
Abstract
Similar to embryonic stem cells, induced pluripotent stem (iPS) cells can differentiate into various cell types upon appropriate induction, and thus, may be valuable cell sources for regenerative medicine. However, iPS cells have not been reported to differentiate into odontogenic cells for tooth regeneration. Here we demonstrated that neural crest-like cells (NCLC) derived from mouse iPS cells have the potential to differentiate into odontogenic mesenchymal cells. We developed an efficient culture protocol to induce the differentiation of mouse iPS cells into NCLC. We confirmed that the cells exhibited neural crest (NC) cell markers as evidenced by immunocytochemistry, flow cytometry, and real-time reverse transcription-polymerase chain reaction. Further, in recombination cultures of NCLC and mouse dental epithelium, NCLC exhibited a gene expression pattern involving dental mesenchymal cells. Some NCLC also expressed dentin sialoprotein. Conditioned medium of mouse dental epithelium cultures further enhanced the differentiation of NCLC into odontoblasts. These results suggest that iPS cells are useful cell sources for tooth regeneration and tooth development studies.
Introduction
O
Induced pluripotent stem (iPS) cells are generated from somatic cells by the simultaneous introduction of several factors; and they differentiate into the 3 embryonic germ layers with an extensive proliferative capacity [4 –6]. This technique allows us to generate pluripotent stem cells without the use of embryos, to overcome rejection problems after implantation of nonautologous cells, and to avoid ethical issues associated with the use of embryonic stem (ES) cells. Recently, many researchers have reported that iPS cells can differentiate into different cell types, such as neurons, cardiac myocytes, and renal lineage cells, under appropriate conditions [7 –9]. Therefore, iPS cells have emerged as potential cell sources for regenerative medicine. However, their potential to differentiate into odontogenic cells has not yet been investigated.
Tooth development involves a series of reciprocal epithelial–mesenchymal interactions between the mandibular epithelium and neural crest (NC)-derived ectomesenchyme [10,11]. The NC-derived ectomesenchymal cells contribute to the dental mesenchymal cells (DMC) that differentiate into dentin-secreted odontoblasts [12]. Targeted mutations in transcriptional factor genes of NC cells results in craniofacial and tooth development abnormalities [13,14]. Therefore, NC cells are critical for tooth organogenesis. Recently, several reports have proposed effective induction protocols to promote the differentiation of ES cells into NC cells [15,16]. The induced NC cells generate neurons, Schwann cells, and mesenchymal cells as derivatives. Further, the mesenchymal cells differentiate into adipocytes, chondrocytes, osteoblasts, and smooth muscle cells. From these findings, we speculate that iPS cell-derived NC cells can be suitable cell sources for tooth regeneration.
In this study, we established a culture protocol to induce the differentiation of mouse iPS cells into NC-like cells (NCLC), and demonstrated that iPS cell-derived NCLC can differentiate into DMC and odontoblasts upon stimulation by dental epithelial cells.
Materials and Methods
Antibodies
Anti-nestin mouse monoclonal antibodies were purchased from Chemicon (Billerica, MA). Anti-p75NTR rabbit polyclonal antibodies, anti-Wnt-1 goat polyclonal antibodies, anti-Pax9 rat monoclonal antibodies, and anti-dentin sialoprotein (DSP) goat polyclonal antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Anti-STRO-1 mouse monoclonal antibodies were obtained from R&D Systems (Minneapolis, MN); and anti-AP2-α mouse monoclonal antibodies, from Cosmo Bio (Tokyo, Japan). Anti-HNK-1 mouse monoclonal antibodies came from Sigma-Aldrich (Tokyo, Japan). Anti-green fluorescent protein (GFP) monoclonal antibodies were a product of Cell Signaling Technology (Danvers, MA). Anti-Lhx6 rabbit polyclonal antibodies were purchased from Abgent (San Diego, CA); and anti-Msx1 rabbit polyclonal antibodies, from Lifespan Biosciences (Seattle, WA). Hoechst 33342, Alexa Fluor 488, and Alexa Fluor 546 secondary antibodies were procured from Invitrogen (Carlsbad, CA).
Cell culture and NC induction
Cells of the mouse iPS cell line iPS-MEF-Ng-20D-17 (RIKEN BRC, Tsukuba, Japan) [4] were cultured on mitotically inactivated mouse embryonic fibroblasts in Dulbecco's modified Eagle's medium (DMEM; Invitrogen) containing 15% fetal bovine serum, 0.1 mM nonessential amino acids (Invitrogen), 0.1 mM 2-mercaptoethanol (Sigma-Aldrich), 50 U/mL penicillin, 50 μg/mL streptomycin (Invitrogen), and 1,000 U/mL mouse leukemia inhibitory factor (Chemicon). The cultures were maintained at 37°C in a 5% CO2 humidified incubator, and the medium was changed every other day.
