
Abstract
Neural stem cell-based approaches to repair damaged white matter in the central nervous system have shown great promise; however, the optimal cell population to employ in these therapies remains undetermined. A default mechanism of neural induction may function during development, and in embryonic stem cells (ESCs) neural differentiation is elicited in the absence of any extrinsic signaling in minimal, serum-free culture conditions. The default mechanism can be used to derive clonal neurosphere-forming populations of neural stem cells that have been termed leukemia inhibitory factor-dependent primitive neural stem cells (pNSCs), which subsequently give rise to fibroblast growth factor 2-dependent definitive NSCs (dNSCs). Here we characterized the neural differentiation pattern of these two cell types in vitro and in vivo when transplanted into the dysmyelinated spinal cords of shiverer mice. We compared the differentiation pattern to that observed for neural stem/progenitor cells derived from the adult forebrain subependymal zone [adult neural precursor cells (aNPCs)]. dNSCs produced a differentiation pattern similar to that of aNPCs in vitro and in the shiverer model in vivo, where both cell types produced terminally differentiated oligodendrocytes that associated with host axons and expressed myelin basic protein. This is the first demonstration of the in vivo differentiation of NSCs, derived from ESCs through the default mechanism, into the oligodendrocyte lineage. We conclude that dNSCs derived through the default pathway of neural induction are a similar cell population to aNPCs and that the default mechanism is a promising approach to generate NSCs from pluripotent cell populations for use in cell therapy or other research applications.
Introduction
C
Traditional techniques to derive NSCs and their derivatives from pluripotent cell populations involve the induction of embryoid body (EB) formation [10] by the withdrawal of growth factors promoting pluripotency, followed by the selection and subsequent enrichment of ectodermal and ultimately neural cells. The most common techniques have been based on EB formation followed by retinoic acid treatment [3,11]. Less common methods involve co-culture with stromal cell populations with neural inducing properties [12]. Despite the success of these approaches, the involvement of EB formation creates a risk for the presence of non-neural lineage cells in later cell populations. This can lead to tumorigenic potential, as has been observed in oligodendrocyte progenitor derived from murine embryonic stem cells (ESCs) by EB formation followed by treatment with retinoic acid, noggin, fibroblast growth factor (FGF2), and platelet-derived growth factor (PDGF) [13]. The maintenance of non-neural lineage cells in these cultures can likely be traced to the nonspecific differentiation pattern initiated in the process of cell aggregation leading to EB formation. To address this concern, we used the default pathway of neural induction [14 –16] to produce clonal NSC populations directly from ESCs without EB formation.
Our method is based on an intrinsic property of ESCs, whereby the neural lineage is the default pathway of ESC differentiation in the absence of other extrinsic signaling (Fig. 1A) [14,15,17]. This default mechanism has been shown to act in vivo as well as functioning in vitro in ESCs (for review see [16]). The default model of neural induction is based on findings that neural lineage commitment is induced through the inhibition of bone morphogenetic protein signaling through both protein inhibitors and/or modified gene expression and, in the absence of any instructive signaling, ectodermal cells adopt a neural fate [16,18]. Recent findings suggest an important role for the zinc finger protein Zfp521 in this process [19]. When cultured in low density minimal conditions, ESCs rapidly acquire a default neural identity that is seemingly similar to adult neural stem or progenitor cells, as judged by the expression of the neural markers nestin and Sox1 by over 99% of surviving cells after 24 h [14]. Using a limiting dilution cell assay [15], similar to that used to generate neurosphere colonies of adult neural precursor cells (aNPCs) from the adult brain, it has been demonstrated that a small percentage (∼0.2%) of ESCs possess the ability to generate NSC colonies when cultured at low density, in defined serum-free media (SFM), and in the presence of leukemia inhibitory factor (LIF). This cell population was termed the primitive NSC (pNSC). While evidence for the adoption of a neural fate is strong when culture conditions are extremely minimal, when pNSCs are cultured to produce neurospheres in the presence of LIF, the commitment to the neural lineage is incomplete. Cells are found to still express the pluripotency marker octamer-binding transcription factor 4 (Oct4) and possess multi-germline potential in mouse embryo chimera experiments [15]. However, when the primary neurosphere colonies are passaged, they develop dependence on exogenous basic FGF2, similar to that observed in aNPCs; they no longer express Oct4; and they lose the ability to incorporate into the developing blastocyst in chimera experiments. These secondary neurosphere colonies have been named definitive NSCs (dNSCs) [14,15]. Here, we hypothesized that the generation of dNSCs through the default pathway would act as a useful method to generate NSCs from a pluripotent cell population with a therapeutic potential similar to that of aNPCs for cell-based remyelinating therapies. In this study we performed gene expression profiling of pNSC, dNSC, and aNPC neurosphere colonies, and characterized the neural differentiation pattern of NSC populations that were derived through the default pathway both in vitro and in vivo, after transplantation into the dysmyelinated spinal cords of shiverer mice. We compared the differentiation pattern of the pNSCs and dNSCs to that of aNPCs. The results suggest that the production of NSC from ESCs, through the default pathway, is a promising method to produce neural lineage cells for the development of cell-based remyelinating therapies or other research purposes.
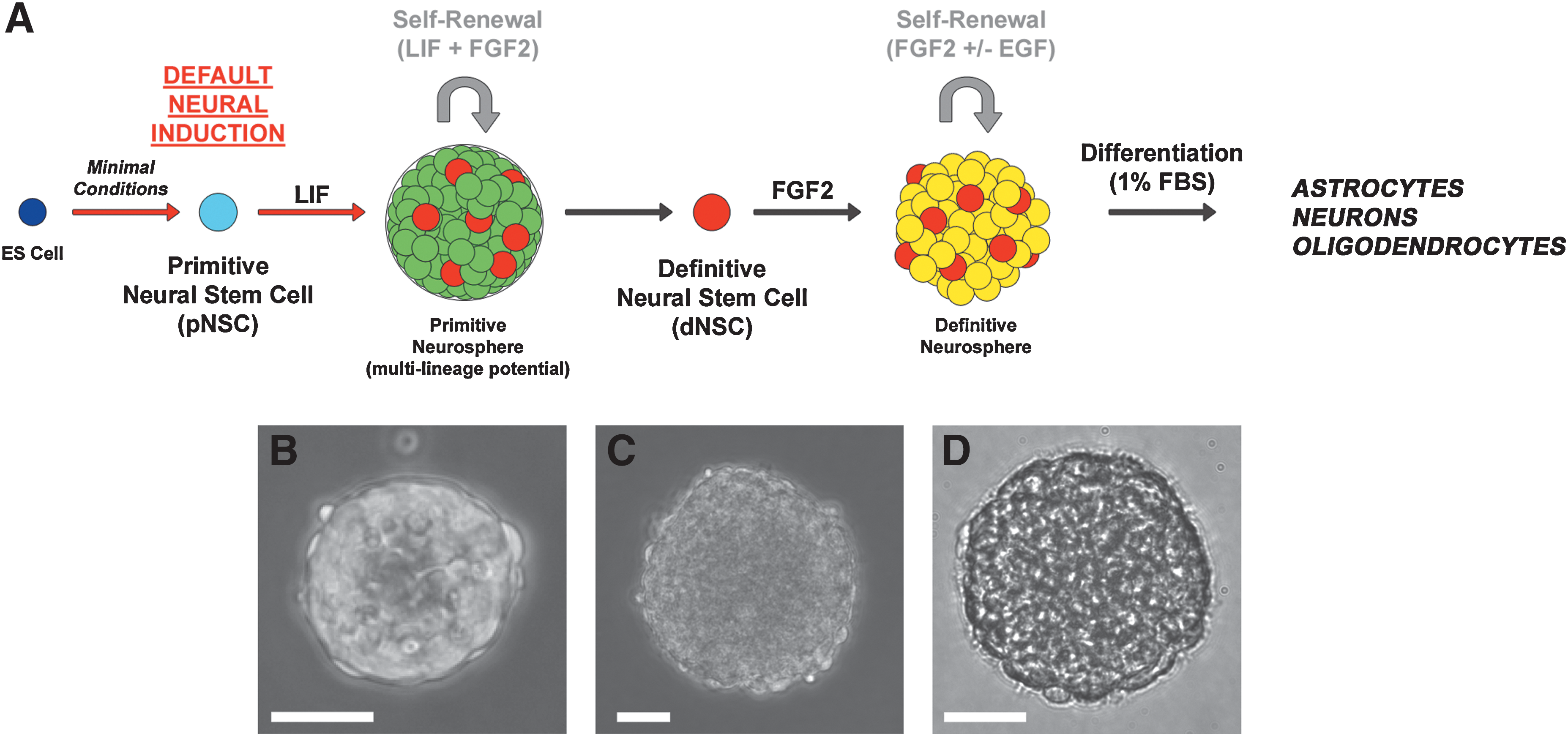
Default neural differentiation of embryonic stem cells.
Materials and Methods
All experimental animal procedures were approved by the animal care committee of the University Health Network and are in accordance with the policies and guidelines of the Canadian Council on Animal Care.
