
Abstract
The regulation of cell proliferation in the external granular layer (EGL) of the developing cerebellum is important for its normal patterning. An important signal that regulates EGL cell proliferation is Sonic hedgehog (Shh). Shh is secreted by the Purkinje cells (PC) and has a mitogenic effect on the granule cell precursors of the EGL. Deregulation of Shh signaling has been associated with abnormal development, and been implicated in medulloblastomas, which are tumors that arise from the cerebellum. Given the importance of the Shh pathway in cerebellum development and disease, there has been no systematic study of its expression pattern during human cerebellum development. In this study, we describe the expression pattern of Shh, its receptor patched, smoothened, and its effectors that belong to the Gli family of transcription factors, during normal human cerebellum development from 10 weeks of gestational age, and in medulloblastomas that represents a case of abnormal cell proliferation in the cerebellum. This expression pattern is compared to equivalent stages in the normal development of cerebellum in mouse, as well as in tumors. Important differences between human and mouse that reflect differences in the normal developmental program between the 2 species are observed. First, in humans there appears to be a stage of Shh signaling within the EGL, when the PC are not yet the source of Shh. Second, unlike in the postnatal mouse cerebellum, expression of Shh in the PC in the postnatal human cerebellum is downregulated. Finally, medulloblastomas in the human but not in patched heterozygote mouse express Shh. These results highlight cross-species differences in the regulation of the Shh signaling pathway.
Introduction
T
A role for Shh in normal cerebellum development was initially hypothesized based on the observations that mutations in the Patched (Ptc) receptor was associated with sporadic medulloblastomas [1,2] and were also seen in Basal Cell Nevus Syndrome that was associated with an increased frequency of developing medulloblastomas [3]. Subsequent studies elucidated that Shh was secreted by Purkinje cells (PC) and bound to the Ptc receptors that were present on the GCP. The binding of Shh to Ptc relieved the inhibition of the Ptc receptor on Smoothened (Smo), which through a family of zinc finger transcription factors known as Glis downstream triggered a proliferation response in the GCP [4,5].
In the mouse, PCs are born in the ventricular zone from about E11 to E13. These begin to express Calbindin by around E15.5, and Shh by E17.5 [4,6]. By P0, the PCs begin to coalesce into a monolayer beneath the EGL layer, and by P8 they are well differentiated and clearly recognizable with their large cell bodies and a developing dendritic arbor that extend into the molecular layer [7]. Relative to the development of the PCs, the GCP of the EGL originate from the rhombic lip and in the mouse these cells begin to migrate out of the rhombic lip area to establish the EGL between E13.5 and E15.5 [8,9]. Over the next 2 postnatal weeks, the GCP continue to proliferate and produce postmitotic GCs until the EGL disappears by the third postnatal week [10].
In contrast, in humans the development of the PCs takes place over an extended period [11]. PCs are in an immature neuroblastic stage until around the 16th gestational week (g.w.). Thereafter, until around the 28th g.w. they assume a multipolar morphology, and at this stage they also rearrange themselves into a monolayer. The final stage of differentiation that involves the maturation of its characteristic dendritic tree continues into the first postnatal year [11]. Even keeping the absolute size of the dendritic arbor constant, PCs in the human cerebellum take 6-fold longer to mature than in the mouse [11]. Even though the PCs have an immature morphology until the 16th g.w., as early as the 10th g.w, the EGL in the human is clearly visible and is in fact several layers thick and at 22 g.w., 30% of the EGL cells are labeled with thymidine-H3 [12]. The inward migration of postmitotic GCs begins as early as the 11th g.w even as the EGL continues to expand. The formation of the IGL continues postnatally until the last of the GCs finish their migration inward to the IGL around 8 months of age [12]. Thus, unlike in the mouse, human GCP expansion occurs mainly in utero [12].
From the above summary, it is seen that the relative time course of differentiation of PCs and GCs in human and mouse are different. Given this, it is not clear whether Shh is involved in the very early stages of EGL proliferation in humans since the PCs are immature and have not finished migrating. Unlike in the case of the rodent [13,14] there has been no systematic characterization of Hedgehog-Patched signaling pathway molecules during human cerebellum development and thus it is also not known how expression of Shh correlates with the timing of the development of the EGL and IGL.
In addition, although the Hedgehog-Patched signaling pathway has been implicated in human medulloblastomas, it is not clear whether Shh expression persists in cases of abnormal cell proliferation. Given this, the following questions are being addressed: (1) Are the PCs at the neuroblast stage of development capable of producing Shh? (2) How is this correlated with evidence of Shh signaling in the EGL? (3) Is the disappearance of the EGL layer in humans correlated with the downregulation of Shh? (4) Is Shh upregulated in cases of human medulloblastomas that represent a case of abnormal cell proliferation?
