors of the developing and adult forebrain [34 –36]. In addition, it has been suggested that during neurogenesis of mESC, Shh may not play an important role in neural determination, but rather improves survival and proliferation of Sox1-positive cells [17]. Taken together, our results here suggest that overexpression of SHH increases the expansion of hESC-derived NP.
Overexpression of SHH leads to increase in DA neurons
The enrichment of NP-expressing NSC markers within SHH-NP offered the prospect that there may be downstream effects of an increase in differentiated progeny. In this case, the DA neuronal lineage was investigated for NP differentiation efficiency because Noggin has been shown to improve DA neuron differentiation from hESC differentiated on PA6 cells [37]. In addition, it has been demonstrated that efficient derivation of DA neurons from NP required the presence of SHH. To do this, SHH-NP, control H3-NP, and Vector-NP were differentiated into DA neurons with FGF8 and AA and further with different growth factors. Exogenous SHH was added into control H3-NP and Vector-NP only. During SHH-NP differentiation into neurons, long neurite extensions were evident within 2–3 days and subsequently complex neurite networks were formed. After 14 days of differentiation, observation by immunofluorescence staining showed that there were more TH+ and β-tubulin III-positive (β-tubulin III+) neurons derived from SHH-NP as compared with control neurons from H3-NP and Vector-NP (Fig. 4A–C). Quantification of DA neurons was carried out, and of the β-tubulin III+ neurons derived from SHH-NP, significantly 50% were TH+ (Fig. 4D). In contrast, only ∼30% of the β-tubulin III+ neurons were TH+ in control neurons derived from H3-NP and Vector-NP. This represented a 20% increase in yield of TH+ DA neurons. PTCH1 and SHH mRNA expression was upregulated in SHH-NP-derived DA neurons, affirming that overexpression of SHH and activation of the pathway was sustained throughout the differentiation process. In addition, there was an ∼1.5-fold increase in mRNA expression of the dopamine enzymes AADC and TH in SHH-NP derived DA neurons (SHH-DN) as compared with control DA neurons from H3-NP (H3-DN) and Vector-NP (Vector-DN) (Fig. 4E). There was also a 6-fold increase in the midbrain DA marker Engrailed-1(EN1) in SHH-DN (Fig. 4E). Therefore, the results showed that overexpression of SHH led to the increased production of TH+DA neurons that expressed appropriate gene markers of DA neurons. We postulate that the increase in TH+ DA neurons derived from SHH-NP was due to the higher starting number of NSC in SHH-NP that were able to eventually differentiate successfully into DA neurons. Signaling factors (eg, SHH and FGF8) have been shown to enhance specification of midbrain DA neurons during embryogenesis, and they also seem to influence neuronal fate during hESC neural differentiation. It has been suggested that the functional outcome of SHH signaling might be dependent upon the stage of neural differentiation [38]. Collectively, as compared with other differentiation processes using SHH at the later stages of neural patterning and differentiation, our data indicate that overexpression of SHH from the undifferentiated stage through the entire process of neural induction and differentiation could lead to an overall increase in the yield of downstream neuronal subtypes.

Overexpression of SHH in hESC-derived NP leads to an increase in TH+ DA neurons.
Identification of direct SHH target genes in hESC-derived NP
Given the importance of SHH in promoting hESC neural differentiation, genome-wide transcriptome profiling analysis of SHH-overexpressing NP was carried out to understand the molecular targets and pathways downstream of SHH. A comparative gene expression analysis of SHH-NP, H3-NP, and Vector-NP was performed using the Affymetrix Human Genome U133 array, which analyzed the expression of 38,500 well-characterized human genes. For identification of relevant differentially expressed genes (DEG), only genes with more than 1.5-fold difference in expression were considered for further evaluation. On top of that, the genes had to be differentially expressed in both SHH-NP versus H3-NP and SHH-NP versus Vector-NP data sets. A total of 337 annotated genes were identified by the array: 182 were upregulated and 155 were downregulated in SHH-NP when compared with H3-NP and Vector-NP (Fig. 5A, B).