NCLC were derived from mouse iPS cells by a previously described method [17] with modifications. In brief, iPS cells were dissociated with 0.25% trypsin/EDTA and transferred to nonadherent culture dishes (HydroCell, CellSeed, Tokyo, Japan) with neural induction medium for suspension culture to form neuroectodermal spheres. The neural induction medium consisted of a 1:1 ratio of DMEM/F12 (Invitrogen) and neurobasal medium (Invitrogen) supplemented with 0.5× N2 (Invitrogen), 0.5× B27 (Invitrogen), 5 μg/mL insulin (Sigma-Aldrich), 20 ng/mL basic fibroblast growth factor (bFGF) (R&D Systems), 20 ng/mL epidermal growth factor (R&D Systems), 50 U/mL penicillin, and 50 μg/mL streptomycin. After 4 days in suspension culture, the spheres were transferred to fibronectin-coated culture dishes (BD Biosciences, San Jose, CA). After 8–10 days, the spheres or rosettes of NCLC were manually removed, and the resultant NCLC were dissociated with 0.25% trypsin/EDTA, and plated on fibronectin-coated culture dishes. The isolation of HNK-1-positive cells was carried out with a magnetically-activated cell sorting separator (Miltenyi Biotec, Bergisch-Gladbach, Germany) using magnetic microbeads (Miltenyi Biotec) coupled to anti-HNK-1 mouse monoclonal antibody.
Immunofluorescence
Immunofluorescence was performed according to our previously described method [18]. In brief, the cultured cells were fixed with 4% paraformaldehyde, rendered permeable in 0.1% Triton X-100 (Sigma-Aldrich), and blocked with 5% serum. The cells were then incubated with primary antibodies for 1 h at room temperature. Alexa Fluor 488- and Alexa Fluor 546-conjugated secondary antibodies were used for detection of the target. Secondary antibodies were added to frozen sections that had been incubated with primary antibodies overnight at room temperature. Fluorescent images were obtained by using a fluorescence microscope (IX71; Olympus, Tokyo, Japan). Images were analyzed with software supplied with the confocal microscope or standard image analysis software (Metamorph; Universal Imaging Corporation, Downingtown, PA). All imaging data were obtained from experiments replicated at least 4 times.
Flow cytometry
The cells were detached with 0.25% trypsin/EDTA. For cellular staining, ∼0.5–1×106 cells were incubated with the primary antibody on ice for 30 min. Phycoerythrin-conjugated secondary antibody was then added. Cells were analyzed with a flow cytometer (EPICS XL ADC; Beckman Coulter, Tokyo, Japan) and the supplied software. Experiments were performed in triplicate.
Real-time reverse transcription-polymerase chain reaction
Real-time reverse transcription (RT)-polymerase chain reaction (PCR) was performed according to our previously described method [18]. In brief, total RNA was isolated by using an RNeasy Mini Kit (Qiagen, Tokyo, Japan) according to the manufacturer's protocol. Reverse transcription of total RNA was performed by use of a PrimeScript RT reagent kit (Takara Bio, Siga, Japan). cDNA amplification was performed by real-time RT-PCR using SYBR® Premix Ex Taq™ (Takara Bio) with a Thermal Cycler Dice (Takara Bio) according to the manufacturer's protocol. Primer sequences for each cDNA were as follows: for GAPDH, 5′-GTC TCC TCT GACTTC AAC A-3′ (forward) and 5′-CAG GAA ATG AGC TTG ACA AA-3′ (reverse); for Pax3, 5′-AAC AAG CTG GAG CCA ATC AAC TG-3′ (forward) and 5′-CTG AGG TCT GTG GAC GGT GCT A-3′ (reverse); for Snail, 5′-TCT GAA GAT GCA CAT CCG AAG C-3′ (forward) and 5′-TTG CAGTGG GAG CAG GAG AAT-3′ (reverse); for Slug, 5′-GGC TGC TTCAAG GAC ACA TTA GAA C-3′ (forward) and 5′-GGT CTG CAG ATGTGC CCT CA-3′ (reverse); for Foxd3, 5′-CGG CGC CTG TGT TCT CAG TA-3′ (forward) and 5′-CTG CGC AGA GTG AAC CTT CAA A-3′ (reverse); for DSPP, 5′-GAG CAA CAC GGA TGG ATG ATT TC-3′ (forward) and 5′-CAC TCT TGT CAC GCA CAG CCT TA-3′ (reverse); and for DMP1, 5′-GAT GTC AGA GCA CAC ATG GTG AGA-3′ (forward) and 5′-CAG GTT GGT GAA CCA GAG CAT C-3′ (reverse). Target gene expression levels were normalized to those of GAPDH. Further, relative gene expression levels were calculated relative to levels in control cultures by using the comparative Ct (2−ΔΔCt) method. Experiments were performed in triplicate.