ESC culture and production of ES-derived neurospheres through the default mechanism
All ES-derived neurospheres colonies were produced using previously reported techniques [14,15,20]. The ESC line used was the YC5 variant of the R1 mouse ESC line [15] that expresses yellow fluorescent protein (YFP). To produce primary neurospheres generated by pNSCs, ES colonies grown under standard ES conditions on feeder cells were dissociated using Trypsin–ethylenediaminetetra-acetic acid (Sigma) and seeded in uncoated plastic culture flasks into chemically defined SFM [21] containing LIF (1,000 U/mL) at a density of 10 cells/μL. At this cell density, the primary spheres produced have been previously demonstrated to be clonally derived [22]. Primary spheres were present after 7 days, passaged as described above, and plated at 10 cells/μL into SFM containing LIF (Millipore), FGF2 (10 μg/100 mL; Sigma), and Heparin (2 μg/100 mL; Sigma). Secondary spheres were present after 7 days and could be passaged to derive tertiary sphere populations. Definitive neurospheres, derived from dNSCs, were produced by the passage of single cell dissociated LIF-dependent pNSC spheres, at passage 3, into SFM containing FGF2 and Heparin, but not LIF. B27 nutrient supplement (Gibco) was added to enhance cell survival in the absence of LIF [14] (Fig. 1A).
Adult NSC isolation and culture
aNPCs were obtained from the brains of transgenic mice expressing enhanced YFP (EYFP) [129-Tg (ACTB-EYFP) 2Nagy/J; The Jackson Laboratory]. Cells were harvested from the forebrain subependymal zone using previously described methods [1,23,24]. Primary neurospheres were present after approximately 7 days after initial isolation and passaged weekly by mechanical dissociation (no trypsin) and were seeded into SFM containing FGF2, epidermal growth factor (EGF), and heparin at a density of 10–20 cells/μL.
In vitro cell differentiation and immunocytochemistry
For the in vitro analysis of cell fate potential, neurospheres were differentiated by removing growth factors (LIF, FGF2, Heparin, and EGF) from the media and adding 1% fetal bovine serum, as previously reported [1]. All differentiation experiments were conducted using neurospheres between passages 3 and 5. The spheres were dissociated as described and seeded onto Matrigel (Sigma)-coated, multi-well glass slides (Thermo Scientific/Nunc). Wells were seeded at 5,000 cells per well for pNSCs and 15,000 cells per well for dNSCs and aNPCs. Cells were allowed to differentiate for 7 days before being fixed for immunocytochemical analysis with 4% paraformaldehyde (PFA) at room temperature. Nonspecific binding of antibodies was blocked by incubation for 1 h with phosphate-buffered saline (PBS) containing 1% bovine serum albumin (BSA), 10% normal goat serum (NGS), and 0.25% Triton X-100. Cultures were treated with primary antibodies (Table 1) for 2 h at room temperature and washed 3 times before incubation with appropriate fluorescently conjugated secondary antibodies for 1 h. Nuclei were counterstained by the use of VECTASHIELD mounting medium with 4′,6-diamidino-2-phenylindole (DAPI; Vector Labs).
APC, adenomatous polyposis coli; GFAP, glial fibrillary acidic protein; MBP, myelin basic protein; PDGFRα, platelet-derived growth factor alpha; NSC, neural stem cells; NPCs, neural precursor cells.
Real-time PCR
Real-time reverse transcription–polymerase chain reaction (RT–PCR) analysis was conducted using TaqMan-based preloaded microfluidic array cards and the ABI PRISM® 7900HT Sequence Detection Systems (Applied Biosystems) to generate a limited gene expression profile for neurospheres produced by each NSC type. TaqMan cards were used according to the manufacturers' instructions and all PCR assays were performed in triplicate. RNA was isolated using Trizol for initial cell disruption, followed by the commercially available RNeasy Micro kit (Qiagen). RNA was isolated from intact sphere colonies at passage 3. Gene expression in the ESC line, used to produce the pNSC and dNSC spheres, was used at the baseline expression level for calculating relative expression values. The expression was normalized to glyceraldehyde 3-phosphate dehydrogenase. ES colonies used for RNA extraction were grown on matrigel-coated dishes in embryonic fibroblast-conditioned ES media, instead of on feeder cells. Gene expression levels were compared using the 2−ΔΔCT method [25] using the Applied Biosystems SDS software package. The genes used to compare expression profiles of the cell types were as follows: Pluripotency (Oct4, nanog, and Sox 2), cell differentiation [teratocarcinoma-derived growth factor 1 (TDGF1), DNM3TB, gamma-aminobutyric acid (GABA) A receptor, beta 3 (GABRB3), growth differentiation factor-3 (GDF3), Lefty, Nodal, and semaphorin 3a (Sema3a)], and neural lineage specification [paired box gene 6 (Pax6), Nestin, oligodendrocyte transcription factor 2 (Olig2), glial fibrillary acidic protein (GFAP), and synaptophysin]. TaqMan primer sequence information is proprietary (
Animal care and surgical procedures
Adult (6–8 weeks old) shiverer mice (shi/shi) (C3Fe.SWV-Mbp-Shi/J; The Jackson Laboratory) were used for transplantation experiments. The shiverer mouse has a deletion in the myelin basic protein (MBP) gene resulting in poorly compacted and sparse CNS myelin [26]. The shiverer mouse lacks the endogenous capacity for remyelination and is well suited for evaluating the myelinating potential of transplanted cell types [7,27]. To characterize the neural differentiation pattern of the NSC populations in vivo, we transplanted the cell types into the spinal cords of shiverer mice. Before surgery, mice were immunosuppressed by the administration of cyclosporine A (20 mg/kg, 10 mg/kg after 14 days) in their drinking water 48 h before receiving transplants and continuing until the animals were sacrificed. Mice were also treated with Minocycline (50 mg/kg) beginning 48 h before surgery for 10 days for both its immunosuppressive properties, the inhibition of microglial activation [28], and its modest neuroprotective properties [29,30] to limit potential injury due to the injection procedure. Under isofluorane anesthesia (1%–2% with 1:1 mixture of O2/N2O) a T6 laminectomy was performed to expose the spinal cord, and cells were injected using a pulled glass micropipette attached to a 5 μL Hamilton syringe. Cells were transplanted bilaterally adjacent to the midline dorsal vein at a depth of 0.5 mm. All cell types were prepared for transplantation by dissociation and resuspension at a concentration of 50,000 cells/μL in SFM. The injection volume was 2 μL per site and cells were injected at a rate of 0.5 μL/min by a computer-controlled syringe pump (World Precision Instruments). The cell injection procedure was well tolerated and did not result in any detectable locomotor or behavioral deficits in the mice. No behavioral assessment was conducted on transplanted animals. For this study the shiverer model was employed as a means to directly compare the neural differentiation pattern of dNSCs and aNPCs and not as a method to assess or compare therapeutic potential.
Immunohistochemistry and histology
Six weeks after cell transplantation, animals were administered an overdose of sodium pentobarbital (Somnotol) and transcardially perfused with ice-cold PBS followed by 4% PFA in PBS (pH 7.4). A 1 cm section of spinal cord centered on the injection site was removed and 14-μm-thick longitudinal spinal cord cryostat sections were prepared. Before staining, nonspecific antibody binding was blocked by incubation in PBS containing 1% BSA, 5% NGS, 5% Nonfat milk powder, and 0.25% Triton X-100 for 1 h at room temperature. Primary antibodies (Table 1) were applied in the same solution overnight at 4°C and slides were washed before application of the appropriate fluorescently conjugated secondary antibodies. Nuclei were counterstained by the use of VECTASHIELD mounting medium with DAPI (Vector Labs).
Cell counting and imaging methods
Quantification of in vitro differentiation was conducted by cell counts to determine the proportion of cells expressing a given cell marker. For each lineage marker, 12 separate wells were quantified for each primary antibody with 3 separate cell differentiation experiments conducted from distinct starting ESC populations. Using a Nikon C1 confocal microscope, 9 nonoverlapping images were collected in each culture slide well using a 20× objective. For each well, the number of positively stained cells and the total number of cells present were counted by DAPI-stained nuclei, to determine the proportion of cells expressing a given marker. Cells were counted using ImageJ software (
Statistical analysis
Cell count data are presented using descriptive statistics. The differences between groups were analyzed using z-tests for comparing proportions with the Bonferroni correction applied for multiple comparisons. Real-time quantitative PCR data was presented as a log fold expression change relative to the control. The differences between groups were analyzed using one-way ANOVA and the post hoc Student's t-test, and P values of <0.05 were considered statistically significant. Statistical analysis was performed using Microsoft Excel. Data are presented as means±standard error of the mean or as proportions±standard error of the proportion.
Results
Production of clonal neurospheres from ESCs through the default pathway
The dissociation of ES colonies into a single-cell solution under colony-forming assay conditions (10 cells/μL) in defined SFM containing LIF led to the formation of floating spherical colonies of cells, termed primitive neurospheres and derived clonally from ESCs as previously reported [14,15]. Primitive neurospheres appeared as dense cellular accumulations with a smooth outer surface when observed under a phase-contrast microscope. Primitive neurospheres could be passaged into SFM containing FGF2 and heparin (no LIF) to produce definitive neurospheres, clonally derived from dNSCs. Neurospheres derived from dNSCSs could also be passaged repeatedly in agreement with previous findings [14,15] (Fig. 1A). The gross morphology of definitive, dNSC-derived neurospheres was distinct from primitive neurospheres in that they did not have a smooth outer surface and individual cell profiles could be seen. Definitive neurospheres appear similar in morphology to adult brain-derived neurospheres colonies (Fig. 1B–D).