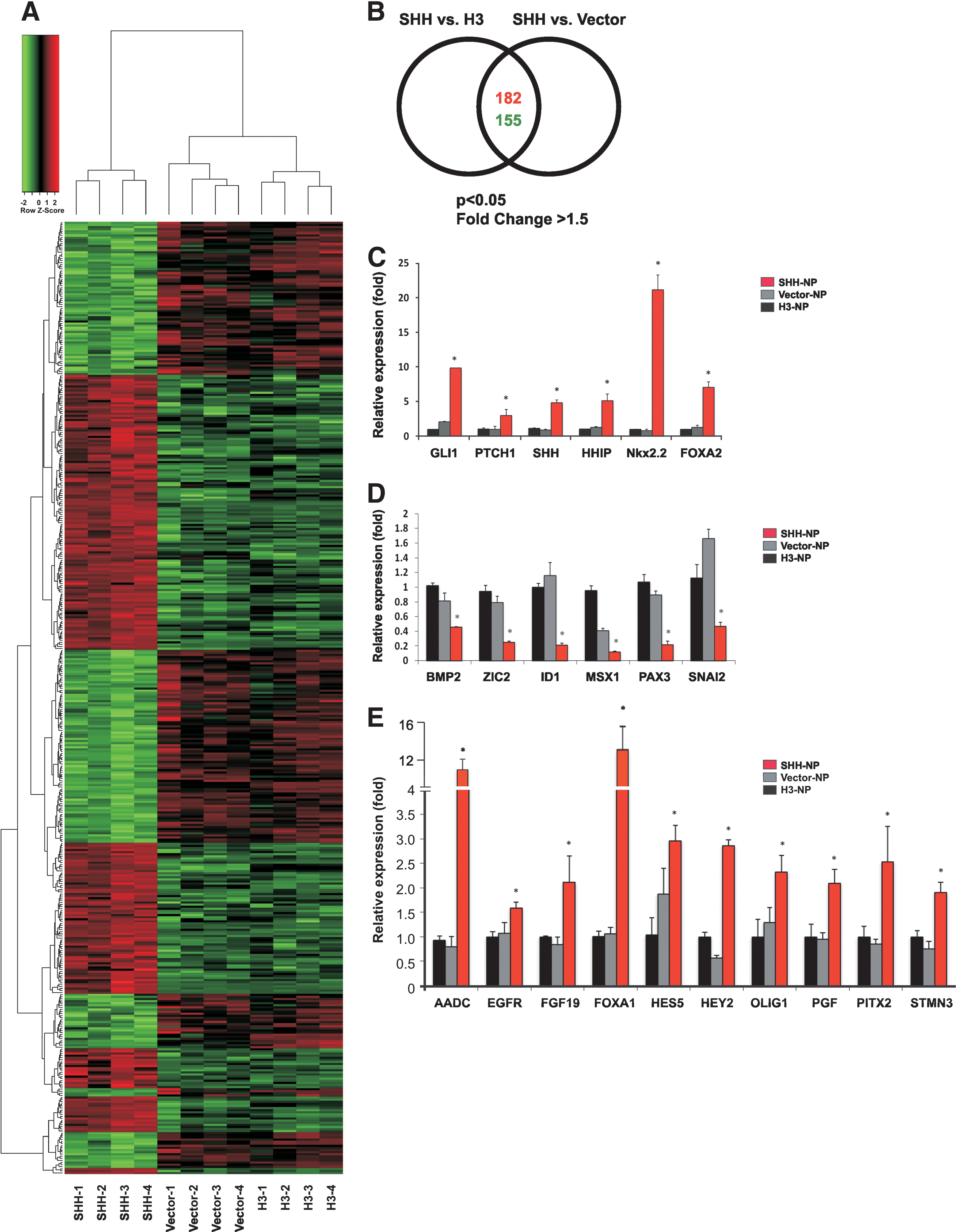
Genome-wide gene expression profiling of SHH-NP.
The DEG were then categorized according to the GO biological processes and the top 8 categories of genes enriched in SHH-NP were cellular developmental process, nervous system development, generation of neurons, neurite morphogenesis, neuron development, cellular morphogenesis during differentiation, and cell adhesion (Supplementary Fig. S4 and Supplementary Table S4). For better resolution of their potential function, the selected DEG from the GO categories were clustered into more precise functional group (Supplementary Table S5). This analysis showed that the potential SHH target genes were those involved in neural induction, NSC proliferation, dorsal–ventral patterning, DA neuron development and function, axon guidance, and neural development.