Teratoma formation
Teratoma formation was assessed according to our previously described method [19]. In brief, iPS cells (1×106 cells) were mixed with 500 μL of Cellmatrix type I-A (Nitta Gelatin, Osaka, Japan). The mixture was subcutaneously injected into the lower flank of age-matched male KSN/Slc nude mice (Japan SLC, Shizuoka, Japan) or SCID-CB17 mice (CLEA Japan, Tokyo, Japan). After 4 weeks, Angiosense 750 (Olympus) was injected via a tail vein to highlight perfused vessels within the teratoma. One day after the injection, the fluorescence intensities and images were obtained by using an IVIS imaging system (Xenogen, Alameda, CA). The teratomas were then surgically extracted, fixed in 4% paraformaldehyde, decalcified, dehydrated through an ethanol series, embedded in paraffin, and serially sectioned at a 6-μm thickness. Finally the sections were placed on slides and then stained with hematoxylin and eosin.
All experiments were performed in accordance with the Protocols for the Humane Treatment of Animals of Iwate Medical University.
Tissue recombination
Dental epithelium was dissected from the lower incisors of transgenic mice expressing GFP. The apical ends of the incisors were removed and incubated for 10 min at 37°C in a solution of 1% Dispase (Roche, Tokyo, Japan) in PBS (−) containing 20 U/mL DNase I (Invitrogen). After incubation, the epithelium was mechanically separated by using fine forceps and needles. The absence of mesenchymal cell contamination was confirmed by visual inspection under a stereomicroscope.
After NCLC had been detached by using 0.25% trypsin/EDTA, they were centrifuged. The cell pellet (0.3–0.5 μL) was seeded in a drop of 30 μL of polymerized Cellmatrix type I-A on transplant filters (0.1-μm pore size; Millipore, Billerica, MA). A piece of dental epithelium was then placed over the gel, and the recombinant explants were cultured by the Trowell technique [20]. After 2 weeks in culture, the explants were sequentially fixed with 4% paraformaldehyde, immersed in a series of graded sucrose solutions, embedded in Tissue-Tek OCT (Sakura-Finetek USA, Torrance, CA), and frozen.
Conditioned medium of dental epithelial cell cultures
For preparation of medium conditioned by dental epithelial cells, we used the dental epithelial cell line HAT-7 [21]. The cells were maintained in DMEM/F12 medium containing 10% fetal bovine serum, 50 U/mL penicillin, and 50 μg/mL streptomycin. The culture medium was replaced every 2 days until the cells had reached 80%–90% confluence. The supernatant was filtered and stored at −80°C before use.
Statistics
All data were reported as the means±SD. Statistical significance was assessed by using the 2-tailed Student's t-test for 2 groups or analysis of variance (Tukey's test) for >2 groups. P<0.05 was considered as statistically significant.
Results
Derivation of NCLC from mouse iPS cells
Since DMC are generated from cranial NC cells [22], we hypothesized that the induction of iPS cells to differentiate into NCLC could be an effective method to obtain DMC. Previous studies have shown that human ES cell (hESC)-derived neural rosettes can differentiate into NC cells [16,17]. Here we modified the differentiation protocol for mouse iPS cells. Mouse iPS cells were cultured in suspension for 4 days to form neural spheres (Fig. 1A). The neural spheres were then transferred to fibronectin-coated dishes, and they spontaneously attached to the dishes within 1 day (Fig. 1B). The spheres formed rosette-like structures and gave rise to migratory cells with a stellate morphology (Fig. 1B, C). The rosettes were then mechanically removed, and the remaining migratory cells formed a uniform cell population. These migratory cells were passaged and maintained on fibronectin-coated dishes for subsequent experiments (Fig. 1D). Expression of NC marker protein in the derived cells was assessed by immunofluorescence and flow cytometry. The immunofluorescence study showed that >90% of the cells were positive for nestin (91.9±4.1, n=4), AP2-α (90.2±3.4, n=4) p75NTR (90.9±2.4, n=4), and Wnt-1 (94.5±4.8, n=3), which are NC lineage-specific markers (Fig. 2A). They were also partially positive for the mesenchymal stem cell marker STRO-1 (62.5±2.8, n=4). In contrast, Nanog-GFP, which is expressed in undifferentiated iPS cells (data not shown), was not detected in the derived cells (Fig. 2A). Flow cytometry demonstrated that the number of HNK-1 (marker of migrating NC cells)- positive cells among the derived cells was increased compared with that among the undifferentiated iPS cells (Fig. 2B). Real-time RT-PCR further confirmed that mRNA expression of NC-specific transcriptional factors, such as Pax3, Snail, and Slug, was markedly higher in derived cells than in undifferentiated iPS cells (Fig. 2C); whereas Foxd3 was not detected (data not shown). On the basis of these observations, we termed these derived cells mouse iPS cell-derived NCLC. Further, we examined the expression of DMC markers Lhx6, Msx1, and Pax9, and the odontoblast marker DSP, [13,14,23,24], in the NCLC by immunofluorescence. Although Lhx6, Msx1, and Pax9 are expressed in cells other than dental mesenchyme, the combined expression of these genes is specific to DMC [25]. Lhx6 and Msx1 were detected in the NCLC, whereas Pax 9 and DSP were not (Fig. 3).