Materials and Methods
Tissue
All samples used in this project were collected in accordance with the guidelines laid down by the Institutional Human Ethics Committee, National Brain Research Centre, Manesar, which have been framed by the Indian Council of Medical Research, India. Fetal and postnatal cerebellum of different ages (10–13 weeks, n=3; 14–19, n=4, 20–32 weeks, n=20; 33–39 weeks, n=15; 1 month–5 postnatal years, n=6, Medulloblastoma, n=8) was obtained from the Department of Pathology, All India Institute of Medical Sciences, New Delhi, and Army Base Hospital, New Delhi (Supplementary Table S1; Supplementary Data are available online at
For mouse development studies, C57BL/6J mice were crossed and the day of plug was taken as embryonic day (E) 0.5. Embryos were dissected out at days 11.5, 13.5, 15.5, and 18.5. The postnatal mice (P2, P4, and P8) were perfused using 4% PFA and their cerebella were dissected out and processed for cryosectioning. The procedure was identical to that mentioned earlier.
Immunohistochemistry
Antigen retrieval method
Following dewaxing and rehydration, the sections were subjected to antigen retrieval using an antigen unmasking solution (Vector Laboratories). Slides were placed in a Coplin jar containing a preheated solution of the antigen unmasking solution in PBS (1:70). The jar was then placed in a pressure cooker and heated for another 10 min (a minimum of 2 whistles). After this, the slides were cooled at room temperature and washed thrice in 1×PBS.
Histological analysis
Immunohistochemistry was performed on serial sections. After antigen retrieval, sections were quenched using 3% hydrogen peroxide for half an hour to remove endogenous peroxidase activity. This step was skipped for sections on which immunofluorescence was carried out. The sections were blocked with 5% serum for 2 h to reduce nonspecific binding. Serum from the host species of the secondary antibody was used for the blocking step. Five percent bovine serum albumin was used for blocking, in colocalization studies. This step was followed by incubating the sections in primary antibody solution for 16–48 h at 4°C at optimum dilutions in a humid chamber (dilutions mentioned below). The primary antibodies used in this study include Shh (Santa Cruz, 1:100), Ptc, Smo, Gli-1, Gli-2 (All Abcam, 1:250), Calbindin (Swant, 1:3,000), and MATH-1 (1:200; Chemicon). Sections were then washed with 1×PBS thrice after which they were incubated in appropriate secondary antibodies–biotinylated (1:250; Vector Laboratories) or fluorescent dye labeled (Alexa fluors 488 and 594, 1:1,000; Molecular Probes), for 1½ hours. The sections were washed with 1×PBS thrice, after which sections labeled with the fluorescent dye-labeled antibody were dried and mounted in Vectashield mounting media with 4′,6-diamidino-2-phenylindole (DAPI) (Vector Laboratories). The ones labeled with the biotinylated antibody were also washed with 1×PBS thrice, following which they were incubated for 2 h in the ABC reagent (1:50, Avidin-Biotin complex reagent; Elite ABC kit; Vector Laboratories). Lastly, sections were washed with 1×PBS and developed using the 3,3′-diaminobenzidine (DAB) Kit (Vector Laboratories) for 1–2 min. After the sections were rinsed with 1×PBS, they were air-dried overnight and cover-slipped with distyrene, plasticizer, xylene (DPX) mounting medium. For each antibody, one section was used as a negative control where in the section was incubated with all the above solutions except the primary antibody.
For hematoxylin and eosin staining (H and E staining), sections were stained in hematoxylin for 5 min, after which they were washed in running tap water. These sections were then differentiated in 1% acid alcohol solution and treated with ammonia water, before counter staining with eosin for 1 min. The sections, following staining were then dehydrated for 5 min, followed by immersion in xylene and then cover-slipped with DPX mounting medium.
Two experiments were done to confirm the specificity of the anti-Shh antibody used in this study. First, cerebellum sections from human and mouse were incubated with a mixture of anti Shh antibody (Santa Cruz) and recombinant Shh peptide (R&D Systems), in a 1:1 ratio. This was done to compete out specific binding to Shh with the Shh peptide. Second, the specificity of the Shh antibody was confirmed by using 2 other antibodies to Shh from Abcam (ab50515, ab53281; 1:200).
Western blotting
Radioimmunoprecipitation assay buffer was added to human (26 g.w.) and P6 mouse cerebella (for 1 mg of tissue, 100 μL of buffer was added) and homogenized. The homogenized mixture was subjected to centrifugation. The supernatant (whole protein extract) was collected. The protein concentration was estimated using Bradford's reagent and 60 μg of protein was electrophorosed on 7%–10% polyacrylamide gel. The proteins on the gel that were transferred to polyvinylidene difluoride (PVDF) membrane were probed with primary antibody overnight: Shh (Santa Cruz, Abcam, 1:1,000), Ptc, Smo, Gli-1, and Gli-2 (All Abcam, 1:1,000). After appropriate washes with 1×Tris-buffered saline with Tween 20 (TBST) the blots were probed with secondary antibody (Vector Laboratories) for 2 h. Finally, the blots were washed with 1×TBST and developed using chemiluminescent reagent (Amersham) and exposed to Chemigenius, Bioimaging System (Syngene UK) for developing. Images were captured using Genesnap software (Syngene).