In a comparison with a recent study that conducted transcriptional profiling of Shh responsive genes in neuralized mouse EB [39], several genes identified in that study to be upregulated by Shh overlapped with those described here, including DDC, EBF3, FABP7, NKX6-1, NR2E1, NTN1, OLIG1, SLIT2, and STMN3 (Supplementary Table S6). Vokes et al. also described data from another independent study that listed several genes that were downregulated by Shh in the mouse neural tube [39]. Downregulated genes observed in both our SHH-NP and that listed in their study include CNTNAP2, FAP, GLI3, MSX1, MSX2, PRRX11, SNAI2, TWIST1, ZIC2, and ZIC5 (Supplementary Table S6). These conserved genes observed between species could prove to be important in SHH-regulated neural differentiation and warrant more in-depth studies in the future.
The transcriptome profiling confirmed the upregulation of the canonical target genes of the SHH pathway in SHH-NP, namely, PTCH1, GLI1, and HHIP, as anticipated. There was also a high fold change of neural tissue-specific target genes FOXA2 and NKX2.2 (Supplementary Tables S5 and S6). Similarly, there was downregulation of the known negatively regulated genes of the pathway, including BOC and CDON [40] (Supplementary Tables S5 and S6). Real-time PCR analysis was carried out to confirm the differential expression profile of 6 known SHH target genes and 16 selected genes, and the results paralleled the findings of the transcriptome profiling analysis (Fig. 5C–E). To determine if the SHH-regulated genes observed in the microarray experiment are broadly observed, the differentiated expression patterns of the selected genes were analyzed in the human iPSC (hiPSC)-derived NPs treated with recombinant SHH (Supplementary Fig. S5). There was a strong correlation in the gene expression pattern between hESC- and hiPSC-derived NP in response to SHH treatment. Together, this provided greater confidence that the other DEG found by the transcriptome profiling analysis were valid targets of the pathway and could be expected to play a role in SHH-dependent neural differentiation.
To understand the molecular mechanisms by which SHH controls hESC neural differentiation and how SHH activity controls the genes that showed differential expression in our profiling, in particular genes that exhibited an upregulated expression, we have to determine whether they are directly activated by the GLI proteins or are induced by secondary transcriptional events downstream. The first line of investigation in this direction will involve the identification of the consensus Gli-binding motif with the regulatory sequences of these genes. To do so, in silico analysis using TRANSFAC match program was carried out to identify GLI consensus binding sites on the 5′ upstream and 3′ downstream regulatory elements of DEG. This GLI consensus binding sequence GACCACCCA [41] was shown to be bound by all 3 GLI proteins [42]. Of the 182 upregulated genes found in SHH-NP, 129 genes contained at least 1 putative GLI-binding site within the 5 kb of upstream sequences relative to the transcription start sites (TSS) and 123 genes had at least 1 putative GLI-binding site within the 5 kb of downstream sequences relative to the TSS (Supplementary Table S7). We then further investigated the overall location of GLI-binding sites in the DEG, and results showed that these genes did contain the GLI-binding sites and evenly distributed within their upstream and downstream regulatory element sequences (Supplementary Fig. S6). This finding substantiates the view that their expression is indeed directly induced by SHH activity. Together, it suggests that they are potential direct targets of SHH signaling.
Of the potential direct target genes of SHH arising from our bioinformatics search, well-established direct SHH target genes were identified. They are PTCH1 GLI1, FOXA2, FOXF1, NKX2-2, and SHH, which all had 4 or more putative GLI-binding sites on their 5′ promoter region (Supplementary Tables S5 and S7), further confirming the computational accuracy of the analysis [39,43,44]. Other genes, such as HES5, FOXA1, OLIG1, STMN3, and FGF19, had been reported to be induced by SHH in other tissue or organisms [39,45 –48]; our study showed that these genes contained 4 or more putative GLI-binding sites and were involved in neural induction and neuron development (Supplementary Tables S5 and S7). More importantly, HEY2, PGF, and FZD9 were identified as novel SHH target genes in our analysis. Thus, SHH-mediated GLI signaling appears to transcriptionally regulate a subset of genes with specific functions in neural induction and development, NSC proliferation, DA neuron function, and axon guidance (Supplementary Tables S5 and S7).