Derivation of NCLC from mouse iPS cells.
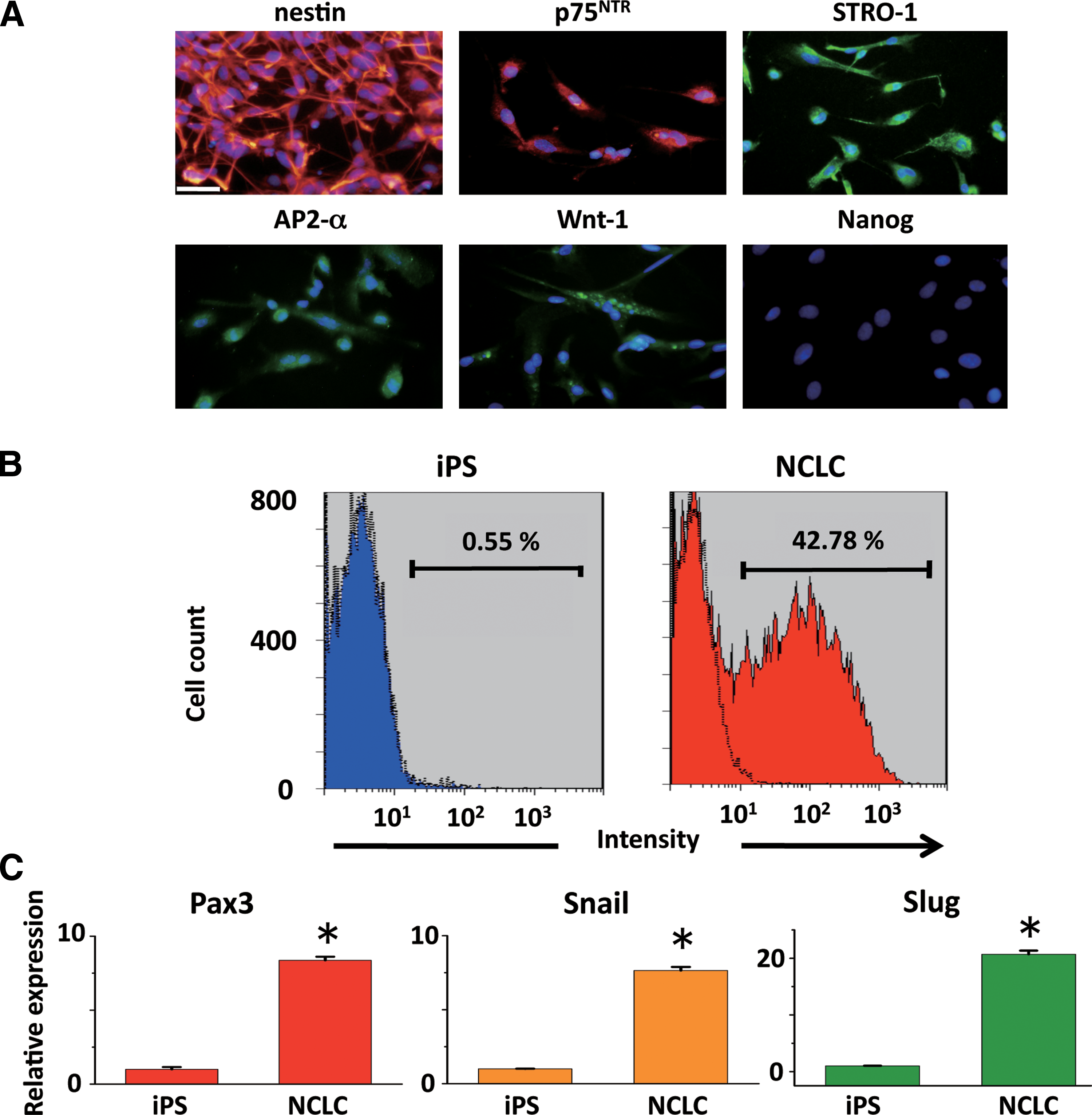
Characterization of NCLC induced from iPS cells.