In vitro comparative gene expression profiling
The genes examined fell into 3 broad categories. (1) Genes involved in the maintenance of pluripotency [Oct4, nanog, and SRY (sex determining region Y)-box 2 (Sox2)], (2) genes involved in maintenance of an undifferentiated cell state (TDGF1, DNM3TB, GABRB3, GDF3, and Sema3a), and (3) genes involved in neural differentiation (Pax6, Nestin, Olig2, and synaptophysin). The expression level of the genes in undifferentiated ESCs was used as the baseline for calculating relative expression levels. We hypothesized that the expression pattern observed in pNSCs would be somewhat similar to that seen in ESCs, whereas the dNSCs would show a pattern of expression more similar to aNPCs. For the pluripotency genes, we observed a downregulation of expression of Oct4, nanog, and Sox2 in pNSCs versus ESCs and a downregulation of the genes in dNSCs and aNPCs for nanog and Oct4. Sox2 was expressed at similar levels in all cell types likely due to its role in both pluripotency [31] and neural stem cells [32] (Fig. 2A). For genes related to maintenance of an undifferentiated cell state, we observed a similar pattern of expression for all the genes in dNSCs and aNPCs, where again the pNSCs were more similar to ESCs (Fig. 2B). For the genes related to neural differentiation, we found a similar level expression in dNSCs and aNPCs for all 4 genes examined. Olig2 was the only neural gene that was upregulated in pNSCs versus the ESC control (Fig. 2C). Based on the gene expression patterns observed, the default-derived dNSC and aNPCs had a similar expression profile, whereas the pNSC had a profile that was intermediate between ESCs and more committed NSCs.
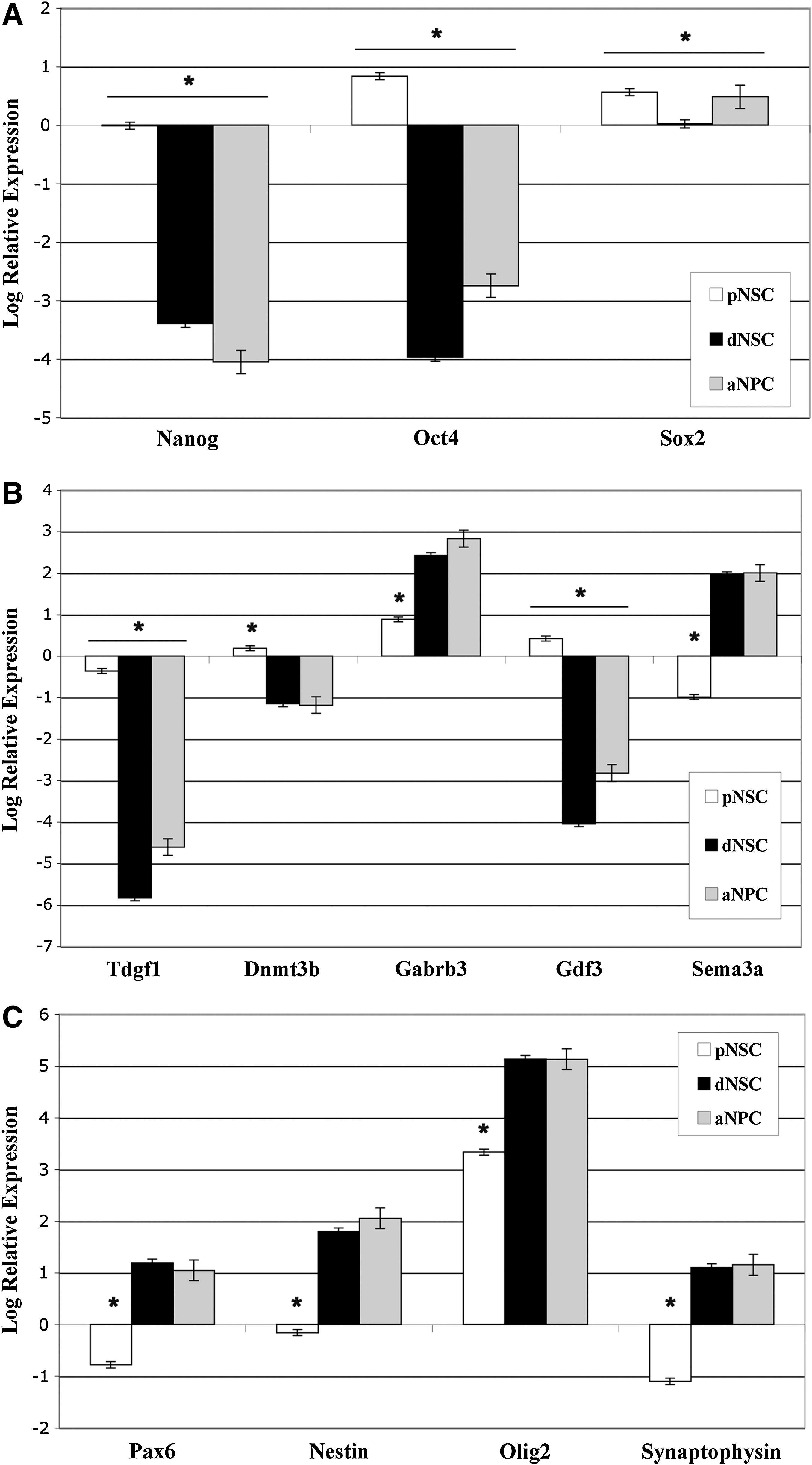
Gene expression relative to undifferentiated ES colonies in neurospheres generated from pNSCs, dNSCs, and aNPCs. Relative quantification by real-time reverse transcription–polymerase chain reaction analysis of expression of selected genes.
Characterization of neural differentiation patterns in vitro
The differentiation of aNPCs resulted in the production of all 3 cell types of the neural lineage—neurons, astrocytes, and oligodendrocytes—as previously reported [1,21,24]. Astrocyte lineage cells, as indicated by expression of GFAP, were the major cell type observed with 65%±1.5% of cells staining positively for GFAP. Oligodendrocytes accounted for 18%±1.2% of the cells as indicated by expression of Olig2, which is expressed by cells in all stages of the oligodendrocyte lineage. PDGFRα was expressed by 13%±1.1% of cells identifying them as oligodendrocyte progenitors or precursor cells [33,34]. Neurons represented 7%±0.8% of differentiated aNPCs as indicated by their expression of the neuronal marker beta-III tubulin. Approximately 1%±0.3% of aNPCs continued to express the neural stem/progenitor cell marker nestin (Fig. 3 and Supplementary Fig. S1; Supplementary Data are available online at
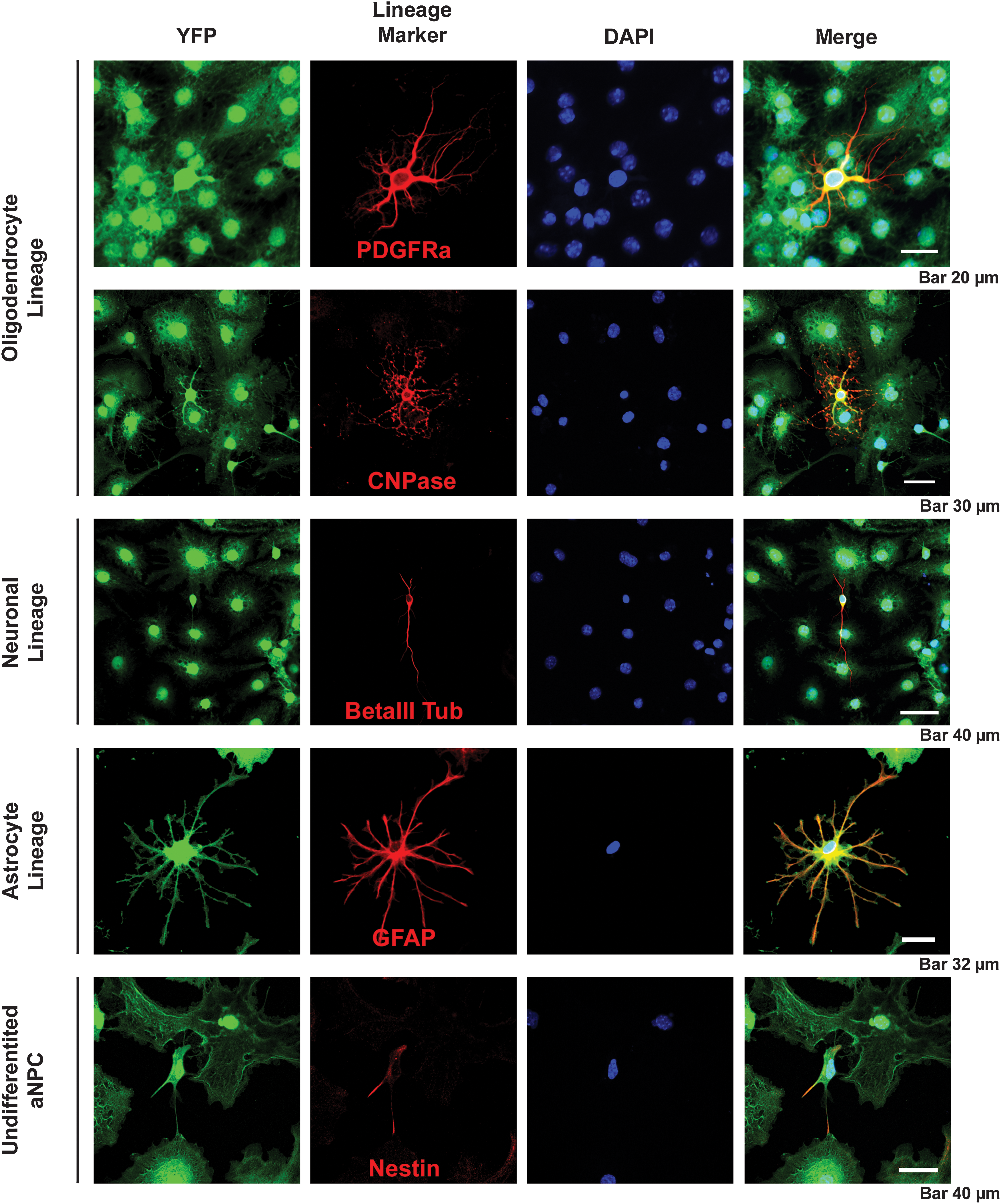
Characterizing the pattern of in vitro neural differentiation of aNPCs. All 3 neural linage cell types are produced by aNPC-generated neurosphere colonies when differentiated for 1 week in 1% FBS. Lineages were identified using immunocytochemical identification of the following lineage antigens: PDGFRα and CNPase for the oligodendrocyte lineage, βIII tubulin for the neuronal lineage, GFAP for the astrocyte lineage, and nestin expression was used to identify undifferentiated aNPCs. FBS, fetal bovine serum; PDGFRα, platelet-derived growth factor alpha; GFAP, glial fibrillary acidic protein.