Interestingly, HEY2 and HES5, NOTCH signaling transcription factors, were identified to be potential direct target genes of GLI, with 3 and 6 putative GLI-binding sites in their 5′ promoter region, respectively (Supplementary Tables S5 and S7). Further, HEY2 and HES5 expression levels were increased in SHH-NP (average 5.2-fold increase compared with control Vector-NP and H3-NP; Supplementary Tables S5 and S6). Several NOTCH transcription factors have been demonstrated to be induced by SHH in other cell types. These include Hes1 and Hes5 in retinal explants [47], and Hes1 in mesenchymal and NSC [49]. As the NOTCH signaling pathway is important in the maintenance of NSC during development [50,51], it suggested that there was a cross-talk between these pathways in hESC-derived NP.
HEY2 is a direct target gene of SHH through GLI1 protein binding
To identify whether there was a specific protein–DNA interaction between GLI1 and the 5′ promoter region of HEY2, EMSA was performed. A probe within the HEY2 promoter containing the putative GLI-binding site was used. Results showed that the HEY2 probe specifically shifted in the presence of nuclear extracts of GLI1-overexpressing 293FT cells and the shifted band were abrogated by nonlabeled competitor or HEY2 probe with mutated GLI-binding site (Fig. 6A). Supershift assay using hESC-derived NP nuclear extracts also showed that there was reduced mobility of the DNA–protein complex after incubation of complexes with anti-GLI1 antibody (Fig. 6B). EMSA was also performed for 2 other putative target genes OLIG1 and STMN3 and a similar shift was observed (Supplementary Fig. S7). This provides further evidence that OLIG1 and STMN3 are indeed direct targets of GLI.
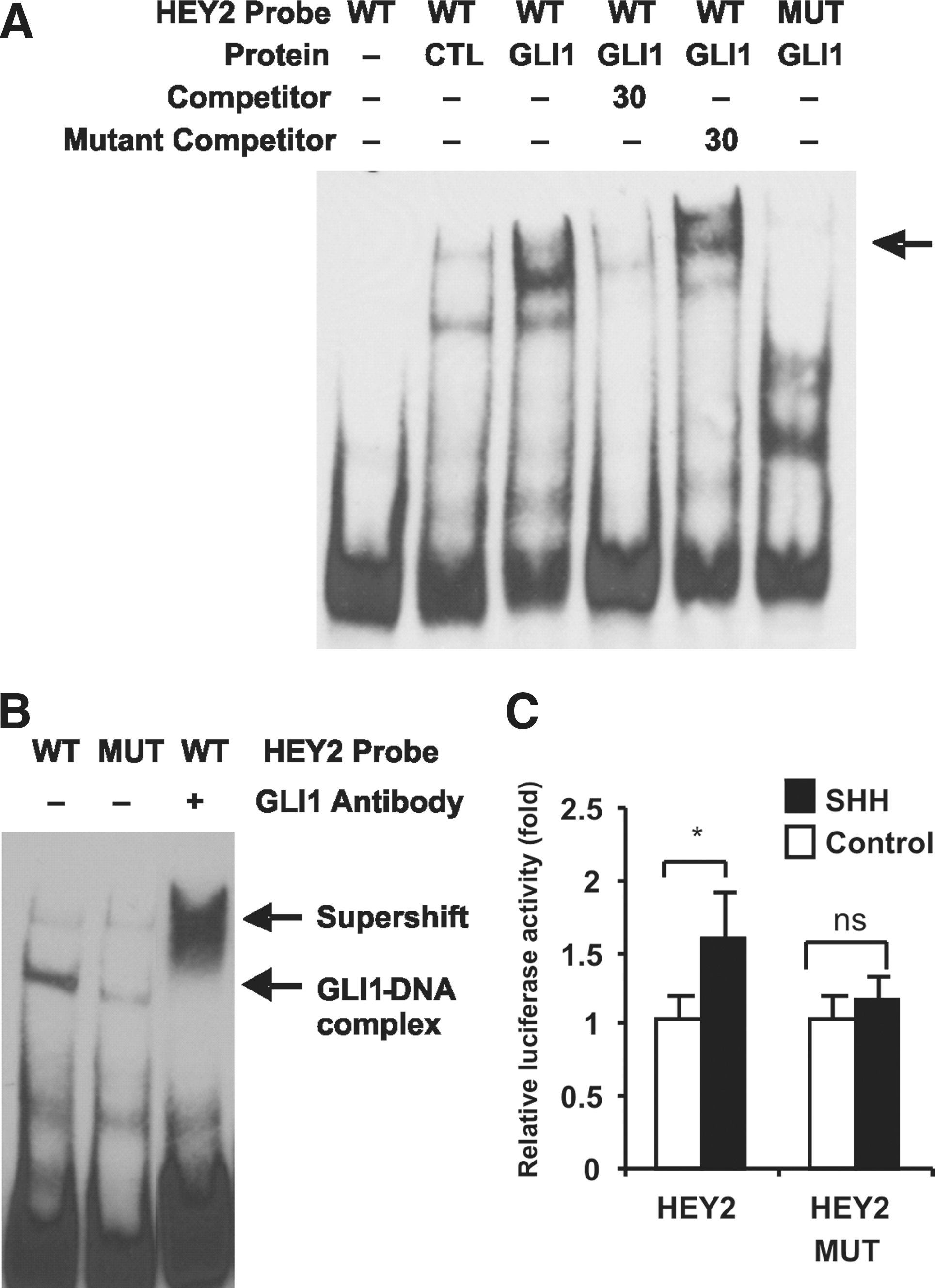
GLI1 binds to and regulates the HEY2 promoter.