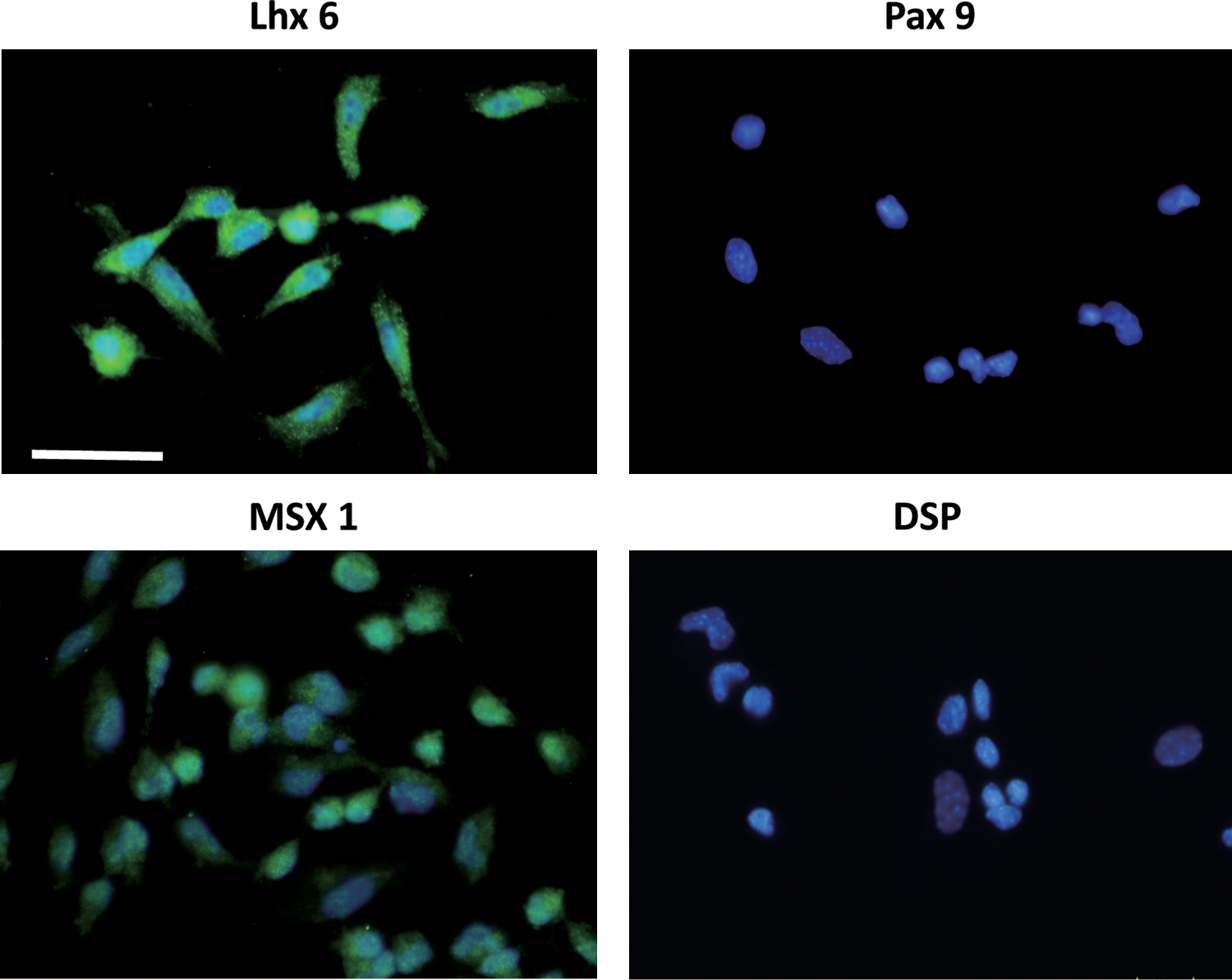
Expression of DMC and odontoblast marker proteins in NCLC. NCLC were immunostained with the indicated antibodies. Nuclei were stained with Hoechst 33342 (blue). Scale bar=50 μm. DMC, dental mesenchymal cells. Color images available online at
Further, to evaluate their tumorigenicity we subcutaneously injected NCLC into immunodeficient mice. After 4 weeks, these transplanted NCLC did not form visible teratomas (Fig. 4A), as expected from the absence of Nanog expression; whereas parental undifferentiated iPS formed teratomas including those containing various types of tissues such as bone, cartilage, epidermis, muscle, adipose tissue, and gut-like epithelium (Fig. 4A, B). To further confirm that the NCLC did not form tumors, we performed in vivo imaging to assess tumor angiogenesis. The mice injected with undifferentiated iPS cells showed significant fluorescence of the blood pool-imaging agent at the teratoma site, whereas the NCLC-injected ones showed no imaging signals, indicating the absence of a tumor.