Based on previous findings [14] and the similar gene expression pattern we observed in dNSCs and aNPCs, we hypothesized that the differentiation of dNSC neurospheres colonies would produce a similar pattern as that observed for aNPCs for the panel of neural antigens. The observed pattern contained all 3 neural lineages, and astrocytes accounted for the majority of cells observed with 61%±1.5% of cells expressing GFAP. In contrast to aNPCs, differentiated dNSCs produced more neurons than oligodendrocytes with 11%±1.0% of cells expressing beta III-tubulin, 6.1%±0.7% expressing Olig2, and only 2%±0.5% expressing PDGFRα. Nestin expression was maintained in 9.5%±0.9% of dNSCs after 1 week in differentiating conditions (Fig. 4 and Supplementary Fig. S2). Based on these data, the total percentage of differentiated dNSCs defined by our assay was 89%, leaving 11% of the cells undefined. While GFAP-expressing cells were the majority of the cells produced by both aNPCs and dNSCs in vitro, we found that the dNSCs produced a significantly higher proportion (2-tail P<0.05) of presumed neuronal lineage cells, as judged by beta III-tubulin expression, and a significantly lower proportion of Olig2-positive cells relative to aNPCs. The subset of the Olig2-expressing cell population that expresses the oligodendrocyte progenitor marker PDGRFα was also significantly lower in the dNSC group versus the aNPC (Fig. 5). dNSCs produced mature oligodendrocytes; some cultures contained CNPase-expressing cells with the morphology of fully matured in vitro oligodendrocytes [33] (Fig. 6).
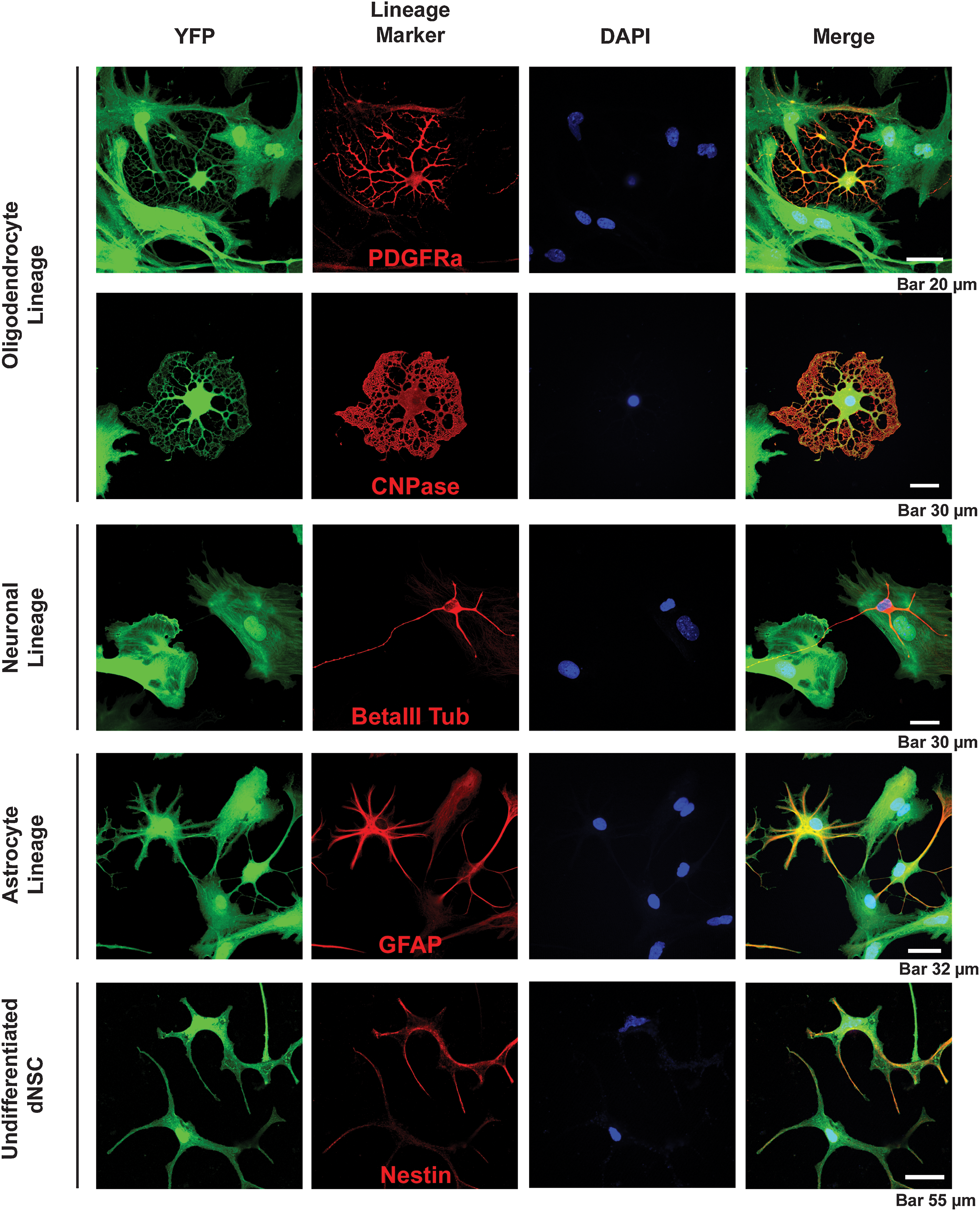
Characterizing the pattern of in vitro neural differentiation of dNSCs. All 3 neural linage cell types are produced by dNSC-generated neurosphere colonies when differentiated for 1 week in 1% FBS. Lineages were identified using immunocytochemical identification of the following lineage antigens: PDGFRα and CNPase for oligodendrocyte lineage cells, βIII tubulin for neuronal lineage, GFAP for the astrocyte lineage, and nestin expression was used to identify undifferentiated dNSCs. DAPI, 4′,6-diamidino-2-phenylindole; YFP, yellow fluorescent protein.