Reverse transcriptase-polymerase chain reaction
The total RNA of an 18 g.w fetal cerebellum was isolated using Trizol Reagent (Sigma). cDNA was constructed by using reverse transcriptase (Fermentas) and random primers. Amplification was performed with about 500 ng cDNA. Specific primer pairs for Shh (forward, GAAACTCCGAGCG ATTTAAGGA; reverse, GGCCCTCGTAGTGCAGAGA), Ptc (forward, GGGGGAGAACCTTCGCATC; reverse, GCCCA GATACTGTTCCCAGAA), Smo (forward, CGGAACTCGA ATCGCTACCC; reverse, CAAGCCTCATGGTGCCATC), and Gli-1 (forward, CCAGGAATTTGACTCCCAAGAG; reverse, CAGCATGTACTGGGCTTTGAA) were designed and used. The amplification was done for 40 cycles (94°C 30 s, 60°C–65°C 30 s, and 72°C 1 min).
Microscopy
All bright-field images were captured using Leica DFC 320 and the images were analyzed using the Leica IM50 software. Fluorescent images were captured on a Zeiss Axioplan 2 microscope using an Axio Cam HRC CCD camera, and analysis was done using the Zeiss KS 400 3.0 software. The images required minor adjustment of contrast and brightness. No additional image alteration was necessary.
Results
Shh-patched signaling pathway in the early cerebellar development
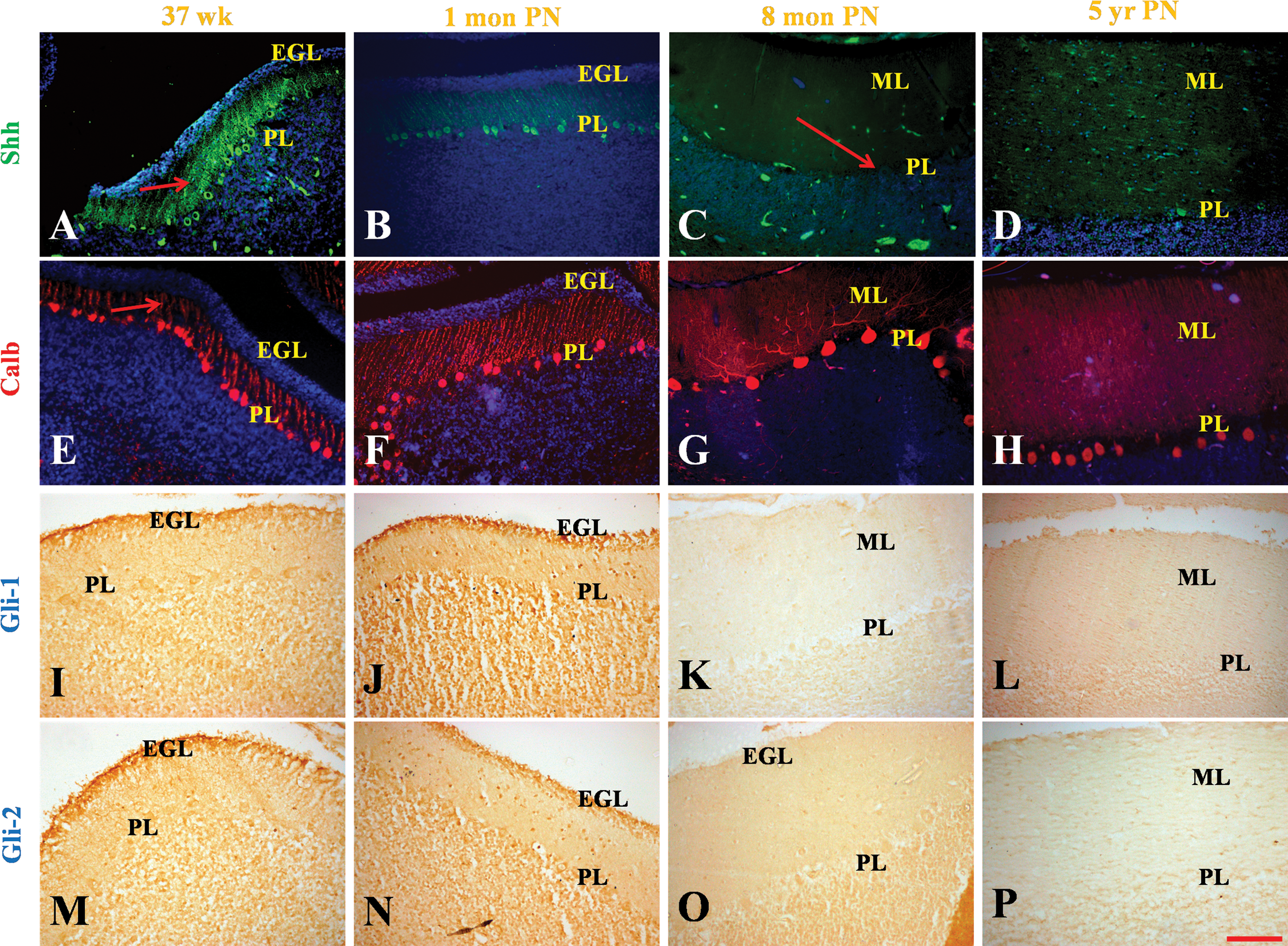
To address our first question, the endogenous expression of Shh in the developing human cerebellum was characterized (Figs. 1A1–E1, and 2A–D). The youngest specimens that we had were of 10 g.w. In these samples Shh was detected in a distinct thick band below the pia which could be identified as the EGL based on the closely packed small cell bodies seen by DAPI stain (Fig. 1A1, box). Apart from the strong expression in the EGL, scattered cells are also seen in the deeper layers of the EGL. This is not seen in the controls where no primary antibody was added. Immunohistochemistry for Calbindin showed no expression of Calbindin at the 10 g.w. stage (Fig. 1A2, box). Two weeks later Shh continued to be detected strongly in the EGL (Fig. 1B1, arrow) and is also found in the deeper layers of the cerebellum (Fig. 1B1, asterisk). Staining for Calbindin revealed specific staining of the deeper layers and also showed that the Shh expression overlapped with Calbindin, suggesting that these are the migrating PC neuroblasts that express Shh (Fig. 1B2, arrow). The 14 g.w. sample showed Shh expression in the EGL and at this stage expression of Shh was also seen in a restricted multilayer band of Calbindin expressing cells (Fig. 1C1, arrow). There was also a decline in expression of Shh in the EGL from 14 to 17 g.w. (Fig. 1C1, D1, arrow). At 20 g.w., Shh expression could no longer be detected in the EGL (Fig. 1E1, arrow). At this stage strong expression of Shh was seen in the PL below the EGL, a region that coincided with a narrow band of Calbindin expressing PCs (Fig. 1E1, E2, box). This expression pattern is consistent with what is seen in postnatal mouse cerebellar development [14,15]. H and E staining of the cerebellar sections was done at the relevant ages to correlate histology with the appearance of Calbindin-positive cells (Supplementary Fig. S1A–H).