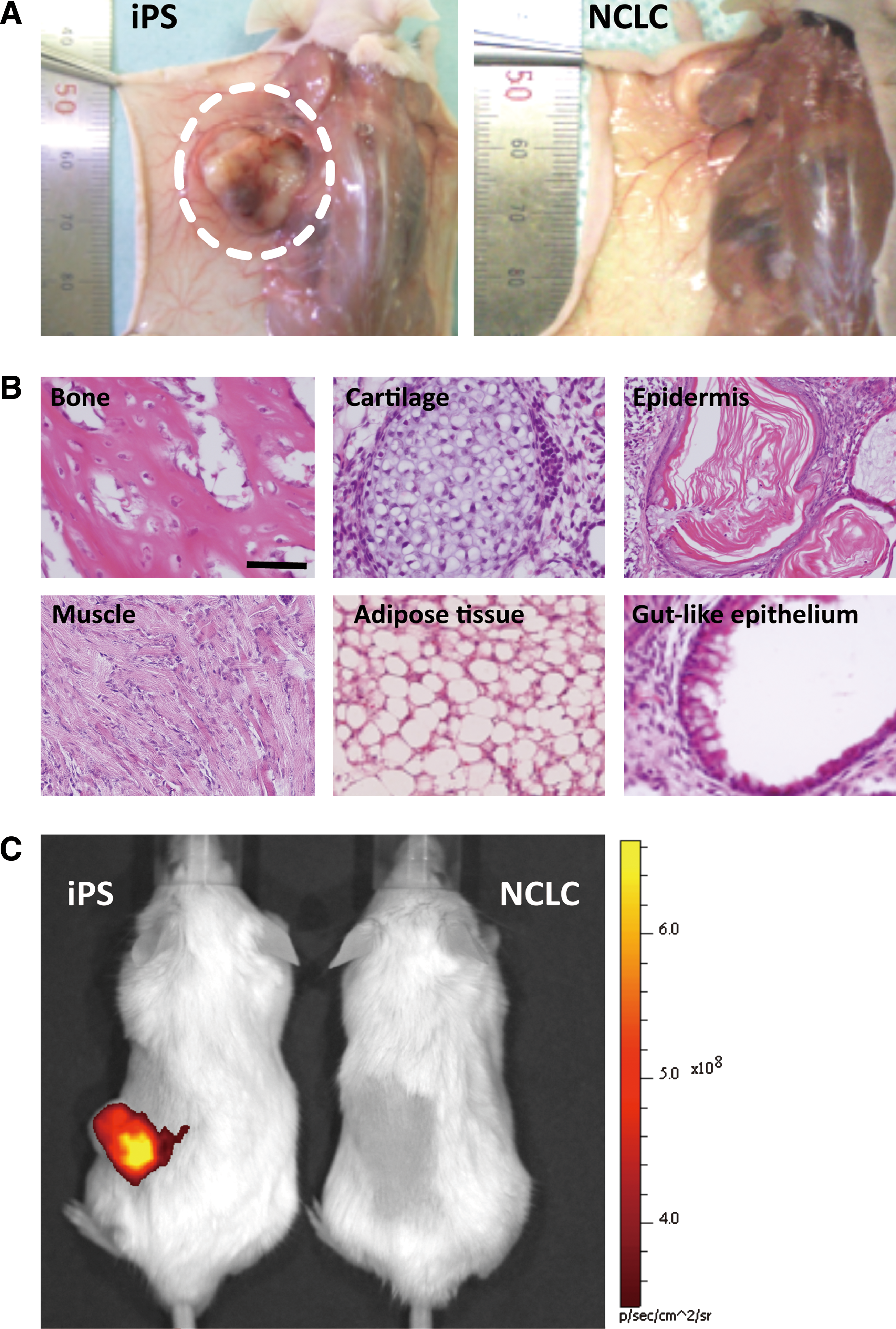
Teratoma formation of NCLC.
Together, these results suggest that we could obtain nontumorigenic NCLC efficiently from mouse iPS cells by using this protocol.
Odontogenic response of iPS cell-derived NCLC
To determine whether the NCLC had the capacity to differentiate into DMC, we prepared combination cultures of NCLC and mouse dental epithelium taken from the apical ends of the lower incisors. The combined cells were cultured in vitro and analyzed for the expression of molecular markers of tooth development. After 2 weeks in culture, NCLC aggregation adjacent to the dental epithelium was observed (Fig. 5A). Pax9, Msx1, and Lhx6 were expressed in the NCLC but not in the GFP-expressing dental epithelium (Fig. 5B–D), and Pax9 was co-expressed with Msx1 and Lhx6 (Fig. 5E–J), suggesting the differentiation of NCLC into DMC.