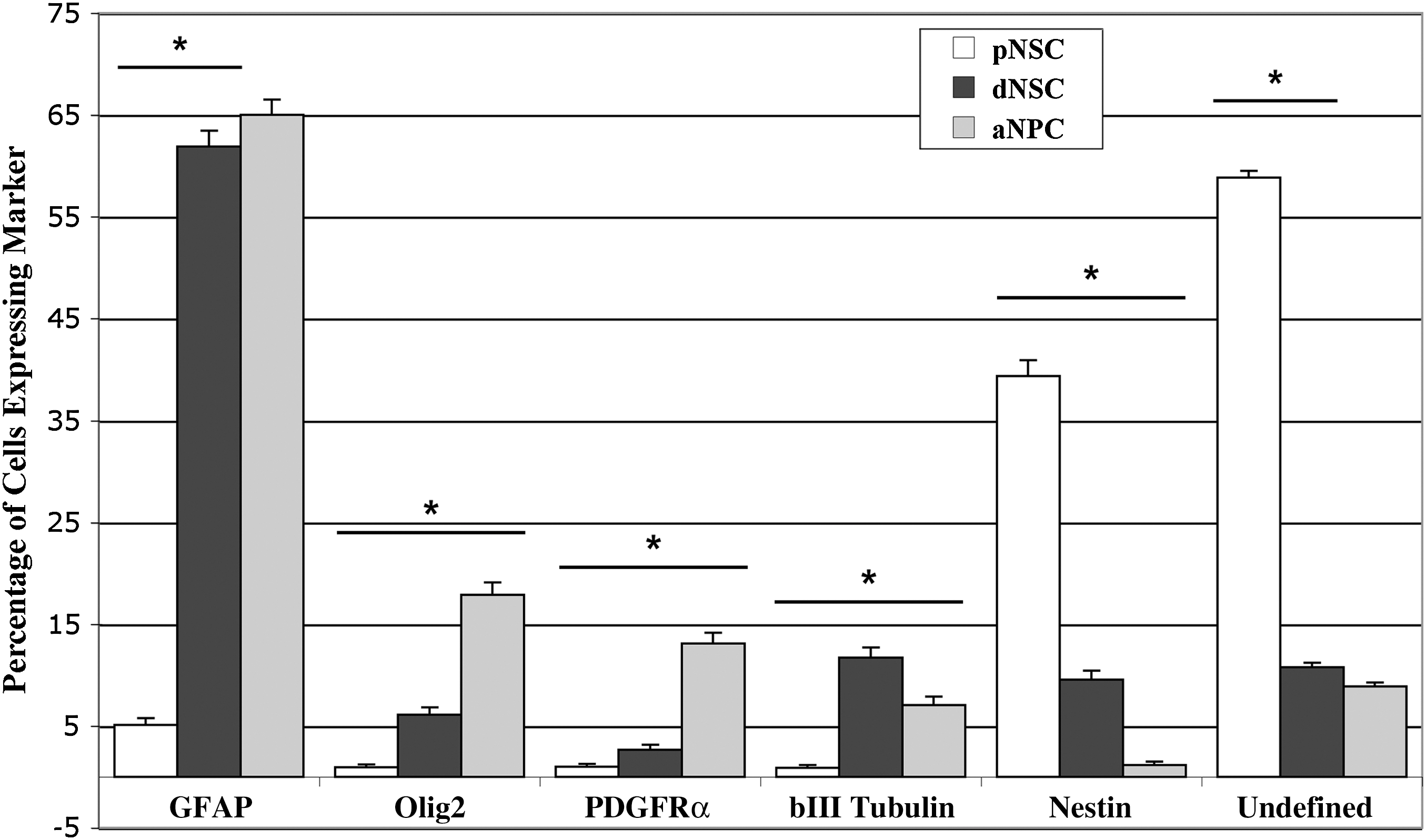
Quantification of the in vitro neural differentiation pattern of pNSCs, dNSCs, and aNPCs after 1 week in nonspecific differentiation conditions (1% FBS). Quantification of the neural differentiation pattern as proportions of cells reveals that the major cell type produced by both dNSCs and aNPCs after in vitro differentiation is GFAP-expressing astrocyte lineage cells. For pNSCs, after 1 week the largest proportion did not express any of the neural antigens used and the next largest group was nestin-expressing, undifferentiated neural stem or progenitor cells. *P<0.05.

Mature oligodendrocyte derived from ESCs using the default pathway of neural differentiation. A CNPase-expressing oligodendroycte derived from ESCs using the default pathway of neural induction. CNPase is a marker for mature oligodendrocytes in vitro. Scale bar represents 20 μm.
Base on the ES-like pattern of gene expression observed for pNSCs, it was hypothesized that the differentiation pattern of pNSCs in vitro would not produce a pure population of neural lineage cells. This was observed with the differentiation pattern of these cells appearing somewhat similar to that of ES differentiation through EB formation with many distinct morphologies that did not express any neural markers for which they were stained (data not shown). Despite this, pNSC differentiation did result in many cells expressing neural markers. Quantification revealed that the most prevalent neural marker observed was nestin, which was seen in 39%±0.8% of cells. 5%±1.5% of cells expressed GFAP, and approximately 1% of cells expressed Olig2 or PDGFRα. In areas of high cell density, beta-III tubulin-expressing cells extended long axonal-like processes that formed complex networks (Fig. 7). Accurately counting cells in these areas, using the DAPI-stained nuclei counting technique, was nearly impossible due to dense cell accumulations. Individual cells expressing beta-III tubulin that could be identified accounted for approximately 1% of pNSCs. Based on the panel of markers used, 46% of cells in 1 week differentiated pNSC cultures were defined, leaving 54% of cells undefined (Fig. 5).
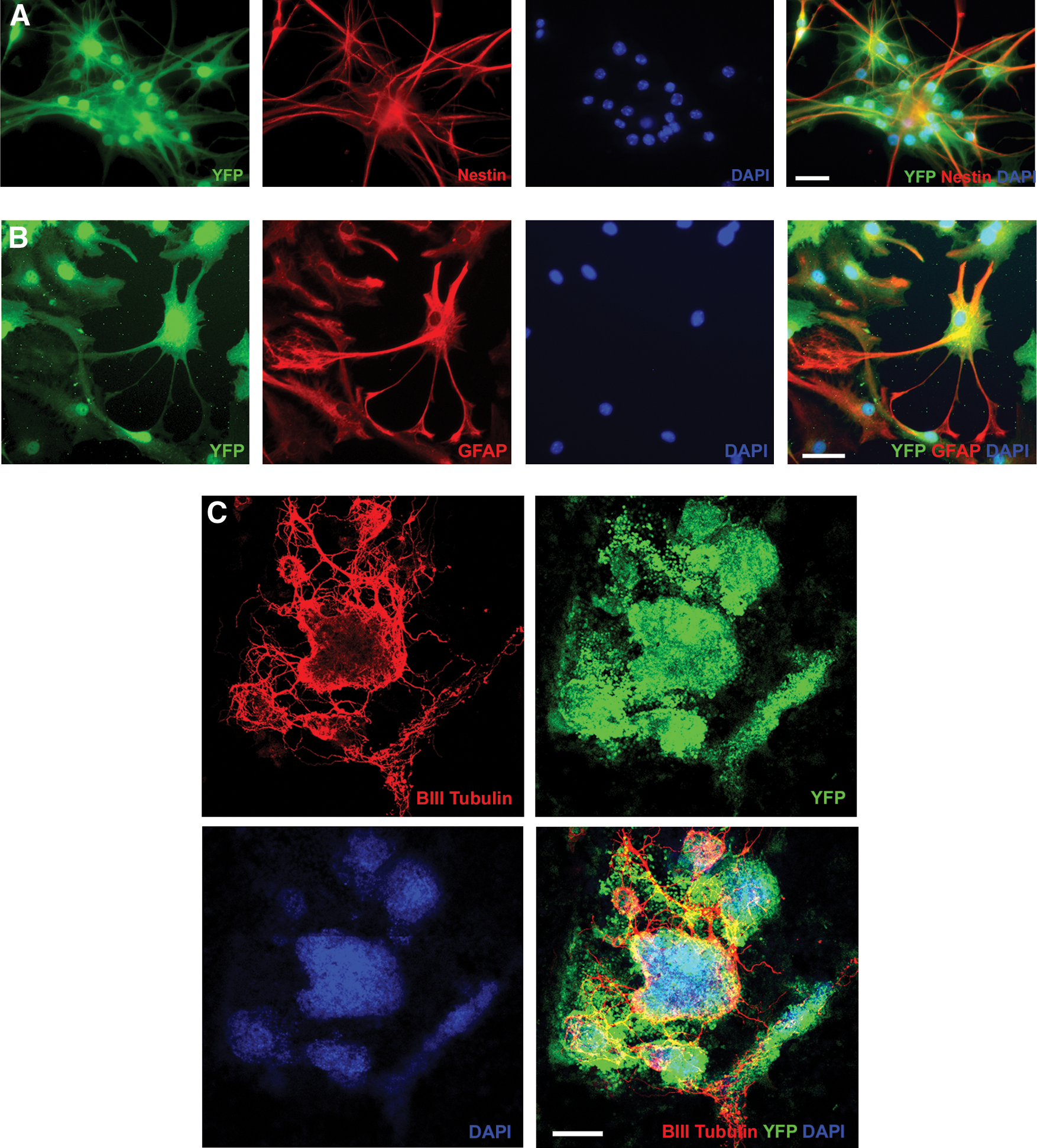
Characterizing the pattern of in vitro neural differentiation of pNSCs. The differentiation of pNSCs for 1 week in 1% FBS produced some cells that expressed neural markers.
Differentiation after transplantation into the dysmyelinated spinal cord
The findings that the properties of dNSCs in vitro were similar to aNPCs were encouraging, and suggested that the two cell types might differentiate similarly when transplanted into the dysmyelinated spinal cords of shiverer mice. In agreement with previous reports from our laboratory [27], the lack of compact myelin and MBP expression was confirmed in the shiverer mice used for this study (Supplementary Fig. S3).