Expression of Shh
Expression of Shh
To prove the specificity of the Shh antibody used in our study, we incubated cerebellar sections from human and mouse with a mixture of anti Shh antibody (Santa Cruz) and recombinant Shh peptide, in an equimolar ratio. We detected no staining under this condition, a result identical to what was observed in the negative control in which no primary antibody was added (Supplementary Fig. S2, compare SA2–SC2 with SA3–SC3). The positive control showed specific staining confined to the EGL and PL at 12 g.w. (Supplementary Fig. S2A1), and in the PL alone, at 37 g.w. in the human (Supplementary Fig. S2B1), and at P6 in the mouse (Supplementary Fig. S2C1). We also incubated human cerebellar sections of age 37 g.w., with 2 other anti-Shh antibodies from Abcam found that all the antibodies gave an identical staining profile to what was obtained with the Santa Cruz antibody used in our study (Supplementary Fig. S3A, B), with Shh expression being confined to the PL. Results from the western blot showed that all 3 antibodies recognize proteins of the same molecular weight, which is similar to the observed molecular weights of the bands given in the specification sheet (Supplementary Fig. S4). These molecular weights are different from the predicted molecular weight of Shh due to the extensive post-translational modification that Shh undergoes [16 –19].
Expression of Gli1 and Gli2 showed that they were all strongly expressed in the EGL starting at 10 g.w. (Fig. 1A3, A4). Ptc and Smo expression was seen in the EGL during early cerebellum development (Supplementary Fig. S3C–J). Expression of both Gli1 and Gli2 was weak in the PL as compared to the EGL with Gli2 expression not being detected in the PL at 20 g.w (Fig. 1, compare Fig. D3, E3 with D4, E4, arrow). Reverse transcriptase (RT)-polymerase chain reaction on an 18 g.w. cerebellar tissue, using specific primers for Shh, Ptc, Smo, and Gli-1 indicated expression of all of the above genes (Supplementary Fig. S3O). We further validated these results by performing a western blot using the same antibodies (Supplementary Figs. S3P and S4).
Shh-patched signaling pathway in late human cerebellar development
Expression analysis of the 37 g.w. fetus revealed strong expression of Shh in the PL. These were now easily distinguished by their large cell bodies and apical dendrites that were both positive for Shh. Calbindin expression was coincident with Shh expression and clearly revealed the developing dendritic arbor (Fig. 2A, E, arrow). When a 1-month-old postnatal sample was compared to the 37 g.w. sample, a dramatic reduction in expression of Shh was seen in the PL (Fig. 2B). In addition, compared to the 37 g.w. sample, the 1 month postnatal sample showed that the PC cell bodies were more widely spaced (Fig. 2F). Concomitantly the dendritic arbor was also spread out and the dendrites were longer as they traversed the expanding molecular layer. At 8 months postnatal, PL could be clearly seen by Calbindin that labeled both the cell body and the processes (Fig. 2G). However, expression of Shh was no longer detectable (Fig. 2C, arrow). Expression of Gli1 (Fig. 2I, J) and Gli2 (Fig. 2M, N) in the EGL was seen in the outer regions of the EGL at 37 g.w. and 1 month postnatal specimens. At 8 months postnatal age, an EGL layer was no longer be seen by DAPI staining and expression of Gli1 and Gli2 was no longer detected (Fig. 2K, O). The PCs continued to express Calbindin, which could be seen in the cell bodies and the long dendritic processes that were all perpendicularly oriented to the plane of section (Fig. 2G). In addition, it can be seen that scattered PCs also express Smo (Supplementary Fig. S3J) and Gli1 (Fig. 2I). The presence of Smo and Gli1 immunoreactivity in some PCs may indicate autocrine Shh response.