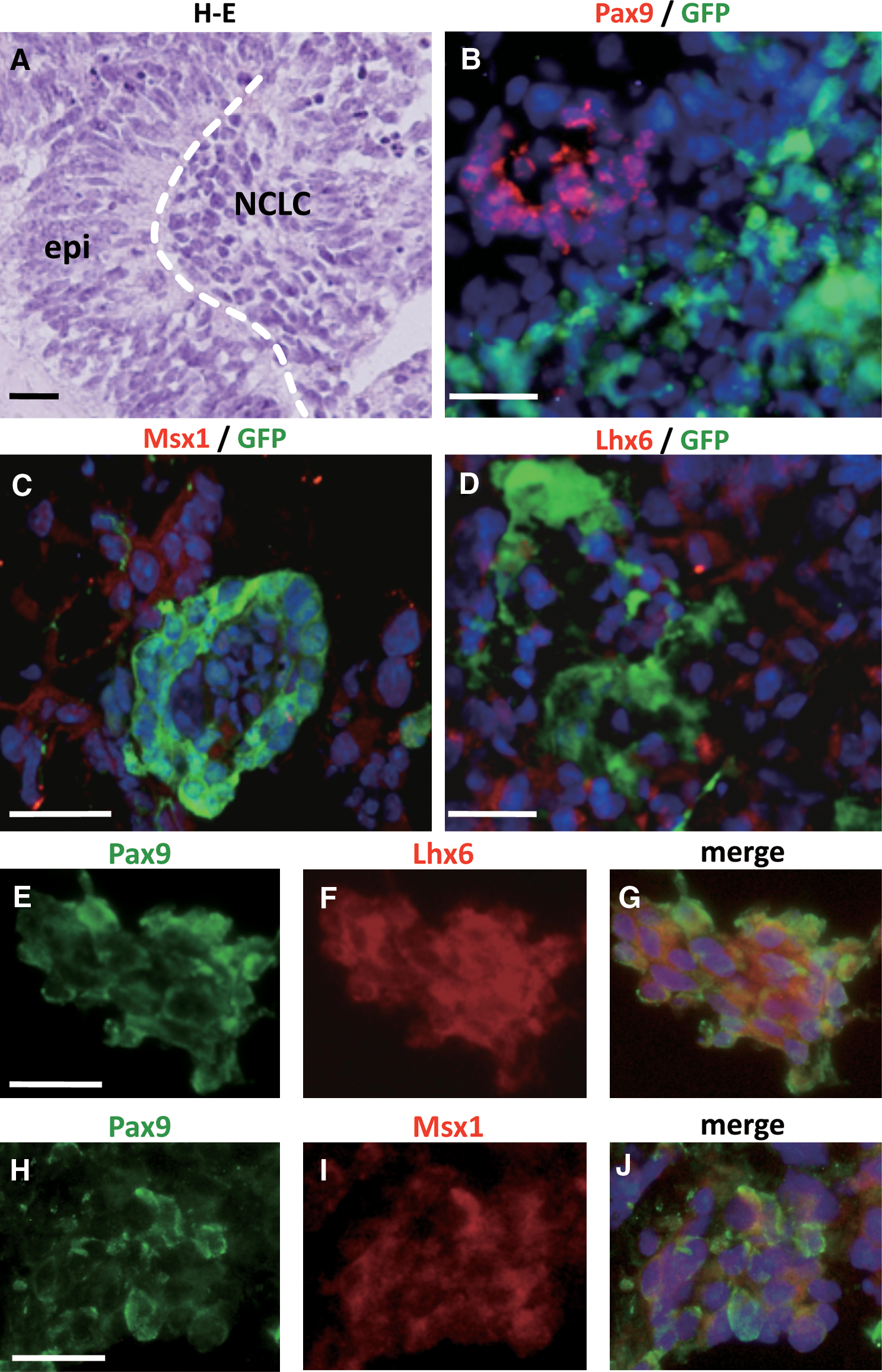
Combination cultures of NCLC and dental epithelium.
Interestingly, some NCLC also expressed the odontoblast marker DSP (Fig. 6A). In contrast, no marker expression was observed in combination cultures of undifferentiated iPS cells and dental epithelium (data not shown). Additionally, when NCLC were cultured in the presence of serum, dentin sialophosphoprotein (DSPP) and dentin morphological protein 1 (DMP1) mRNA expression increased; and this increase was further enhanced with the conditioned medium (CM) of dental epithelial cell cultures (Fig. 6B, C). Moreover, the HNK-1-positive cells isolated from the NCLC population presented higher expression of DSPP and DMP1 than the HNK-1-negative cells when cultured with serum or with CM from dental epithelial cell cultures (Fig. 6D, E). Taken together, these results indicate that iPS cell-derived NCLC, especially the HNK-1 positive cells, had the potential to differentiate into odontoblast progenitor cells upon stimulation with dental epithelium.

Odontogenic response of NCLC to dental epithelium.