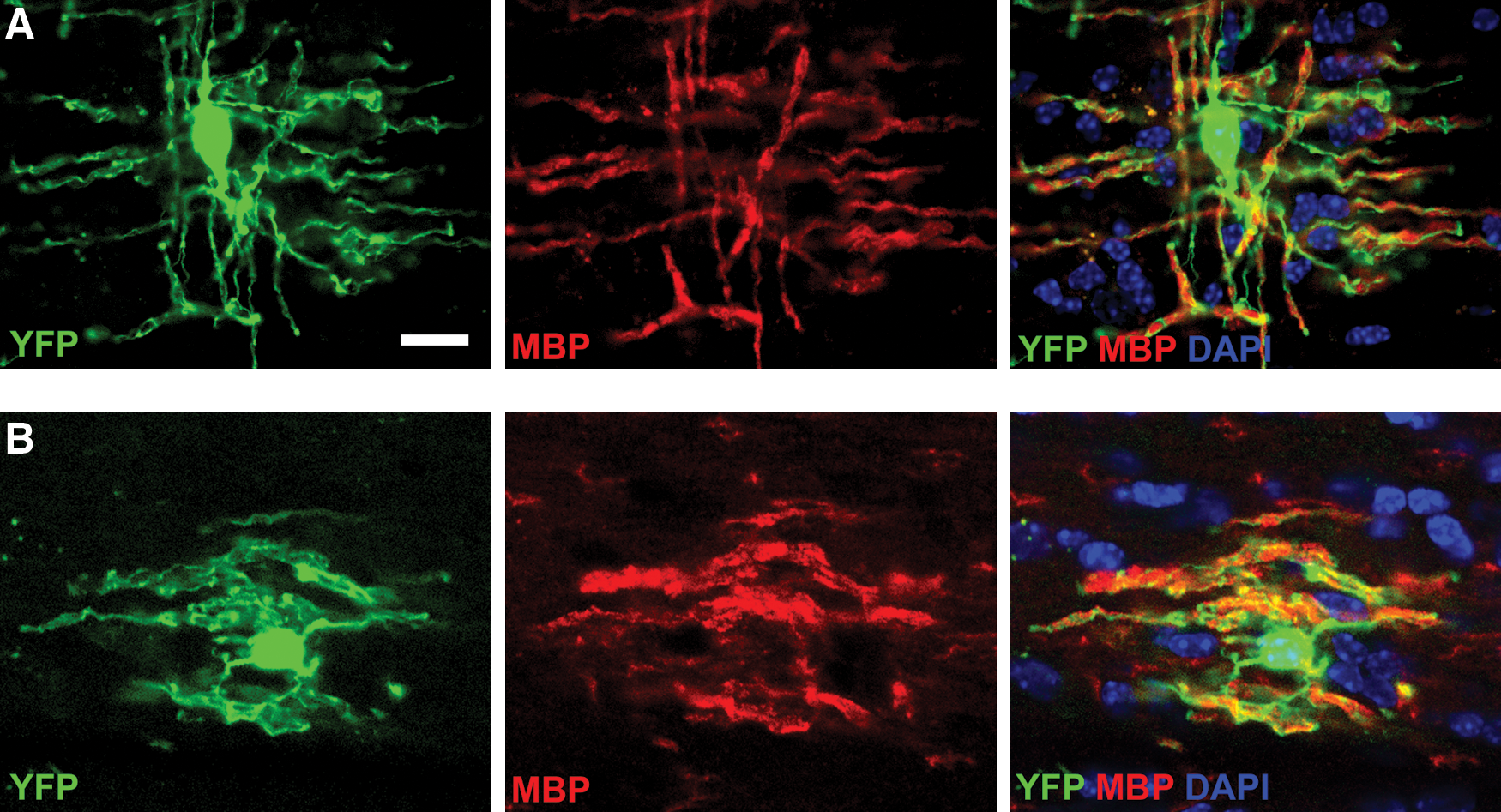
The results of our transplantation of aNPCs confirmed previous findings that aNPCs transplanted into white matter of the adult CNS can survive and integrate well into the host spinal cord parenchyma and differentiate into oligodendrocytes with multiple processes that express MBP (Figs. 8A and 9A) [1,27]. We quantified the pattern of differentiation of the aNPCs and observed that based on the neural antigens we used the majority of YFP-expressing transplanted cells differentiated into the oligodendrocyte lineage, as indicated by the expression of Olig2 by 47%±2.7%, adenomatous polyposis coli (APC) by 34%±2.7%, and PDGFRα by 15%±3.7% of transplanted cells. Note that the percentage of cells expressing APC or PDGFRα is a subpopulation of the Olig2-positive proportion. We also detected YFP-expressing cells that expressed GFAP (3.4%±3.0%), microtubule-associated protein 2 (MAP2; 3.8%±3.6%), and rare cells that maintained expression of nestin (0.7%±0.38%) (Supplementary Figs. S4 and S5). When single transplanted cells were examined at high magnification using confocal microscopy, some of the cells displayed an oligodendroctye-like morphology with multiple extended processes that aligned with host axons and stained positively for MBP (Fig. 9A).
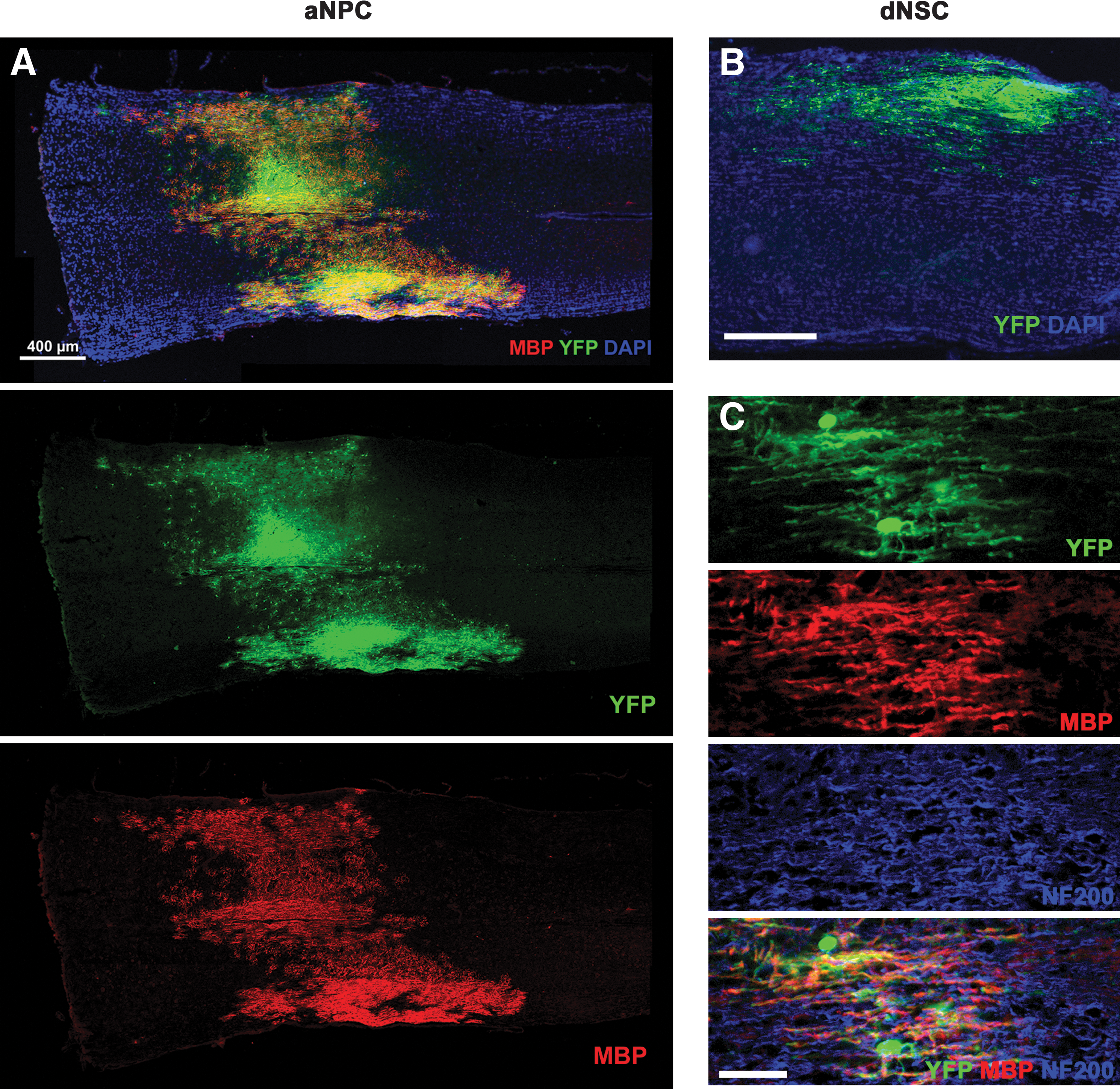
Transplantation of ES-derived definitive neurospheres and adult brain-derived neurospheres into the spinal cord of shiverer mice.
Transplantation of ESC-derived definitive neurospheres and adult brain-derived neurospheres into the spinal cord of shiverer mice produces some cells that express MBP and have the morphologies of oligodendrocytes. Transplanted aNPCs and dNSCs differentiate into cells with many properties of oligodendrocytes. Both aNPCs
In comparison to the transplantation of aNPCs, the transplantation of pNSCs did not result in successful integration of grafted cells in the spinal cord. Similar to the observations from the in vitro differentiation, a large number of YFP-expressing transplanted pNSCs failed to express any markers for which they were assessed. pNSC-transplanted regions contained areas where transplanted cell types were similar to those observed with aNPCs, although also had areas where highly irregular cell growth were observed, containing cells with an epithelial-like morphology. These cells did not express nestin, which is expressed in neuroepithelial cells (data not shown). The presence of these heterotomas was associated with locomotor impairments in the animals, and the transplantation of pNSC was not pursued further in light of these findings.