Expression of Hedgehog-patched signaling molecules in early cerebellar development in mouse
Expression of Shh in the developing mouse cerebellum was analyzed by immunohistochemistry. Shh expression could not be detected at E11.5 (Fig. 3A) and E13.5 (Fig. 3D) in the developing mouse cerebellum. While at E11.5 the migration of EGL progenitors from the rhombic lip is taking place, the presence of a streak of EGL in the cerebellar anlagen can be seen at E13.5 (Fig. 3G, I arrow). The EGL at this stage does not express Shh. Although the PL is conspicuous by its absence one does find a stream of Calbindin-positive PCs migrating in the direction toward the cerebellar anlagen (Fig. 3E, K, arrow). Shh expression could, however, be detected in the EGL at both E15.5 and E18.5 (Fig. 4A, B, arrow). Calbindin stain at these ages revealed diffuse staining in the inner regions of the cerebellum comparable to what is observed in the human at 12 g.w. (Fig. 4A, B, asterisk). Thus, the staining pattern at E15.5 and E18.5 resembled early cerebellar development in humans. Expression of Gli1 and Gli2 showed that these transcription factors were not expressed in the EGL at E15.5 (Fig. 4C, E, arrow). However, at E18.5, Gli1 expression was detected in the EGL. The intensity of Gli1 expression in the EGL (Fig. 4D, arrow) was higher in those regions of the EGL that had Calbindin-positive cells expressing Shh lying underneath (Fig. 4B, asterisk). Gli2 was expressed more strongly than Gli1 in the EGL and showed a more restricted pattern than Gli1 in the EGL (Fig. 4F, arrow) being almost exclusively expressed in regions the overlay Calbindin-positive cells expressing Shh (Fig. 4B, asterisk). Additionally, both Gli1 and Gli2 were expressed throughout the PL (Fig. 4D, F). Thus, although expression of Shh and Calbindin in the developing mouse cerebellum resembled that of the early human cerebellum development, expression of Glis was quite different. Whereas a strong band of Glis was expressed throughout the EGL as early as 10 g.w. in humans, Gli expression in the mouse did not acquire this expression pattern in utero.
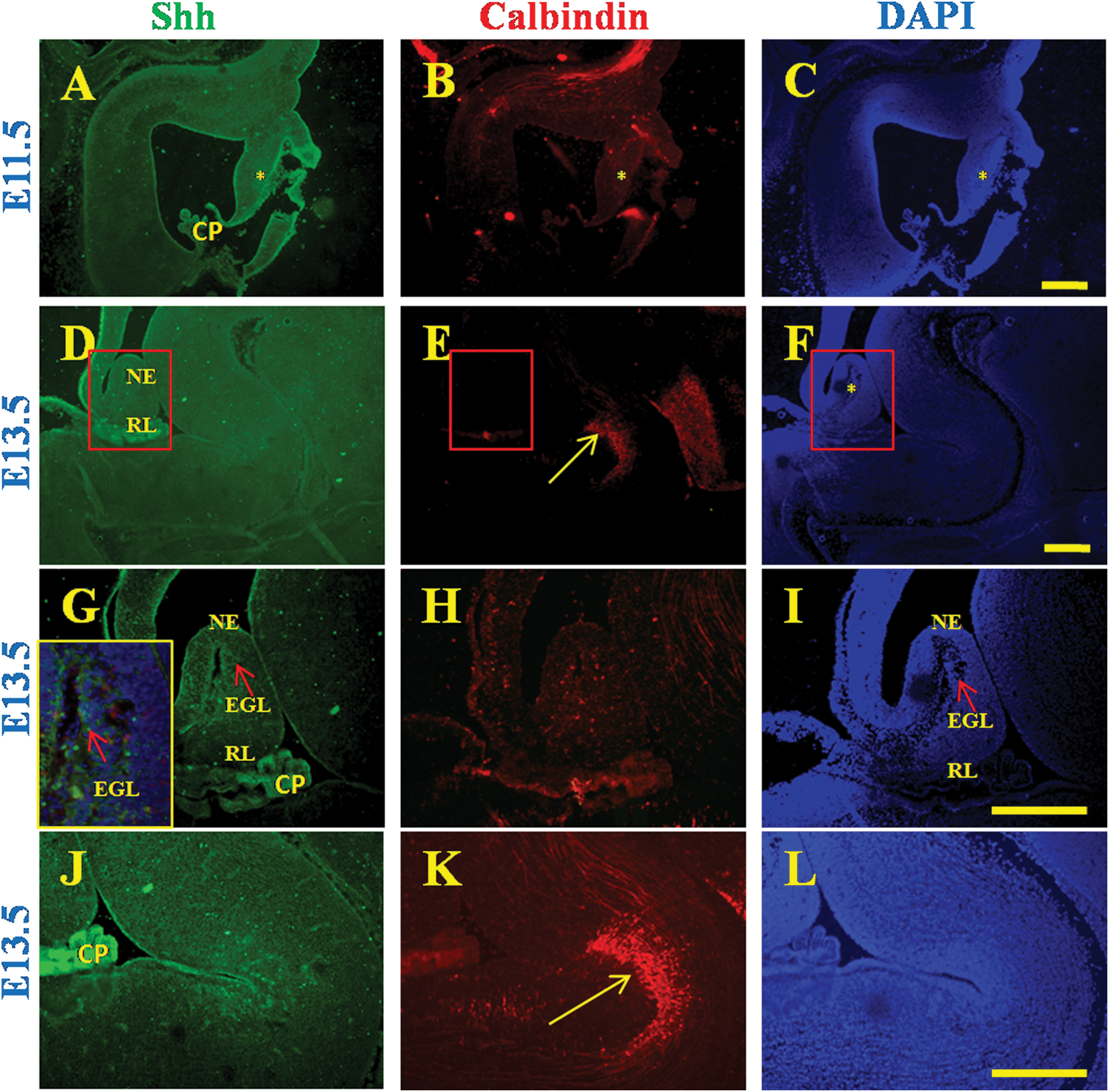
Expression of Shh and Calbindin during early cerebellum development in mouse. Shh expression is absent in the cerebellar anlagen at embryonic ages 11.5

Expression of Shh
Expression of Hedgehog-patched signaling molecules in postnatal cerebellar development in mouse
Shh expression was not detected in the EGL at P2 (Fig. 5A) and P4 (Fig. 5D). However, the PL was strongly positive for Shh (Fig. 5A, D). Thus, the early postnatal expression of Shh and Calbindin in the mouse cerebellum resembled that of 20 g.w. cerebellum in human.