Discussion
In this study, we succeeded in inducing the differentiation of mouse iPS cells into NCLC in vitro, and demonstrated for the first time that NCLC could further differentiate into odontogenic mesenchymal cells, including odontoblasts progenitor cells. We modified a culture protocol for the differentiation of hESCs [17] into NC cells, and showed that it was suitable for use with mouse iPS cells, though the species (murine and primate) and cell types (iPS and ES) were different. With regard to species, the signaling pathways that regulate primate ES cell differentiation are similar to those operating in murine ES cells [25]. Therefore, this confirms the success of transferring differentiation strategies from the primate to the murine system. In fact, the method for inducing differentiation of murine ES cells into neural cells functions with primate ES cells as well [26,27]. Concerning cell types, iPS cells are virtually equivalent to ES cells in terms of pluripotency, ES cell marker expression, and teratoma formation in vivo [4 –6]. Previous reports demonstrated that iPS cells differentiate into various cell types, including NC cells, under the same culture conditions as used for ES cells [7 –9]. However, some reports documented that the differentiation capability of iPS cells is lower than that of ES cells [8,9]. Hence, further improvements are needed for enhancing iPS cell differentiation capacity.
Our iPS cell-derived NCLC expressed NC markers such as nestin, p75NTR, AP2-α, and Wnt-1 (Fig. 2). However, interestingly, Foxd3 was not expressed. In Mundell's report, Foxd3 is downregulated in cranial NC mesenchyme, and Foxd3-null cranial NC cells show accelerated differentiation into mesenchymal cells [28]. Using our differentiation protocol, we found that NCLC expressed the mesenchymal stem cell markers STRO-1, Lhx6, and Msx1, which are expressed in dental mesenchyme. Hence, NCLC may have preferentially differentiated into mesenchymal cells. In addition to Lhx6 and Msx1 expression, NCLC expressed Pax9 (Fig. 5), when cultured with dental epithelium. This result is consistent with Ohazama's report showing that ES cells, neural stem cells, and adult bone marrow-derived cells express Lhx6, Msx1, and Pax9 when cultured with embryonic oral epithelium [29]. Thus, similar to ES and other stem cells, our iPS cell-derived NCLC showed the capacity to differentiate into dental mesenchyme upon stimulation with dental epithelium.
We further showed that NCLC in combined culture with dental epithelium differentiated into DSP-expressing cells, indicating differentiation into odontoblasts (Fig. 6A). In this experiment, we detected DSP expression in NCLC around the dental epithelium. In addition, the serum and CM from dental epithelial cell cultures enhanced DSPP and DMP1 expression (Fig. 6B–E). These results suggest that the serum and soluble components from dental epithelial cells provided NCLC with an environment suitable for odontoblast differentiation. Since NCLC derived from hESCs differentiate into several mesenchymal lineages in the presence of serum [15], our iPS cells-derived NCLC may have been induced to differentiate into DMC lineages in the presence of serum and further to differentiate into odontoblasts in the presence of soluble components of the dental epithelium. Medium conditioned by tooth germs or ameloblasts has the potential to induce stem cells to differentiate into odontogenic cells [1,30]. Thus, CM, including ours, may contain important signal molecules for tooth development, such as Notch-1 and FGFs [20]. Further investigation is needed to identify the factors that promote effective odontoblast induction. Since the detailed mechanism of the spatio-temporal regulation of odontoblast differentiation is still unclear, our culture protocol will be most useful for studying odontoblast differentiation. In addition to odontoblasts, dental pulp cells and dental follicle cells are also believed to arise from DMC during tooth germ development. Although not addressed here, it would be interesting to know whether NCLC have the capability to differentiate into those 2 cell types.
Recent advances in tissue engineering techniques indicate that the bioengineering approach may be successful for the regeneration of dental tissue. The main concept of tooth regeneration is to mimic the process of natural tooth development, either in vitro or in vivo. By taking advantage of reciprocal epithelial–mesenchymal interactions, numerous studies have shown that dental epithelial and mesenchymal cells in fetal tooth germs can form bioengineered teeth [31 –33]. However, regarding clinical applications, the use of fetal tissue and/or cells gives rise to the same ethical issues as those encountered with ES cells. Therefore, iPS cell-derived DMC generated in this study have the potential to overcome the problems related to cell sources for stem cell-based tooth regeneration and treatment of tooth-related diseases.
Footnotes
Acknowledgments
This work was supported, in part, by the Iwate Medical University Open Research Project (2007–2011; to K.O., N.F., and H.H.), by grants from the programs Grants-in-Aid for Scientific Research (C; No. 19562128 to N.F.) and Grants-in-Aid for Young Scientists (C; No. 20679006 to S.F.) from MEXT, and by the Next Program LS010 (to S.F.)
Author Disclosure Statement
No competing financial interests exist.