In contrast to the results obtained with pNSCs, dNSCs survived and integrated into the spinal cord white matter in a similar manner as NPCs (Figs. 8B, C and 9B). Transplanted dNSCs extended multiple processes, associated with host NF200-expressing axons, and expressed MBP consistent with the myelination of axons previously reported [27]. Individual transplanted dNSCs, viewed under high magnification confocal microscopy, had a similar morphology to that observed for aNPCs. Some extended multiple processes that aligned with host axons and expressed MBP, suggesting that they were mature, myelin-forming oligodendrocytes (Fig. 9B). Quantification of the differentiation pattern of transplanted dNSCs revealed that the majority of cells differentiated into the oligodendrocyte lineage (Figs. 10 and 11). This finding is similar to that of transplanted aNPCs, although the proportion of oligodendrocytes was significantly higher (two-tailed P<0.05) for dNSCs than for aNPCs. Cells expressing Olig2 accounted for 58%±4.5% of YFP-positive transplanted cells. Oligodendrocyte lineage-specific markers were also expressed by transplanted dNSCs, with 42±6.2% expressing APC and 19±3.0% expressing PDGFRα. GFAP was expressed by 4.2±0.4% and the neuronal marker MAP2 was detected in 3.1±0.4% of cells. Nestin-expressing cells accounted for 3.0±0.4% of all cells (Supplementary Fig. S6). Based on these data and using the expression of Olig2, GFAP, MAP2, and nestin to represent the neural lineages and undifferentiated NSCs, we found that 68% of cells of transplanted dNSCs were accounted for by our differentiation assay. The same summation of the proportional data for transplanted aNPCs showed 56%, a significantly lower percentage (2-tailed P<0.05), of cells were defined by the neural differentiation assay. Stated another way, 32% of transplanted dNSCs and 44% of transplanted aNPCs were unaccounted for based on the neural antigen panel used for quantification (Fig. 11). Previous work by our laboratory involving the quantification of the in vivo differentiation of aNPCs has observed similar portions of transplanted cell types failing to express neural markers [1].
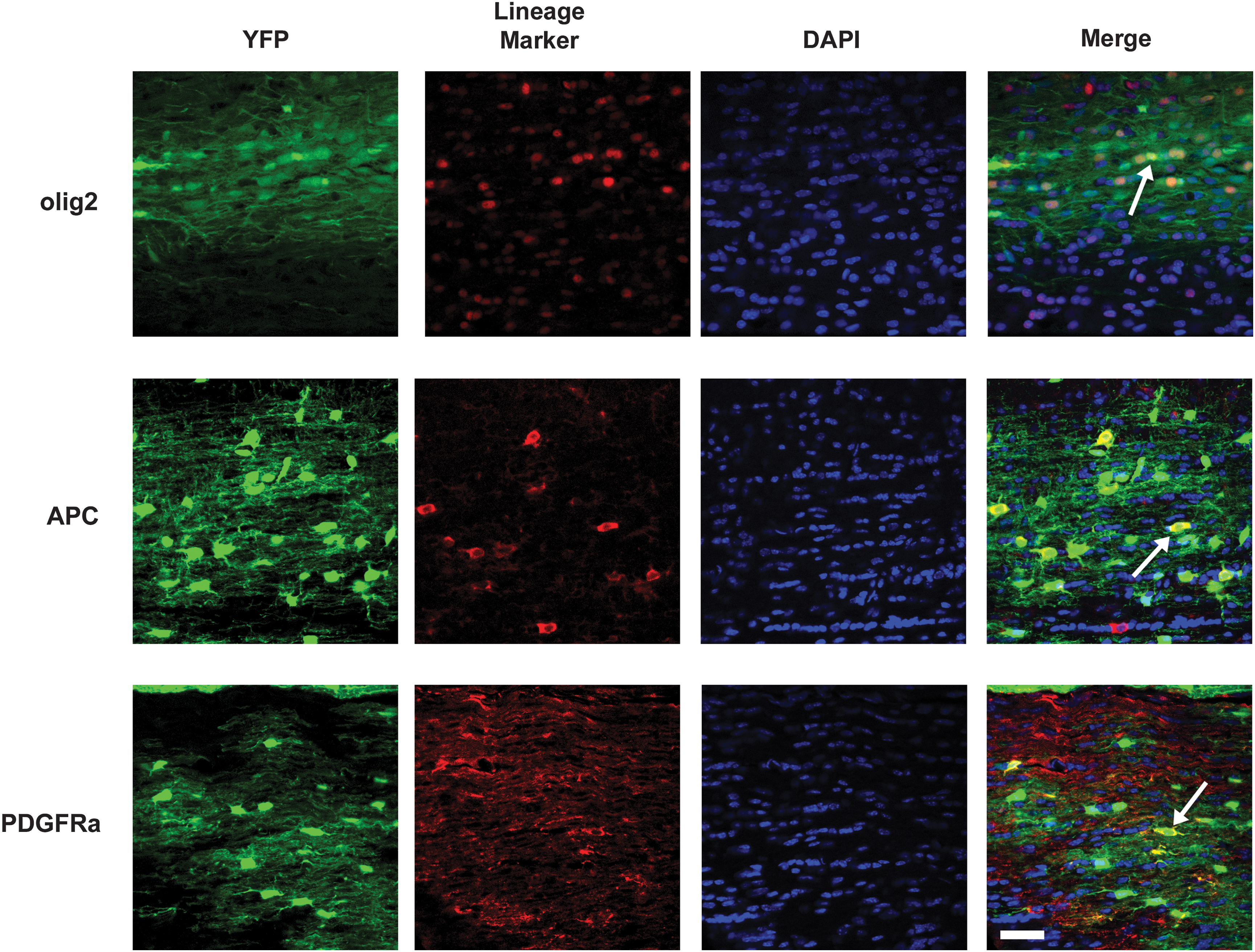
Sample counting fields for oligodendrocyte lineage markers used in quantification of the in vivo neural differentiation of pattern of dNSCs 6 weeks after transplantation into the spinal cord of shiverer mice. Typical image fields used for cell quantification are shown for transplanted dNSCs stained for the oligodendrocyte lineage markers Olig2, APC, and PDGFRα. Arrowheads indicate cells in which the lineage marker is observed to colocalize with YFP of the transplanted dNSCs. Scale bar represents 15 μm.

Neural differentiation pattern of dNSCs and aNPCs after transplantation into the dysmyelinated spinal cord of shiverer mice. Quantification of the neural differentiation pattern as proportions of surviving transplanted cells reveals that the major cell type produced by both dNSCs and aNPCs when transplanted into the shiverer spinal cord is the oligodendrocyte. Olig2-expressing oligodendrocyte lineage cells accounted for a significantly higher proportion of transplanted dNSCs then aNPCs. The Olig2-expressing cell population consisted largely of APC-expressing mature oligodendrocytes and PDGFRα-expressing oligodendroyte progenitor cells. Astrocytes and neuronal lineage cells made up a small proportion of surviving differentiating cells for both dNSCs and aNPCs. The proportion of nestin-expressing transplanted cells was small for both cell types, but a significantly higher proportion of nestin-expressing cells were observed for surviving transplanted dNSCs relative to aNPCs. *P<0.05.
Discussion
In this report we have demonstrated that the default pathway for production of NSCs from ESCs [14,15,35] is a promising method to produce neurosphere generating, clonal NSCs with properties similar to that of adult brain-derived NSCs (Table 2). Most important, we also have shown the first in vivo differentiation of NSCs (dNSCs), derived from ESCs through the default mechanism, into the oligodendrocyte lineage. We found that the dNSCs produced through the default pathway are similar to aNPCs in their neurosphere morphology, gene expression profile, in vitro pattern of neural differentiation, and in vivo pattern of neural differentiation after transplantation into the dysmyelinated spinal cord. In the shiverer spinal cord, both transplanted dNSCs and aNPCs differentiated primarily into the oligodendrocyte lineage, integrated well into the spinal cord parenchyma, and expressed MBP, suggesting the formation of myelin. Further investigation of the potential of transplanted dNSCs to produce myelinating oligodendrocytes in relevant animal models is required to determine their therapeutic potential. However, our findings are in parallel with previous studies from our laboratory [27] that showed that MBP-expressing cells derived from transplanted aNPCs can ensheath multiple axons and form compact myelin—as shown using electron microscopy—that led to improved axonal conduction as confirmed using in vivo electrophysiology. The therapeutic potential of these myelinating cells was demonstrated in a rodent model of spinal cord injury where they have been found to significantly enhance functional recovery [1,36].
FGF, fibroblast growth factor; LIF, leukemia inhibitory factor; CNS, central nervous system; EGF, epidermal growth factor; FBS, fetal bovine serum.
The default pathway and development
The production of NSCs from ESCs using the default pathway of neural induction is a powerful tool to generate large quantities of neural lineage cells. Although the precise details of the lineage relationship between the pNSC, dNSC, and aNPC remain unclear, it is likely that neural induction through the default mechanism largely recreates the neural lineage progression as it functions in normal development [37]. The default model was originally proposed based on findings in Drosophila [38] and Xenopus [39,40], but subsequent findings using murine [14,15,19] and human ESCs [41,42] have supported this model. There are striking similarities between the sequence of LIF dependence in pNSCs followed by FGF2 dependence in dNSCs observed after default neural induction in ESCs to the populations of LIF responsive neural lineage cells between embryonic days 5.5 and 7.5 (E5.5–7.5), which give rise to an FGF2-dependent cell population after E8.5 in the developing mouse embryo [37]. When looking forward to the production of cell populations for clinical application, the use of cell differentiation techniques that recreate or activate naturally occurring developmental programs is likely a valuable strategy in producing the safe and predictable cell populations.