To establish a functional role between the GLI-binding site and SHH-mediated HEY2 promoter activity, mutagenesis was performed to generate mutated luciferase reporter construct driven by HEY2 promoter with disruptions within the GLI-binding sites. Cotransfection of the wild-type HEY2 promoter luciferase plasmid with the SHH expression plasmid resulted in a 1.6-fold increase in luciferase activity (Fig. 6C). Mutagenesis of the GLI-binding site resulted in a reduction in luciferase activity compared with wild-type, indicating specificity of the transcriptional activation on HEY2 promoter by SHH-mediated GLI-binding (Fig. 6C). Therefore, the results showed that GLI1 directly binds to HEY2 promoter and regulates HEY2 expression. This evidence strongly suggests that HEY2 is a direct target gene of SHH signaling in hESC-derived NP.
HEY2, a member of the basic helix-loop-helix transcription factor family, is a key target gene of the NOTCH pathway, and its upregulation is associated with active NOTCH signaling [50]. Our results suggest that SHH may be able to activate the NOTCH pathway through its transcription factor HEY2. It is not known if the induction of HEY2 by SHH occurs independently of an active NOTCH pathway since the NOTCH pathway has been demonstrated to be endogenously active in hESC-derived NP [52]. However, there is evidence of cross-talk of the SHH and NOTCH pathways in hESC-derived NP, as transcriptional profiling of NP derived from Notch pathway-activated Hes5-reporter transgenic mESC showed enrichment for Shh pathway genes Smo, Gli2, and Gli3 [53]. The role of the NOTCH pathway in proliferation has been demonstrated in hESC-derived rosettes whereby inhibition of NOTCH signaling decreased the number of rosette-like structures in culture [54]. A recent study showed that hESC-derived rosettes treated with the NOTCH ligands, JAG1 and DLL4, plus SHH had the most robust growth compared with those treated with other signaling molecules like retinoic acid (RA), Noggin, and WNT3a [10]. SHH could also act indirectly via activation of Notch signaling, as shown in cerebellar granule cell proliferation [55]. Our recent findings indicated that overexpressing HEY2 in hESC increases the number of NP (unpublished data). Together, our results suggest that there is a cross-talk between NOTCH and SHH signaling pathways in maintaining proliferation of hESC-derived NP.
Conclusion
In summary, overexpression of SHH in hESC resulted in an enriched NP population that had appropriate NSC identity. Further differentiation of the NP showed that overexpression of SHH led to an increased production of DA neurons, which, we postulate, was due to the higher starting NSC population in SHH-overexpressing NP. These findings suggest that apart from its known function in neural subtype specification, SHH when present at the earlier stages of neural differentiation is able to promote NP expansion. Microarray and GLI-binding site analysis of the SHH-overexpressing NP revealed an extensive transcriptional network downstream of SHH activation, which could potentially mediate the multiple functions of SHH during hESC neural differentiation. HEY2, a target gene of the NOTCH signaling pathway, was identified in this study to be a novel direct target of the SHH pathway. Thus, this systematic study of the role of SHH in neural differentiation and the target gene networks that SHH controls has provided insight for the future hESC differentiation processes for regenerative medicine.
Footnotes
Acknowledgment
This work was supported by the Biochemical Research Council of A*STAR (Agency for Science Technology and Research), Singapore.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.