Expression of Shh
Expression of Shh in human medulloblastomas
Expression of Shh was analyzed in cases of medulloblastoma from patients aged 7–36 years. In particular, we were interested in tumors that had been removed along with the normal adjoining cerebellum. It was found that the tumor-containing portion of the cerebellum showed strong expression of Shh (Fig. 6A blue box and Fig. 6D arrow). Further the region of the cerebellum closest to the tumor showed remnants of EGL that also expressed Shh (Fig. 6A red box, Fig. 6C arrow). However, in a normal cerebellum of the equivalent age, the EGL was no longer present. Although the EGL remnant adjoining the tumor showed expression of Shh, the PL did not express Shh (Fig. 6C, PL). The PL, however, expressed Calbindin (Fig. 6E, F, PL). The portion of the cerebellum farthest from the tumor had no EGL (Fig. 6A yellow box, Fig. 6B and E), and was comparable to a normal age-matched cerebellum. This region as expected showed no expression of Shh (Fig. 6B) in the PL that was identified by expression of Calbindin (Fig. 6E). Finally, the tumor also expressed MATH-1, a marker of EGL (Fig. 6H, I). Expression of Shh (Fig. 6G) and MATH-1 (Fig. 6J) was found in both desmoplastic medulloblastoma.

Expression of Shh in human medulloblastoma
Expression of Shh in murine medulloblastomas
Medulloblastoma sections from the Ptc +/− mice did not express Shh in the EGL or PL (Fig. 7A, C), although the PL was found to express Calbindin (Fig. 7B, D).
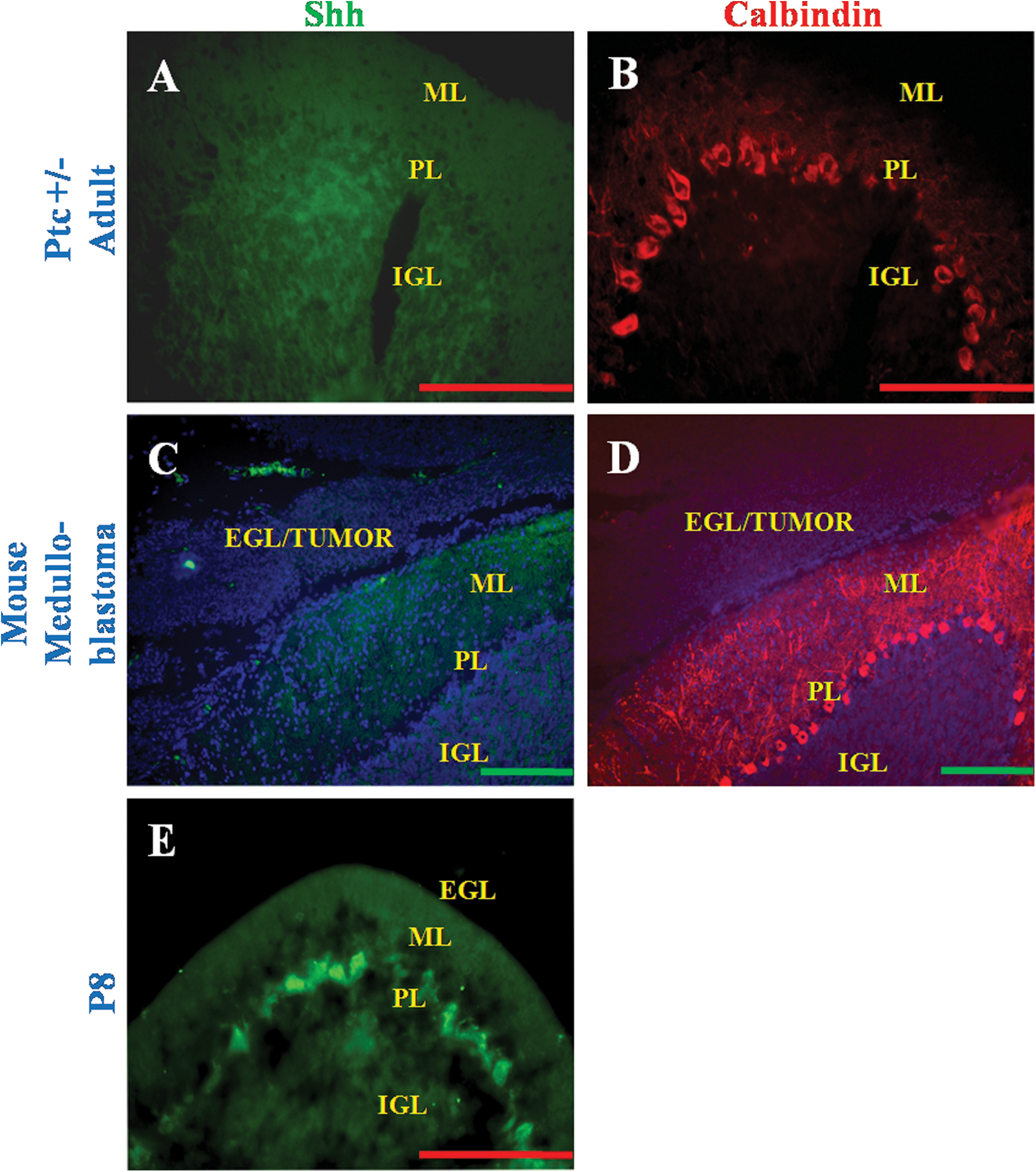
Expression of Shh in Ptc +/− model of mouse medulloblastoma
Discussion
In this study, we have characterized the Shh signaling pathway during human cerebellar development. We have also characterized expression of Hedgehog-Patched pathway molecules in both murine and human medulloblastomas. All antibodies that have been used in this study have been previously validated on sections in other studies [20 –24]. We also carried out a biochemical validation of the antibodies on human and mouse cerebellum using western blot (Supplementary Fig. S3P and S4). Studies on human cerebellum development have shown that the PCs go through different stages of maturation [11]. The first stage of PC differentiation lasts till around the 16th week of gestation and our study shows Calbindin expression in the intermediate zone beneath the marginal layer during this time [12]. As the cells migrate to their final destination from the 14th to the 20th week, Calbindin expression gradually became restricted. In the third trimester Calbindin expression was confined to PC bodies and their dendrites. Expression of Shh was detected along with Calbindin as early as the 12th week of gestation