The removal of LIF and passage of pNSCs to the dNSC results in significant cell death—LIF has been shown to function as a pro-survival factor in this culture system [14]—and this has been suggested as a limitation of ES neural differentiation protocols based on the default model [43]. While cell death is prominent after LIF withdraw once a population of dNSCs is established, the cells can be propagated in a similar fashion to aNPCs allowing for the production of large numbers of cells. It also has been demonstrated recently that when cultured at more physiological relevant oxygen concentrations than standard culture conditions, the cell death observed for both pNSC and dNSC decreases and expression of neural lineage genes such as Sox1, Sox2, nestin, and βIII tubulin is actually enhanced [35]. Therefore, the initial cell death and low yield of pNSCs and dNSCs may not be a significant limitation to the production of dNSCs with this method.
Definitive NSCs as an alternative to adult neural precursors
Based on our findings, the dNSC and aNPC are similar with respect to their gene expression profile and neural differentiation pattern both in vitro and in vivo in a model of CNS dysmyelination. After a week of nonspecific differentiating conditions in vitro, the astrocyte lineage—as defined by GFAP expression—accounted for the largest proportion of differentiated cells produced by both cell types. However, the dNSCs were observed to be more neurogenic than aNPCs under identical differentiation conditions in vitro. Neurons were the second most frequent cell type observed for differentiated dNSCs, whereas the oligodendrocyte lineage (as defined by Olig2 or PDGFRα expression) was more prevalent than the neuronal lineage (as defined by βIII-tubulin expression) in the aNPCs. This discrepancy might be explained by the normal developmental progression of the mammalian CNS in which neurogenesis dominates the early period of development, whereas gliogenesis increases with developmental age. In the developing CNS, neurogenesis is initiated at E9 and the first committed OPCs are not present until approximately E12 [44 –47]. Since the dNSC population can be considered as a developmentally more primitive cell type than the aNPC, based on its lack of EGF-dependence and being a small (<10) number of passages removed from ESCs, it is possible that the observed difference in neuronal versus oligodendrocyte differentiation is a reflection of the normal developmental time course. The comparison of long-term passaged dNSCs where EGF is included in the cultures to replicate the conditions used to culture aNPCs would help address this question, as it is unknown whether dNSCs in vitro can develop the EGF dependence, which is observed in NSCs in developing mouse embryo [37]. Notch signaling has been demonstrated to likely play an important role in this process although it is unclear if exogenous notch activation is required or if the development of EGF dependence does occur spontaneously in ES-derived dNSCs in vitro although at a very low rate [37]. Despite the differences observed during the 1-week in vitro differentiation in the more clinically relevant, 6-week differentiation period within the shiverer spinal cord there was no observed preference for dNSCs to adopt a neural fate above that of aNPCs (Fig. 11). However, despite the similar proportion of cells adopting a neuronal fate (3.1% and 3.8% for dNSCs and aNPCs, respectively) there were differences observed in the in vivo neural differentiation pattern between the 2 cell types. While Olig2-expresssing cells were the predominant cell type produced by both dNSCs and aNPCs, dNSCs did produce a significantly higher proportion of Olig2-positive cells (58% and 47%, respectively) and a significantly lower proportion of undefined cells (32% and 44%, respectively). When we examine the largest component of the Olig2-positive cell population—the oligodendrocyte lineage—we see larger proportions of both PDGFRα-expressing oligodendrocyte progenitors (19% and 15%, respectively) and mature oligodendrocytes (42% and 34%, respectively) for the progeny of the dNSCs relative to the aNPCs. We also observed enhanced proportional nestin expression (3% and 0.7%, respectively) for the progeny of dNSCs versus aNPCs. These differences could be a result of staining and counting errors or small sample size, although they also may be the result of enhanced oligodendrocyte differentiation being an inherent property of dNSCs relative to aNPCs. This is an important question that must be addressed in future work to determine the optimal cell type for remyelinating cell therapy.
The dysmyelinated spinal cord of the shiverer mouse can be considered an instructive or permissive cellular niche that likely promotes transplanted NSCs into preferentially adopting an oligodendrocyte fate. The shiverer has been shown to posses an excess of NG2-expressing oligodendrocyte progenitors [48] and it is likely that the signals resulting in this proliferative response in the NG2 cell population could also affect the fate of transplanted cells. The specific signals (and their source) that underlie this proliferative response by oligodendrocyte progenitors in the shiverer remain to be fully elucidated. Previous work has demonstrated that oligodendrocyte progenitors do respond to demyelination by mounting a proliferative response [49]. This response may be induced by the signaling molecules GRO-1 [50] and neuregulin (glial growth factor) [51], which may be released by astrocytes and neurons, respectively. Both molecules have been demonstrated to act as mitogens, to promote survival, and to inhibit differentiation of oligodendrocyte progenitors. It is likely that these signals (or others) resulting in the observed proliferative response in the NG2 cell population would also affect the fate of transplanted NSCs. Previous studies have also suggested the instructive nature of the niche as transplanted cells from ESC [52], fetal [53], and adult sources [27] have been shown to produce myelin-forming oligodendrocytes after transplantation and in other instances have demonstrated large-scale cell replacement and even rescue of the shiverer phenotype [7,8]. Here, we employed the shiverer mouse model not to directly assess the therapeutic potential of the dNSCs but as a means to compare the differentiation pattern of dNSCs to aNPCs. Our evidence for the similar behavior of the dNSC and aNPC suggests that the dNSC may be of equal therapeutic benefit as the aNPCs, which have been shown to function to replace lost cells throughout adult life in the brain [21,23,24] and have been demonstrated to have a marked ability to integrate well into CNS parenchyma when transplanted into the adult CNS—a finding that has been observed again in the experiments reported here. The observation of terminally differentiated oligodendrocytes both in vitro and after transplant into the shiverer mouse shows that differentiation based on the default pathway of neural induction is a promising method for producing clonal NSCs and their derivatives from a pluripotent cell population. Most important, this method could be of high clinical relevance by providing therapeutic NSC populations from a pluiropotent source population and by minimizing the risk of contamination of unwanted cell lineages by obviating the need for EB formation.
Addressing the undefined proportion after differentiation
Quantification of the pattern of neural differentiation both after in vitro differentiation and after transplantation in the shiverer mouse resulted in portions of the differentiated cell population being unaccounted for by the panel of neural antigens we used. The undefined proportion of the cells was difference of 1 minus the sum of the proportions for the markers Olig2, GFAP, beta-III tubulin or MAP2, and nestin, which represent the 3 neural lineages and undifferentiated NSCs. Technical staining and counting errors likely contribute to the undefined proportion of the cell populations, particulary for the quantification of the in vivo pattern of differentiation as it was not uncommon for adjacent transplanted cells with very similar morphologies to not express the same marker. In such cases, one cell was correctly counted as negative for the given marker. MBP was a very reliable indicator of mature oligodendrocytes but could not be used to for quantification since the pattern of oligodendrocyte myelination results in adjacent myelin segments on an axon coming from multiple nearby oligodendrocytes, which makes quantification very difficult and inaccurate. Previous work has demonstrated that aNPCs do not differentiate into Schwann cells (SCs) based on expression of the p75 neurotrophin receptor and the peripheral myelin marker P0 [1,27]. Future studies should address the possibility of SC differentiation, although based on the morphology of transplanted aNPCs and dNSCs observed, we hypothesize that dNSCs do not produce SCs under the differentiating conditions examined in this report. Accounting for an undefined proportion of labeled cells is further complicated by cell markers not being unique to a specific cell lineage. For example, Olig2 is typically considered an oligodendrocyte lineage marker [33] but is also expressed by NSCs and in neurospheres (data not shown), has been shown to have a role in white matter astrocyte development [54], and is expressed in the pMN progenitor cell population that yields motor neurons and oligodendrocytes [55]. While further work is required to fully characterize the progeny of both aNPCs and dNSCs, our findings here suggest that dNSCs might be of equal, if not greater, therapeutic potential as aNPCs.
Conclusions
This work has demonstrated that neural induction in ESCs through the default mechanism can produce a cell type—the definitive NSC—that is a potential substitute for NSC derived from the forebrain subependymal cells of the adult brain. Application of this approach using human ESC or induced pluripotent stem cells (iPSCs) is a very promising strategy to produce NSC for research or potential therapeutic applications such as remyelination after CNS injury. Further investigation of the cells produced through the default pathway when applied to human ES or iPS cells is required to fully determine the potential and clinical relevance of this neural induction method.
Footnotes
Acknowledgments
The authors would like to thank Desiree Schut and Jian Wang of the Fehlings laboratory and Sue Runciman and Brenda Coles of the van der Kooy laboratory for their assistance with cell culture and Behzad Ahzad for his assistance with animal care. The authors would also like to thank Dr. Soheila Karimi-Abdolrezaee for her technical expertise and guidance while conducting these experiments and Lauren Fehlings and Madeleine O'Higgins for their assistance with the article. This work was made possible by funding from the MS Society of Canada (studentship to J.W.R.), the Canadian Institutes of Health Research (operating grants to M.G.F.), and the Krembil Foundation (M.G.F. and D.V.K.).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.