when these cells had not yet migrated to their final location. Although PCs could be identified by Calbindin expression through the first postnatal year, Shh could only be detected in the first postnatal month and its expression could not be detected at 8 months. This suggests that expression of Shh is dramatically downregulated in the PC soon after birth. Therefore, at the last stage of PC differentiation, especially the time between birth and the first postnatal year, when the Purkinje dendritic arbor is acquiring its characteristic shape, Shh is downregulated. The fact that Shh is downregulated upon birth in humans but not in mouse may be related to the fact that GCP proliferation occurs in utero in humans, but after birth in mouse. At present it is not known what causes this downregulation of Shh.
In the developing mouse cerebellum, Calbindin expression was detected in the PL by immunohistochemistry at E15.5. This is in agreement with previous results that have detected Calbindin by in situ hybridization by about E15 [6]. Our immunohistochemistry results further showed that Shh was expressed by the PC at E15.5 and that Shh expression continued after birth, coincident with the time when the GCP are expanding, a process that occurs after birth in mice. One of the important observations of this study is that we detect immunoreactivity to Shh in the human EGL during early PC maturation, which suggests that there is a distinct stage in human cerebellum development, where Shh is present in the EGL, but not in the PL. Characterization of Shh expression in early human cerebellum development shows the presence of Shh in the EGL layer until around the 17th week. After the 20th week of gestation, at the time when the PC are entering their next stage of development and arranging themselves into a monolayer, the Shh immunoreactivity in the EGL is decreased. The source of this Shh is as yet not known and could lie either within the EGL or outside it.
In the mouse, contrasting reports have been obtained with respect to the presence of Shh in the EGL. While no expression in the EGL was detected either by the Shh-lacz knockin or by in situ hybridization [25], various groups, including ours, have reported a variable transient Shh expression in the EGL [4,5,7]. In the current study, we detected expression of Shh in the mouse at E15.5 and E18.5 both in the PL and in the EGL by immunocytochemistry, but only in the PL at P2 and P4. This discrepancy in the results between different groups could be due to the difference in techniques used for detection. However, in contrast to the human, we did not detect the presence of Shh solely in the EGL at any stage of mouse development.
We also looked in the EGL, at expression of Ptc, Smo, and the Gli transcription factors, which are the other components of the Hedgehog-Patched signaling pathway [5,26]. In the developing human cerebellum, strong immunoreactivity to Ptc, Smo, Gli1, and Gli2 was detected in the EGL as early as the 10th week of gestation. Expression of the receptors and downstream transcription factors continued even as Shh expression in the EGL was downregulated. This suggests that the source of Shh had at this stage moved exclusively to the PC. As expected, with the disappearance of Shh from the PC and the thinning of the EGL expression of Ptc, Smo, and Gli downstream molecules is no longer observed. Analysis of the mouse embryonic cerebellum showed that Gli1 and Gli2 are expressed in a region specific manner, which is in agreement with previous results using Gli1-lacz and Gli2-lacz to detect expression of these transcription factors [27]. In a previous study, expression of Gli1 and Gli2 followed expression of Shh in the PL [27]. Our observations concur with this result and also show a gradual increase in Gli1 and Gli2 expression that was correlated with expression of Shh in the PL during the postnatal expansion of GCP. Several articles from Joyner's group have shown that Gli1 expression is a readout of Shh activation [27,28]. Thus, in the current study, if we take expression of Gli1 to represent regions where there is activation of the Shh pathway, the results show that Shh signaling in the EGL occurs in the mouse mainly postnatally, coincident with GCP expansion and the source of Shh is the PL. This result validates the approach and agrees with what is known about EGL expansion in the mouse. In contrast, during human cerebellum development our results based on expression pattern of Gli1 suggest that there is Shh signaling very early on within the EGL, where the source of Shh is not derived from the PC.
The strong link between developmental processes and cancer is becoming increasingly evident. The origin of a subset of medulloblastomas is often attributed to the abnormal proliferation of the GCP of the EGL. In this regard, mutations in other Shh pathway genes such as SUFU, Smo, and β TRCP have also been uncovered, suggesting that a deregulation of Shh signaling can lead to medulloblastomas [24,29,30]. Medulloblastomas have been divided into 2 major classes based on morphological characteristics as well as gene expression [31,32]. Other medulloblastoma classifications have also been proposed based on gene expression studies that have divided medulloblastomas into 5 classes [33]. In general, desmoplastic tumors have been associated with Ptc mutations, whereas classical tumors have been associated with the Wnt signaling pathway. In our study, tumors that had been classified histologically as either desmoplastic or classic showed that the part of the cerebellum containing the tumor has strong expression of Shh and MATH1. It is possible that dysregulation of the Wnt signaling pathway could in turn lead to upregulation of the Shh pathway since these 2 pathways are known to interact with each other [34]. Interestingly, the region of the cerebellum proximal to the tumor showed remnants of EGL and expressed Math1 and Shh, whereas the PL directly beneath did not stain positive for the same. In the tumor samples that also happened to have adjoining normal tissue, we saw that there was no EGL and consequently no Math1 or Shh expression, and this area was comparable with a normal age-matched cerebellum. Medulloblastoma samples from Ptc heterozygote mice on the other hand did not express Shh. These results indicate that in human medulloblastomas, there is a persistent expression of Shh, although a larger study needs to be carried out to look at what percentage of human medulloblastomas express Shh. Interestingly, studies in mice have shown that activating the Shh pathway leads to the formation of tumors and that when Shh signaling is activated upstream of Gli transcription factors, Shh can produce tumors even in the absence of Gli [35]. In addition, studies have shown that in order for the Shh pathway to produce medulloblastomas, the cells have to be committed GCP [36].
In mouse cerebellum development, in contrast to the human situation, Shh signaling in the EGL is not strong at a stage when there is rapid expansion of the GCP. In this context it is interesting that a recent study [37] suggested that while the gene expression profile of the normal mature human cerebellum resembles the murine cerebellum at P30–P60 expression profile of human classic and desmoplastic medulloblastomas resemble that of early postnatal murine cerebellum. This is the stage when EGL precursors are responding to a Shh signal that is derived from PC. Therefore, expression of Shh could be a common factor in cerebellar tumors and could cut across tumor categories. Based on these results and the developmental expression pattern, we put forward the hypothesis that a failure to downregulate Shh in the EGL could lead to a persistence of an earlier stage of GCP, where Shh signaling occurs within the EGL. Thus, even as the PCs downregulate Shh postnatally, these early GCP cells could give rise to medulloblastomas. This may not arise commonly in the mouse where Shh signaling in the EGL is not strong at a stage when there is rapid expansion of the GCP. This remains to be tested for a larger set of tumors.
The idea that due to the timing of PC and GC differentiation in human cerebellum development, medulloblastomas could arise from a primitive population of Shh expressing EGL, which requires an additional hit to form tumors, is speculative and needs to be tested. One approach would be to identify markers of the EGL unique to the GCP from the first trimester of gestation to see if they may be expressed in the tumor. A parallel approach would also be to identify mechanisms by which Shh expression in the EGL is lost as this could also help identify new therapeutic targets to treat medulloblastomas.
Footnotes
Acknowledgments
We thank Kiran (AIIMS, New Delhi), Dr. Ausaf Farooqui, Mohammad Sarfaraz Nawaz, Younis Khan (NBRC, Manesar), Prof. Catherine Verney (INSERM U676, Paris, France), Gautam Surya (Tufts University, Boston), Parul Chachra, and Dr. Shubha Tole (TIFR, Mumbai) for their technical help and support. We also express our gratitude to Prof. James Waschek (UCLA) for providing us with the mouse medulloblastoma tissue. This study was supported by DST-IRHPA project number 943 to S.M., NBRC Core Funding, IFCPAR/CEFIPRA funding project number 3803-3 to S.M. and P.G., and a grant from APHP (Contrat d'Interface to Dr. Pierre Gressens). P.H. is supported by a Senior Research Fellowship from the National Brain Research Centre, Manesar.
Disclosure Statement
All the authors have seen and agreed to the content of this article. None of the authors have any potential financial interests or conflict of interest with respect to this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.